
壳聚糖-碱性成纤维细胞生长因子载体诱导骨髓间充质干细胞向神经细胞分化
2011-02-27 08:31段红梅杨朝阳李晓光
中国康复理论与实践 2011年4期
段红梅,杨朝阳,李晓光
以往的观点认为,干细胞只能向其来源方向的细胞衍生。这一观点严重限制了干细胞的应用。近年的研究成果证实,骨髓间充质干细胞(bone marrow-derived mesenchymal stem cells,MSCs)在体外诱导可以向3个胚层的细胞分化[1-5]。为此,研究者采用二甲基亚砜(DMSO)、叔丁对甲氧酚(BHA)、β-巯基乙醇等各种诱导剂,单独或联合应用来促使 MSCs的分化[6-7]。近几年,在诱导过程中添加各种细胞生长因子引起人们的关注。碱性成纤维细胞生长因子(basic Fibroblast Growth Factor,bFGF)是其中的一种。研究证明,bFGF可以调控神经干细胞的更新和分化,在应用bFGF 1 d后就可以起始分化[8]。本实验主要探索MSCs经壳聚糖-bFGF载体的诱导后定向分化为神经细胞的潜能及诱导产生的细胞,并通过神经细胞的特征标记物和神经细胞的形态来验证。
1 材料和方法
1.1 实验材料 出生7 d的Wistar乳鼠由首都医科大学实验动物中心提供。试剂:α-M EM培养基(Applichem)、Nestin 小鼠单克隆抗体(Chemicon,CA)、βtubulinⅢ兔单克隆抗体(Chemicon,CA)和MAP-2鼠单克隆抗体(Chemicon,CA)。
1.2 MSCs的分离培养 无菌冰冻麻醉条件下取出生7 d的Wistar大鼠股骨和胫骨,去掉骨表面的肌肉筋膜,用PBS(0.01 mol/L)将骨冲洗干净。将骨放入含15%胎牛血清(FBS)的α-M EM培养基中,用小剪刀将骨髓钳出,再将股骨端骨松质剪掉一部分,在膝关节端用5号针头插孔,2 ml无菌注射器彻底冲洗骨髓腔;再用5号针头注射器反复冲打骨髓细胞,使之成为单细胞悬液,将骨髓细胞悬液以1∶1比例,轻轻顺离心管壁加在淋巴细胞分离液之上,1000 r/min,离心10 min分离单个核细胞。去掉顶层脂肪,吸取细胞膜层上部分,用含15%FBS的α-MEM培养基洗涤2次,1000 r/min,离心 10 min。沉淀细胞用含 15%FBS的α-MEM培养基充分分散细胞,计数。接种(1~3)×107/10 ml培养液含15%FBS的α-M EM培养基,90 mm塑料培养皿中,培养于37℃5%CO2饱和湿度培养箱内孵育。24 h后全量换液,以后每3天全量换液1次,以去掉造血细胞干扰。连续培养7 d后,当基质细胞长满培养皿80%时,用0.25%胰酶消化基质细胞,按1∶2传代培养,直至进入指数生长期,可以规则传代。
1.3 MSCs的鉴定 MSCs在连续培养到3~6代后对其进行鉴定。用0.25%胰酶将贴壁的MSCs从塑料皿底部消化下来,4℃1000 r/min离心5 min,弃上清,然后用PBS洗2遍,4℃1000 r/min离心5 min弃上清,离心管底部留200μl的液体分别加入:PE-Cy5标记的小鼠抗大鼠的CD45单克隆抗体(BD Pharmingen)、FITC标记的小鼠抗大鼠的CD90单克隆抗体(BD Pharmingen)一抗;FITC标记的小鼠抗大鼠的CD44单克隆抗体(BD Pharmingen),PE-Cy5标记的小鼠抗大鼠的CD45单克隆抗体(BD Pharmingen)一抗;30 min后流式细胞仪(COUL TER EPICS@XL,Amercia)检测。
1.4 MSCs的诱导分化 将第3代MSCs的细胞培养于铺有盖玻片的6孔板内,分别加入不同的诱导剂,实验组1:单纯壳聚糖(4 mg/ml);实验组2:单纯 b FGF(25 ng/ml);实验组3:壳聚糖-b FGF载体(4 mg/ml);对照组:只加培养基(5 mlα-MEM培养基),培养于37℃5%CO2饱和湿度培养箱内孵育,每3天半量换液1次,连续培养。
1.5 免疫组化染色 以上各组的细胞在诱导后9 d、14 d、21 d,吸出培养基 ,PBS(0.01 mol/L)洗 3 次 ,每次5 min;4%多聚甲醛,4℃固定40 min,PBS洗3次,每次 5 min;0.3%PBST浸泡 5 min,1%羊血清(0.3%PBST配制)封闭30 min;入一抗:Nestin(1∶100),β-tubulinⅢ(1∶100),37℃,孵育90 min,PBS漂洗同上,入荧光二抗(1∶100)避光,37℃,孵育60 min,PBS洗 3次,每次 5 min,Hochest:染核(1∶100),30 min,去离子水漂洗3次,每次5 min,封片。
MSCs诱导14 d后,吸出培养基,0.01 mol/L PBS洗3次,每次5 min;4%多聚甲醛,4℃固定40 min;PBS洗3次,每次5 min;3%双氧水,室温避光10 min;0.3%PBST洗 3次,每次 5 min;1%羊血清(0.3%PBST配制)封闭20 min;入MAP-2一抗(1∶800),37℃,孵育60 min;PBST洗 3次,每次 5 min;二抗(1∶300),37 ℃,孵育40 min;PBST洗3次,每次5 min;三抗(1∶300),37 ℃,孵育40 min;PBST洗3次,每次5 min;DAB呈色,相差显微镜(IX71 Olympus,Tokyo,Japan)观察。
1.6 Western blot 分别收集壳聚糖-b FGF载体诱导后 7 d、9 d、14 d、21 d 的细胞 ,4℃,12000 r/min 离心15 min,加入细胞裂解液,冰上裂解 30 min;4℃,12000 r/min离心15 min,取上清,BCA法蛋白定量,并制成蛋白含量为4.0 g/L的电泳样品,取5μl(20μg总蛋白)用8%SDS-PA GE进行电泳,聚集胶:80 V,50 min;分离胶:100 V,90 min。电泳结束后,进行蛋白质转膜:孔径0.22μm的硝酸纤维素膜(NC),4℃,电流200 mA,150 min。Western blot杂交步骤:10%脱脂牛奶封闭NC膜1 h,TTBS(20 mmol/L Tris-HCl,p H7.5;0.15 mmol/L NaCl;0.05%Tween20)漂洗3次,每次10 min,与MAP-2一抗(1∶1000)杂交反应3 h,TTBS漂洗同上,再将NC膜与辣根过氧化物酶标记的兔抗小鼠二抗(1∶4000)杂交反应1 h,之后用 TTBS漂洗 NC膜3次,每次10 min,加入 ECL试剂吹打3 min,保鲜膜包裹,暗室进行 X-光胶片曝光、显影和定影。
1.7 细胞活性的检测 用MTT(3-(4,5-dimethylthiazol-yl)2,5-diphenyltetrazolium bromide,SIGMA)比色法检测细胞活性。96孔板中每个孔内加入200μl细胞悬液,接种细胞数为104个;实验分组:单纯壳聚糖组(4 mg/ml)、单纯 bFGF组(25 ng/ml)、壳聚糖-bFGF载体组(4 mg/ml)和单纯培养基组。每组6个重复样本,细胞每3天半量换液1次。细胞在诱导1 d、3 d、7 d、14 d、21 d、28 d、35 d、42 d、49 d、56 d 后 ,每孔加入20μl的0.1 M M TT(无血清培养基配制,p H 7.2),孵育 4 h后,吸干上清,每孔内加入 160μl的DMSO振荡10 min,酶标仪570 nm检测吸光值。
1.8 统计分析 X-光胶片的结果应用 Gel-Doc凝胶成像系统扫描后,用Quantity One分析软件进行定量分析;所有的数据资料以(¯x±s)表示,统计处理采用SPSS 16.0软件包进行方差分析(ANOVA)。
2 结果
2.1 MSCs鉴定 典型的MSCs呈扁平形、梭形、三角形细胞,当细胞接近铺满时呈现出成纤维细胞样(图1)。流式结果显示,97%以上的细胞为CD44+/CD45-、CD90+/CD45-,证明从股骨和胫骨的髓腔内提取的细胞在传3~6代后90%以上的细胞为MSCs。见图2~图3。
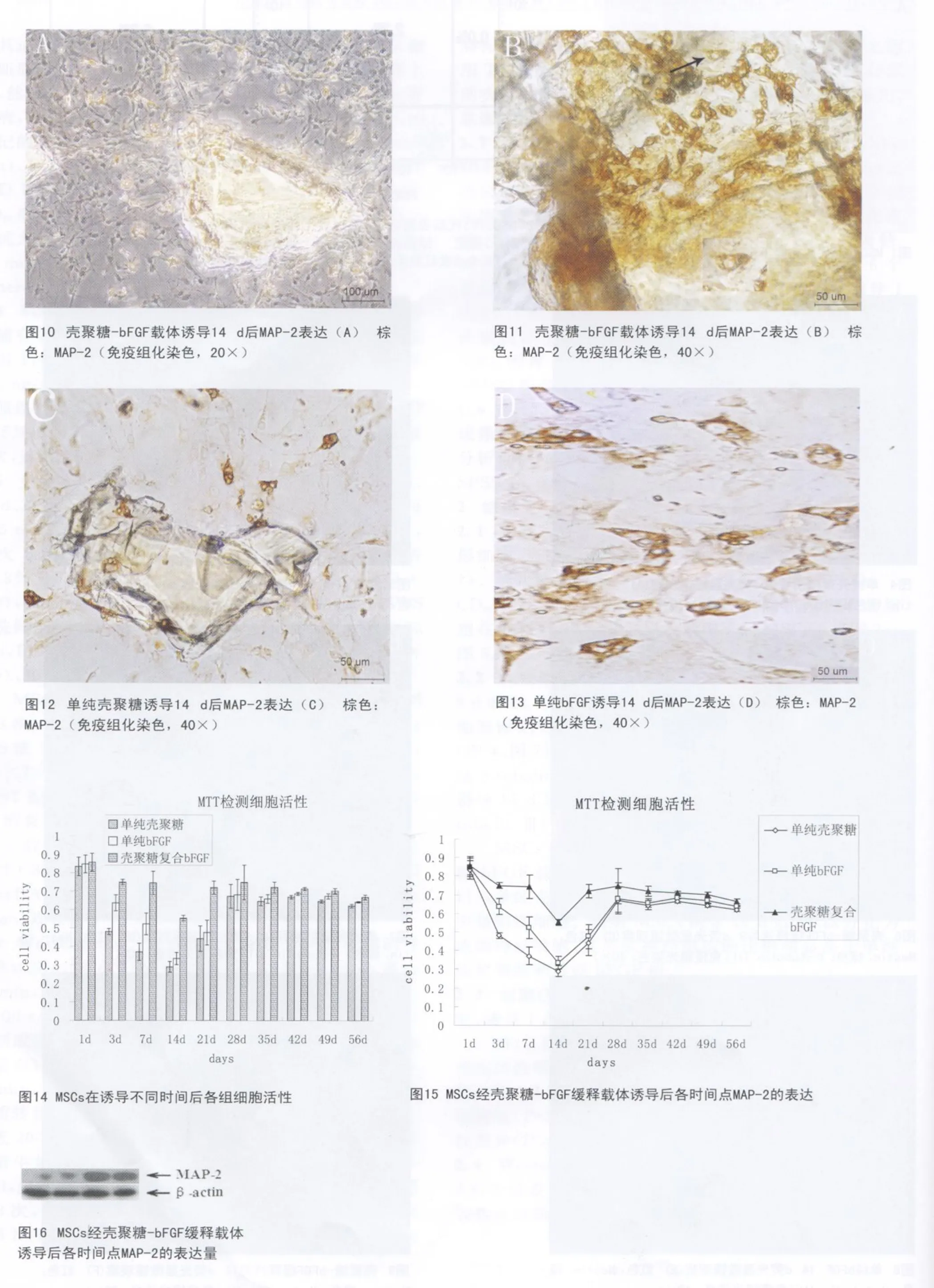
2.2 免疫组化染色 MSCs在壳聚糖-bFGF载体诱导9 d和14 d后表达神经干细胞标记物Nestin和神经细胞的标记物β-tubulin Ⅲ(图6、图9);单纯壳聚糖组(图4、图7)和单纯 bFGF组(图5、图8)仅少量细胞表达β-tubulinⅢ和Nestin;MSCs在壳聚糖-b FGF载体诱导14 d后83.54%细胞表达神经细胞的标记物βtubulin Ⅲ(图 9)。
MSCs在诱导14 d后,免疫组化染色观察到:壳聚糖-b FGF载体组有大量 MAP-2阳性细胞(图 10、图11),单纯壳聚糖组(图12),单纯 bFGF组(图13)仅有少量阳性细胞。表明MSCs诱导14 d后,可以分化为成熟神经细胞,且形成神经细胞的比例明显高于单纯壳聚糖组和单纯bFGF组。
2.3 细胞活性的检测 细胞活性通过M TT比色法测定,诱导1 d后在细胞活性方面3组无显著性差异(P>0.05)。诱导 3 d、7 d、14 d、21 d后壳聚糖-bFGF组细胞活性明显高于单纯壳聚糖组和单纯b FGF组,诱导3 d、7 d后单纯b FGF组细胞活性明显高于单纯壳聚糖组(P<0.01)。诱导28 d后3组细胞活性无显著性差异(P>0.05)。见图14。
2.4 Western blot MSCs经壳聚糖-bFGF诱导后7 d有少量表达MAP-2,随着诱导时间延长MAP-2的表达量逐渐升高,且在14 d后达到稳定(图15、图16)。
3 讨论
近几年很多研究都把目光集中在一些因子对干细胞的增殖和分化作用上,这些因子包括介质中弥散因子[8-9]和参与细胞间相互作用的因子[10-11]。它们可以增强或抑制干细胞的增殖和分化。例如:一直都认为bFGF是神经干细胞(NSC)增殖和分化的一个关键调节因子[12],并且可以促进MSCs和间充质祖细胞向神经细胞分化[13]。研究表明,在缺乏b FGF的小鼠中观察到存在神经缺陷。这一结果证实,bFGF在神经发生中具有十分重要的作用[14]。
在本实验中,壳聚糖-bFGF载体可以定向诱导MSCs向神经细胞分化,且诱导比例明显高于单纯bFGF组和单纯壳聚糖组。其中单纯壳聚糖组分化的比例最低。产生这一现象主要原因为:bFGF加入培养基中很快失去活性,此时被诱导的细胞不能得到持续的b FGF的作用;壳聚糖-b FGF载体对b FGF具有缓释作用,能够长时间缓慢释放bFGF[15],保证bFGF在培养基中持续发挥作用,使细胞长时间保持在稳定而持续的诱导状态中,提高被诱导细胞向神经细胞分化的比例,壳聚糖-b FGF载体对b FGF的缓释作用,在保证高效率诱导的同时又减少bFGF因子的浪费。
细胞黏附分子CD44、CD90是MSCs的表面标志性抗原,白细胞标志抗原CD45是造血干细胞的表面标志性抗原之一。因此,CD44+/CD45-、CD90+/CD45-细胞被认为是骨髓间充质细胞。本实验的流式结果显示,97%以上的细胞为CD44+/CD45-、CD90+/CD45-,证明从股骨和胫骨的髓腔内提取的细胞在传3~6代后90%以上的细胞为MSCs。为了验证壳聚糖-b FGF载体可以使MSCs分化为神经细胞,本实验采用免疫组化染色间接分析各时间点分化细胞的表型。β-tubulinⅢ和MAP-2是成熟神经细胞的标记,Nestin是神经干细胞的标记。形态学和分子水平上的结果均显示,MSCs经壳聚糖-bFGF载体诱导后表达神经细胞的标记物,且在诱导14 d后大量表达成熟神经细胞的标记物MAP-2。证明壳聚糖-bFGF载体可以诱导MSCs形成成熟神经细胞。
M TT是定量测定细胞活性的一种实验方法,也是当前检测生物材料的生物相容性和细胞毒性的主要方法[16]。本实验采用壳聚糖和MSCs共培养,目的是为了研究壳聚糖载体是否对干细胞具有细胞毒性和诱导作用。从M TT的结果可以看出,单纯壳聚糖对干细胞无细胞毒性,不会抑制干细胞的分化;单纯壳聚糖组和壳聚糖-b FGF载体组在诱导28 d后细胞活性并无显著性差异(P>0.05)。
本实验得出,应用壳聚糖-bFGF载体可以诱导MSCs高比例向类神经细胞样细胞分化(83.54%)。关于诱导形成神经细胞是兴奋性还是抑制性,诱导产生的神经细胞是否有功能,及其诱导分化的机制等问题,仍需要进一步验证和探索。
[1]Pittenger MF,Mackay AM,Beck SC,et al.Multilineage potential of adult human mesenchymal stem cells[J].Science,1999,284:143-147.
[2]Makino S,Fukuda K,Miyoshi S,et al.Cardiomyocytes can be generated from marrow stromal cells in vitro[J].J Clin Invest,1999,103:697-705.
[3]Deans RJ,Moseley AB.Mesenchymal stem cells:biology and potential clinical uses[J].Exp Hematol,2000,28:875-884.
[4]Kopen GC,Prockop DJ,Phinney DG.Marrow stromal cells migrate throughout forebrain and cerebellum,and they differentiate into astrocytes after injection into neonatal mouse brains[J].Proc Natl Acad Sci USA,1999,96:10711-10716.
[5]Tropel P,Platet N,Platel JC,et al.Functional neuronal differentiation of bone marrow-derived mesenchymal stem cells[J].Stem Cell,2006,24:2868-2876.
[6]Rismanchi N,Floyd CL,Berman RF,et al.Cell death and long-term maintenance of neuron-like state after differentiation of rat bone marrow stromal cells:A comparison of protocols[J].Brain Res,2003,991:46-55.
[7]Sanchez-Ramos JR.Neural cells derived from adult bone marrow and umbilical cord blood[J].J Neurosci Res,2002,69:880-893.
[8]Cattaneo E,Mc Kay R.Proliferation and differentiation of neuronal stem cells regulated by nerve growth factor[J].Nature,1990,347:762-765.
[9]Tzeng SF.Neural progenitors isolated from newborn rat spinal cords differentiate into neurons and astroglia[J].J Biomed Sci,2002,9:10-16.
[10]Johe KK,Hazel TG,Muller T,et al.Singlefactors direct the differentiation of stem cells from the fetal an adult central nervous system.[J].Genes Dev,1996,10:3129-3140.
[11]Young TH,Hung CH.Behavior of embryonic rat cerebral cortical stem cells on the PVA and EVAL substrates[J].Biomaterials,2005,26:4291-4299.
[12]Palmer TD,Markakis EA,Willhoite AR,et al.Fibroblast growth factor-2 activates a latent neurogenic program in neural stem cellsfrom diverse regions of the adult CNS[J].J Neurosci,1999,19:8487-8497.
[13]Solchaga LA,Penick K,Porter JD,et al.FGF-2 enhances the mitotic and chondrogenic potentials of human adult bone marrow-derived mesenchymal stem cells[J].J Cell Physiol,2005,203:398-409.
[14]Vaccarino FM,Schwartz ML,Raballo R,et al.Changes in cerebral cortex size are governed by fibroblast growth factor during embryogenesis[J].Nat Neurosci,1999,2:246-253.
[15]Yang ZY,Duan HM,Mo L H,et al.The effect of the dosage of NT-3/chitosan carriers on the proliferation and differentiation of neural stem cells[J].Biomaterials,2010,31:4846-4854.
[16]Thonhoff JR,Lou DI,JordanPM,et al.Compatibility of human fetal neural stem cells with hydrogel biomaterials in vitro[J].Brain Res,2008,1187:42-51.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
昆明医科大学学报(2021年10期)2021-12-02
中风与神经疾病杂志(2021年9期)2021-11-08
家庭医药(2021年7期)2021-07-23
河北科技师范学院学报(2021年1期)2021-05-10
昆明医科大学学报(2021年2期)2021-03-29
中成药(2017年12期)2018-01-19
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
