
3*Flag-hPAK4重组质粒的构建及其在COS7细胞中的表达与定位
2011-01-06 06:16张红艳王春玉李彦姝王迪李丰
东南大学学报(医学版) 2011年6期
张红艳,王春玉,李彦姝,王迪,李丰
(中国医科大学基础医学院细胞生物学教研室,细胞生物学卫生部重点实验室,辽宁沈阳 110001)
Rho家族鸟苷三磷酸酶(Rho、Rac、Cdc42)作为调节不同细胞进程的分子开关,参与从细胞黏附、迁移到细胞生存、凋亡的过程[1]。p21活化的激酶(PAKs)是Cdc42和Rac下游重要的靶蛋白。到目前为止,PAK家族共有6个成员[2],所有成员都包含一个高保守的丝氨酸苏氨酸激酶结构域,其激酶活性与PAKs执行细胞功能有关[2]。PAKs与多种细胞内蛋白相互作用,提示PAKs是多向性激酶,参与多种细胞信号通路,例如:参与细胞骨架的信号通路,参与细胞生存通路[3]。根据结构和序列同源性将PAKs分为两大类:Ⅰ类 PAK成员包括 PAK1、PAK2、PAK3,Ⅱ类 PAK成员包括 PAK4、PAK5、PAK6[4]。
PAK4是目前研究较为广泛的Ⅱ类PAK,并且表达广泛,胚胎及成年组织中均有表达,尤其在前列腺、睾丸、结肠组织中高表达[5]。PAK4与肿瘤关系非常密切,PAK4在许多肿瘤细胞系中和源于乳腺、肺、胰腺、结肠、前列腺的肿瘤中高表达[6-9]。PAK4的研究对于肿瘤的发生发展以及治疗有非常重要的作用。本研究构建3*Flag-hPAK4质粒,纯化PAK4蛋白,研究PAK4相互作用蛋白,为肿瘤信号转导通路提供依据,可以使用标签抗体Flag观察PAK4在细胞中的定位。
1 材料与方法
1. 1 菌株、细胞和质粒
大肠杆菌DH5α感受态为本实验室制备,MCF-7乳腺癌细胞系和COS7细胞为本实验室保存,3*Flag载体购自Sigma公司。
1. 2 主要试剂
PyrobestTMDNA polymerase、dNTP、DNA 电泳凝胶回收试剂盒、反转录试剂、限制性核酸内切酶BamHⅠ和EcoRⅠ均购自大连 TaKaRa公司;TRIzol RNA提取试剂购自Invitrogen公司;DNA marker、Protein marker购自GenScript公司;T4 DNA连接酶购自NEB公司;Flag抗体购自上海Genomics公司;HRP标记的羊抗鼠IgG购自北京中杉金桥生物技术有限公司;Alexa Fluor 594、DAPI购自 Invitrogen公司;引物合成、DNA测序由上海生物工程有限公司完成;ECL发光试剂盒购自购自GE Healthcare公司;其他试剂均为国产分析纯。
1. 3 MCF-7乳腺癌细胞RNA提取及反转录
MCF-7细胞用含10%胎牛血清的RPMI-1640培养基培养至90%融合,提取细胞总RNA。在3.5 cm直径的培养板中加入1 ml TRIzol,用移液器反复吹打,直到细胞全部裂解。RNA提取过程按照Invitrogen公司提供的TRIzol说明书进行操作。测RNA的OD260和OD280值,计算比值和浓度。将MCF-7提取的RNA稀释到浓度为 1 μg·μl-1,进行反转录。操作步骤按照TaKaRa反转录试剂盒说明书进行。
1. 4 hPAK4全长基因扩增
设计hPAK4扩增的PCR引物,并在引物中加入了EcoRⅠ和BamHⅠ两个限制性酶切位点。上游hPAK4引物为5'-TCCAGAATTCCATGTTCGGGAAGAG GAAG-3',下游 hPAK4引物为 5'-ATATTGGATCCT CATCTGGTGCGGTTCTG-3'。以MCF-7 cDNA为模板PCR扩增hPAK4全长编码序列。
1. 5 3*Flag-hPAK4表达载体的构建
将3*Flag载体和hPAK4 PCR片段分别用BamHⅠ和EcoRⅠ双酶切后,凝胶回收纯化产物。将两个片断用T4 DNA连接酶常温连接2 h,然后16℃连接过夜,取5 μl转化到大肠杆菌感受态DH5α中,37℃培养过夜。挑取菌落,接种于含氨苄西林的LB培养基中,37℃振荡过夜。用碱裂解法提取质粒DNA,用BamHⅠ和EcoRⅠ双酶切鉴定外源基因的插入,质粒DNA送上海生物工程有限公司进行测序分析。
1. 6 3*Flag-hPAK4瞬时转染COS7细胞系
COS7细胞用含10%胎牛血清的DMEM高糖培养基培养,细胞铺于六孔板至70% ~80%融合,按照Abmart公司提供的TranSmarter转染六孔板的说明书进行操作。
1. 7 蛋白质提取与Western blotting鉴定
转染24 h后,弃掉培养基,用PBS清洗两遍。在细胞中加入60 μl含有蛋白酶抑制剂的RIPA裂解液,用橡皮刮刀将细胞缓慢刮下。收集所有液体到新的离心管中,冰上放置10 min裂解细胞。13 000×g 4℃离心30 min,将上清转到新的离心管中,取上清5 μl,选用G250考马斯亮蓝方法进行蛋白定量。
将蛋白定量后,取 80 μg总蛋白经 10%SDS-PAGE凝胶分离,4℃过夜40 V恒压转移到PVDF膜上,5% 脱脂奶粉封闭1 h。用Flag抗体(1∶1 000稀释)室温孵育2 h,然后TBST洗膜3次,每次10 min。再用辣根过氧化物酶标记的羊抗鼠二抗(1∶5 000稀释)孵育2 h,然后TBST洗膜3次,每次10 min。ECL显影。
1. 8 共聚焦激光扫描显微镜观察3*Flag-PAK4细胞内定位
将3*Flag-PAK4转染到COS7细胞中,24 h后用4%多聚甲醛固定,0.1%TritonX100通透细胞膜,山羊血清封闭1 h,anti-Flag抗体(1∶150稀释)室温孵育1 h,PBS洗3 次,每次 10 min,Alexa Fluor 594(1∶100稀释)室温孵育1 h,PBS洗3次,每次10 min,DAPI(1∶1 000稀释)室温孵育 20 min,PBS洗 3次,每次10 min,使用50%甘油封片。在激光共聚焦显微镜下观察,红色区域为PAK4蛋白在细胞内的定位,蓝色代表细胞核。
1. 9 免疫沉淀
将COS7细胞均匀分配,铺于两个10 cm培养板中至 70% ~80%融合,按照 Abmart公司提供的TranSmarter转染试剂说明书进行操作。其中一个转染3*Flag空载,另一个转染3*Flag-hPAK4质粒。24 h后收集细胞,使用免疫沉淀裂解液裂解细胞,10 000 r·min-1离心30 min,转移上清到新的 EP 管中,加入4 μl anti-Flag抗体,4℃旋转1 h,加入50%Protein A beads 30 μl 4℃旋转3 h,用免疫沉淀裂解液洗涤3次,500×g离心2 min,弃上清。加入等体积的2*loading buffer,SDS-PAGE凝胶分离,4℃过夜40 V恒压转移到PVDF膜上,5% 脱脂奶粉封闭1 h。用anti-Flag抗体(1∶1 000稀释)室温孵育2 h,然后TBST洗膜3次,每次10 min。再用辣根过氧化物酶标记的羊抗鼠二抗(1∶5 000稀释)孵育2 h,然后TBST洗膜3次,每次10 min。ECL显影。
2 结 果
2. 1 PCR反应扩增hPAK4基因全长
以MCF-7 cDNA为模板,特异性的引物进行聚合酶链反应扩增hPAK4基因全长。扩增条件为95℃5 min预变性;95 ℃ 1 min,50 ℃ 1 min,72 ℃ 1 min,进行30个循环;72℃ 10 min,温度降至4℃。1%水平琼脂糖凝胶电泳,溴化乙啶染色后,在紫外灯下观察结果(图1)。
2. 2 重组质粒3*Flag-hPAK4的构建及鉴定
将重组质粒3*Flag-hPAK4用BamHⅠ和EcoRⅠ双酶切后得到约6 100 bp和1 800 bp的两条带(图2),重组质粒的酶切鉴定结果表明质粒构建成功,经过测序分析,外源系列在NCBI-Blast比对结果与hPAK4编码序列一致。
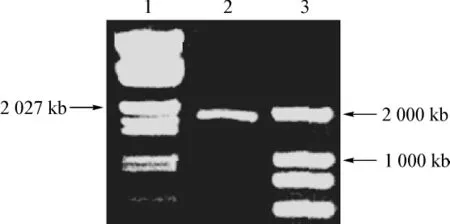
图1 PCR扩增hPAK4基因Fig 1 PCR amplification of the full length hPAK4

图2 BamHⅠ和EcoRⅠ双酶切鉴定重组质粒3*Flag-hPAK4Fig 2 Restriction enzyme digestion analysis of the recombinant plasmid 3*Flag-hPAK4
2. 3 Western blotting检测蛋白表达
分别将3*Flag空载和3*Flag-hPAK4转染COS7细胞,24 h后收集细胞,提取蛋白,经SDS-PAGE凝胶电泳,Western blotting检测蛋白表达,转染3*FlaghPAK4,见一明显条带(图3),空载体对照组未出现反应条带。3*Flag-hPAK4分子质量约为68 kDa,为质粒本身的3*Flag标签分子质量(1.65 kDa)和hPAK4分子质量68 kDa之和。表明构建的真核表达质粒在COS7细胞中表达。
2. 4 3*Flag-hPAK4在细胞内的定位
通过共聚焦激光扫描显微镜下观察3*FlaghPAK4在细胞内的定位(图4),激发光分别为594 nm(红色),340 nm(蓝色)。
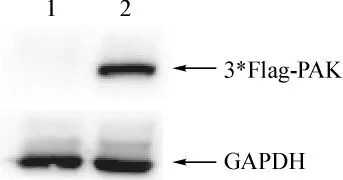
图3 Western blotting检测3*Flag-PAK4融合蛋白的表达Fig 3 Western blotting detect the expression of 3*Flag-PAK4 fusion protein using anti-Flag mAb
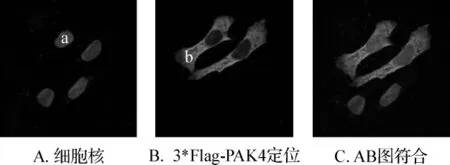
图4 激光扫描共聚焦显微镜观察3*Flag-PAK4在COS7细胞内的定位Fig 4 Confocal microscopy pictures of COS7 cells show the subcellular localization of 3*Flag-PAK4
2. 5 免疫沉淀
分别将3*Flag空载和构建的3*Flag-hPAK4转染COS7细胞,24 h后收集细胞,提取蛋白,使用anti-Flag抗体沉降蛋白,经10%SDS-PAGE凝胶电泳,Western检测到被沉降的3*Flag-hPAK4蛋白的表达(图5)。
3 讨 论
Ⅱ类PAK是多向性的激酶,参与多种细胞功能,包括细胞运动、神经元生长的调节、激素信号通路、基因转录、细胞存活等。因此这些蛋白参与癌症的进展和神经系统疾病。其中PAK4是Ⅱ类PAK中发现最早,也是目前研究的最为广泛的成员。有证据表明PAK4通过两条不同的通路促进细胞存活:PAK4增加BAD 磷酸化水平[10]和延迟半胱天冬酶分裂[10-11]。在正常的细胞中PAK4能使细胞周期停滞,在高表达PAK4的细胞中,上调P53和细胞周期调节蛋白的表达[12]。PAKs影响细胞运动和细胞存活通路,已经成为癌症的治疗靶点。研究证明Ⅱ类PAK的活性影响癌细胞的性能。PAK4高表达是致癌的[6],PAK4能抑制癌细胞凋亡,PAK4与肿瘤坏死因子α受体复合物相互作用,促进细胞存活通路[13]。体内试验表明PAK4能促进肿瘤发生,PAK4表达缺失减弱V12Ras引起的肿瘤发生[14]。肿瘤侵袭和转移是多阶段、多基因参与的过程,相关基因的调节机制异常复杂[15]。利用3*Flag-PAK4质粒研究PAK4对肿瘤转移侵袭的影响很有意义。
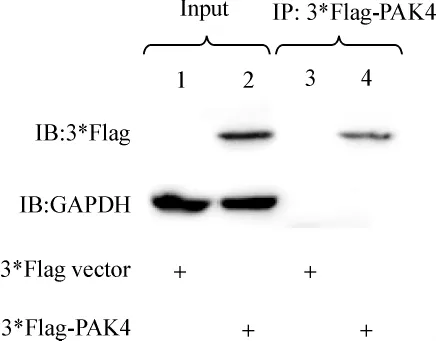
图5 3*Flag抗体免疫沉淀PAK4蛋白Fig 5 3*Flag antibody immunoprecipitate PAK4 protein
本研究为了能在真核细胞中表达PAK4蛋白,用于在体或离体的实验研究,深入探讨PAK4的生物学功能,利用DNA重组技术将PAK4重组到3*Flag载体中,通过酶切和测序保证扩增片段的正确性;通过Western blotting方法证实融合蛋白的正确性。融合蛋白中含有3*Flag亲和标签可用于辅助目标蛋白的纯化和检测。本研究构建了PAK4的真核表达载体,并且重组质粒中带有3*Flag标签,为Western blotting检测基因表达提供了保证。本实验通过亚克隆的方法成功构建了3*hFlag PAK4融合蛋白重组质粒,并验证了表达和定位,为从细胞和分子水平研究PAK家族的生物学功能奠定了良好的基础。
[1]RIDLEY A J,SCHWARTZ M A,BURRIDEG K,et al.Cell migration:integrating signals from front to back[J].Science,2003,302(5651):1704-1709.
[2]ARIAS-ROMERO L E,CHERNOFF J.A tale of two Paks[J].Biol Cell,2008,100(2):97-108.
[3]WELLS C M,JONES G E.The emerging importance of group II PAKs[J].Biochem J,2010,425(3):465-473.
[4]PODHORSKA-OKOLOW M,DZIEGIEL P,GOMULKIEWICZ A,et al.Exercise-induced apoptosis in rat kidney is mediated by both angiotensin II AT1 and AT2 receptors[J].Histol Histopathol,2006,21(5):459-466.
[5]ABO A,QU J,CAMMARANO M S,et al.PAK4,a novel effector for Cdc42Hs,is implicated in the reorganization of the actin cytoskeleton and in the formation of filopodia[J].EMBO,1998,17(22):6527-6540.
[6]CALLOW M G,CLAIVOYANT F,ZHU S,et al.Requirement for PAK4 in the anchorage-independent growth of human cancer cell lines[J].J Biol Chem,2002,277(1):550-558.
[7]CHEN S,AULETTA T,DOVIRAK O,et al.Copy number alterations in pancreatic cancer identify recurrent PAK4 amplification[J].Cancer Biol Ther,2008,7(11):1793-1802.
[8]KIMMELMAN A C,HEZEL A F,AGUIRRE A J,et al.Genomic alterations link Rho family of GTPases to the highly invasive phenotype of pancreas cancer[J].Proc Natl Acad Sci U S A,2008,105(49):19372-19377.
[9]PARSONS D W,WANG T L,SAMUELS Y,et al.Colorectal cancer:mutations in a signalling pathway[J].Nature,2005,436(7052):792.
[10]GNESUTTA N,QU J,MINDEN A.The serine/threonine kinase PAK4 prevents caspase activation and protects cells from apoptosis[J].J Biol Chem,2001,276(17):14414-14419.
[11]LU Y,PAN Z Z,DEVAUX Y,et al.p21-activated protein kinase 4(PAK4)interacts with the keratinocyte growth factor receptor and participates in keratinocyte growth factor-mediated inhibition of oxidant-induced cell death[J].J Biol Chem,2003,278(12):10374-10380.
[12] CAMMARANO M S,NEKRASOVA T,NOEL B,et al.Pak4 induces premature senescence via a pathway requiring p16INK4/p19ARF and mitogen-activated protein kinase signaling[J].Mol Cell Biol,2005,25(21):9532-9542.
[13]LI X,MIDEN A.PAK4 functions in tumor necrosis factor(TNF)α-induced survival pathways by facilitating TRADD binding to the TNF receptor[J].J Biol Chem,2005,280(50):41192-41200.
[14]LIU Y,XIAO H,TIAN Y,et al.The pak4 protein kinase plays a key role in cell survival and tumorigenesis in athymic mice[J].Mol Cancer Res,2008,6(7):1215-1224.
[15]刘飞,黄培林,黄照权,等.外源性FHIT基因转染对人胃癌细胞系MKN45迁移和侵袭能力的影响[J].东南大学学报:医学版,2009,28(5):397-401.
猜你喜欢
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
食品科学(2018年10期)2018-05-23
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中国医药生物技术(2015年4期)2015-12-26
华南农业大学学报(2015年5期)2015-12-04
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
