
艾比湖湿地自然保护区8个乔灌木种群空间分布格局分析
2010-12-31 13:31:32杨晓东吕光辉张雪梅孙丽君何书林
植物资源与环境学报 2010年4期
杨晓东,吕光辉,①,张雪梅,孙丽君,何书林
种群空间分布格局是指种群个体在水平空间的分布状况或配置状况,是种群自身的生物学特性、种内和种间相互关系以及所处生境异质性等综合作用的空间表现形式,不仅能反映某个种群适应生境后对空间的占有格局,还能反映出与其他种群相互作用和影响的过程[1]。种群空间分布格局随着空间和时间限度的改变而变化,反映了种群个体在水平空间上彼此间的相互关系是由种群本身生物学特性、种间关系和环境条件的综合影响所决定的[2]。种群空间分布格局一般分为随机分布、均匀分布和聚集分布3个类型[3]。目前,关于种群空间分布格局的研究方法较多[4-6],许多学者也运用各种方法对不同植物种群的空间分布格局进行了研究[1,7-10],但大多是对单个物种的空间分布格局进行分析,有关物种在相互作用下的空间分布格局的研究尚不多见[11-12]。
艾比湖湿地是中国不多见的荒漠风口湖泊湿地,分布着荒漠、戈壁和湿地等自然景观,区域内的植物起遏制新疆北疆沙尘源头的作用;同时,艾比湖湿地特殊的地理位置和脆弱的生态系统对入湖水量和植被退化等变化非常敏感。作者运用空间分布格局的基本分析方法,结合总体方差比率(VR),研究了艾比湖湿地自然保护区8个乔灌木种群在相互联结情况下的空间分布格局,以期为阐明当地植物群落的形成、维持植物群落的稳定性与演替规律以及为艾比湖湿地自然保护区植被的恢复与重建提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
艾比湖湿地自然保护区位于新疆维吾尔自治区境内,地理坐标为北纬44°30~45°09′、东经82°36′~83°50′,是准噶尔盆地西部最低洼地和水盐汇集中心[13]。它由以湖泊为核心的湿地和湖周林草地两大生态系统构成,总面积2670.85 km2。区内气候极端干燥,降水稀少,属典型温带大陆性干旱气候。年降水量约100mm,蒸发量在1600mm以上;年日照时数约2800 h;极端最高气温44℃,极端最低气温-33℃,年平均气温6℃~8℃。西北部阿拉山口是中国著名的风口,盛行西北风[14]。区域内有石漠、砾漠、沙漠、盐漠、沼泽和盐湖等多种多样化的地貌景观,决定了旱生、超旱生、沙生、盐生、湿生和水生等多种植物群落的形成,生物资源多样性丰富[15]。艾比湖湿地自然保护区的典型植被群落有梭梭〔Haloxylon ammodendron(C.A.Mey.)Bunge〕、芦苇〔Phragmites australis(Cav.)Trin.ex Steud.〕、胡杨(Populus euphraticaO liv.)、柽柳(TamarixchinensisLour.)、甘草(GlycyrrhizauralensisFisch.)、小獐毛 〔Aeluropus pungens(M.Bieb.)C.Koch〕、盐穗木(Halostachys caspicaC.A.Mey.ex Schrenk)、盐节木〔Halocnemum strobilaceum(Pall.)M.Bieb.〕、碱蓬〔Suaedaglauca(Bunge)Bunge〕、枇杷柴 〔Reaumuriasongarica(Pall.)Maxim.〕和盐爪爪〔Kalidiumfoliatum(Pall.) Moq.〕等[15-16]。本研究选取的样地以盐生生境为主,少数伴随荒漠生境和沙漠生境。
1.2 样方设置和数据调查
根据艾比湖湿地典型群落的分类,于2007年7月至9月在艾比湖湿地自然保护区东大桥和鸭子湾管护站附近设置面积10m×10m的样方17个,在其内随机划定面积5m×5m的样方21个,分别调查每个样方中乔灌木的种类、多度、盖度、高度、胸径、冠幅和物候相,同时测定并记录各样地的海拔、经度、纬度、种群微环境和地理地貌。乔灌木种类主要为胡杨、盐节木、多枝柽柳(TamarixramosissimaLedeb.)、盐穗木、枇杷柴、白梭梭(HaloxylonpersicumBunge ex Boiss.etBuhse)、盐豆木〔Halimodendronhalodendron(Pall.)Voss〕和梭梭。
1.3 采用的评价指标及计算方法
根据调查样方中各种类的多度,在该样方中出现记为“1”,不出现记为“0”,然后据此进行进一步的数据计算和处理。

以VR值作为全部种的关联指数。若VR=1,表明8个乔灌木种群间不关联;若VR>1,表明8个乔灌木种群间表现出净的正关联;若VR<1,表明8个乔灌木种群间表现出净的负关联。用统计量W来检验VR值偏离1的显著性,W的计算公式为:W=VR×N。若被调查的8个乔灌木种群间不相关联,则W值落入区间内的概率为90%。
1.3.2 种群聚集强度分析 采用负二项参数(K)[18]、扩散系数(C)[19-20]、平均拥挤度(m*)[21]和Cassie指标(CA)[22]等4个指标分析不同种群的聚集强度和分布格局。以S2表示单种群的方差表示单种群的均值、n为样方数,则4个指标的计算公式如下:
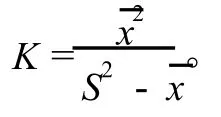
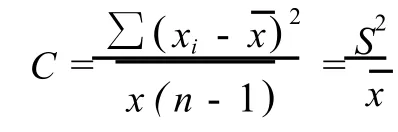
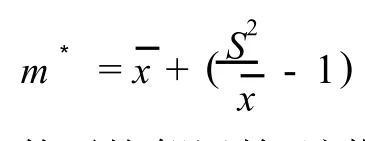
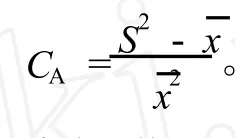
2 结果和分析
2.1 不同面积样方中8个乔灌木种群间的总体联结性比较
在群落中,植物种对间的正关联主要是由于它们具有相近的生物学特性、对生境具有相似的生态适应性和相互重叠的生态位所致;种对间的负关联则主要是由于它们具有不同的生物学特性以及对生境资源的竞争促使生态位相互分离所致[24]。艾比湖湿地自然保护区不同面积样方中乔灌木总体联结性的比较结果见表1。
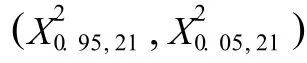

表1 艾比湖湿地自然保护区不同面积样方中乔灌木种群间总体联结性比较Table1 Com par ison of genera l associa tion am ong arbor-shrub popu la tion s in quadra ts with d ifferen t areas of Eb inur LakeWetland Na ture Reserve
2.2 不同面积样方中8个乔灌木种群空间分布格局的分析
艾比湖湿地自然保护区不同面积样方中8个乔灌木种群空间分布格局的分析结果见表2。由表2可以看出:在面积5m×5m的样方中,胡杨、枇杷柴、盐节木、白梭梭、盐豆木和盐穗木种群的负二项参数(K)均大于0、扩散系数(C)均大于1、Cassie指标(CA)均大于0;多枝柽柳种群的K值小于0、C值小于1、CA值小于0;梭梭种群的K值趋于无穷大、C值等于1、CA值等于0。表明在5m×5m的尺度上,胡杨、枇杷柴、盐节木、白梭梭、盐豆木和盐穗木种群的空间分布格局为聚集分布,多枝柽柳种群的空间分布格局为均匀分布,而梭梭种群的空间分布格局为随机分布。对C值进一步的t检验结果表明,在8个乔灌木种群中,只有枇杷柴和盐节木种群的空间分布格局为极显著的聚集分布,而其他6个种群的空间分布格局均为不显著的分布类型。
从表2还可以看出:在面积10m×10m的样方中,8个种群的K值均大于0、C值均大于1、CA值均大于0,表明在10m×10m的尺度上,8个种群的空间分布格局都为聚集分布。对C值进一步的t检验结果显示,8个种群的空间分布格局均为极显著的聚集分布。
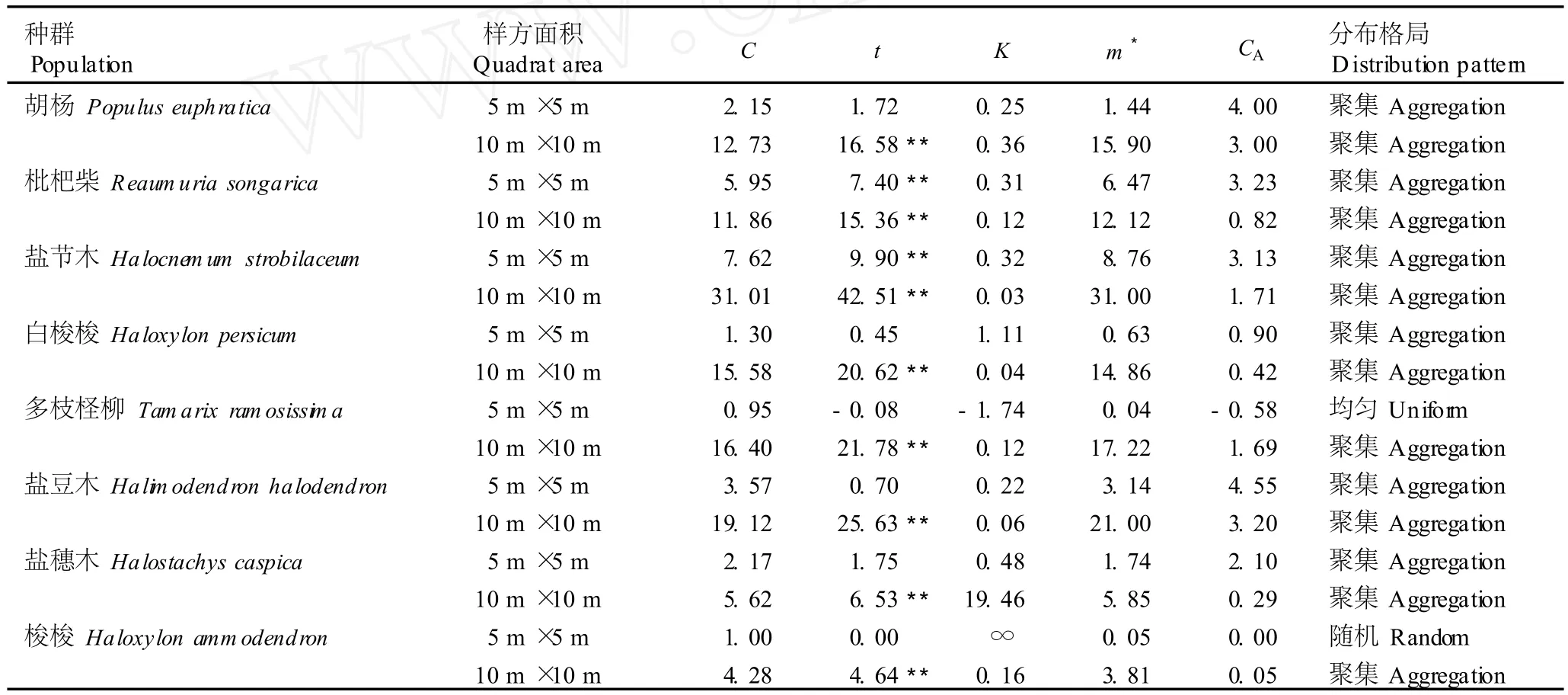
表2 艾比湖湿地自然保护区不同面积样方中8个乔灌木种群空间分布格局的分析1)Table2 Ana lysis on spa tia l d istr ibu tion pa ttern s of eigh t arbor-shrub popu la tion s in quad ra ts with d ifferen t areas of Eb inur LakeWetland Na ture Reserve1)
植物种群的样方面积从5m×5m扩大至10m×10m,若个体的平均拥挤度(m*)的排序号不变,则m*的变化率为0;若m*的排序号向后移动,则m*的变化率取正,每移动1个单位,m*值增加1;若m*的排序号向前移动,则m*的变化率取负,每移动1个单位,m*值减少1。从m*排序号的变化中可以看出:随样方面积的变化,不同植物种群内的密度变化是不同的,也就是说,当种间出现对资源的竞争时,种间竞争能力不强,种内自疏作用不同。
受种群密度的影响,m*表征种群空间分布(即样方之间)的差异以及单位样方内种群的数量和密集程度。艾比湖湿地自然保护区不同面积样方中8个乔灌木种群m*的分析结果显示(表2):在面积5m×5m的样方内,8个乔灌木种群按m*值由大至小依次排序为盐节木、枇杷柴、盐豆木、盐穗木、胡杨、白梭梭、梭梭、多枝柽柳,当样方面积增加至10m×10m,8个乔灌木种群的排序改变为盐节木、盐豆木、多枝柽柳、胡杨、白梭梭、枇杷柴、盐穗木、梭梭。由此看出,样方面积从5m×5m扩大至10m×10m,枇杷柴、盐穗木和梭梭种群的m*排序号分别向后移动4、3和1位,m*的变化率分别为4、3和1;盐节木种群的m*排序号不变,m*的变化率为0;白梭梭、胡杨、盐豆木和多枝柽柳种群的m*排序号分别向前移动了1、1、1和5位,m*的变化率分别为-1、-1、-1和-5。
3 讨论和结论
在种群空间分布格局的研究中,采用不同的取样单位会得出不同的结论[1],因此,在分析种群空间分布格局的过程中,为了避免取样面积较小造成的随机分布,选择合适的取样单位非常关键。例如,在本研究中,在面积5m×5m的样方内,梭梭种群多度小,得出其空间分布格局为随机分布的结论,与野生梭梭种群常呈现聚集分布的空间分布格局不相符,而当样方面积增大至10m×10m时,梭梭种群的空间分布格局又趋向于聚集分布。t检验结果显示,在艾比湖湿地自然保护区内,在面积5m×5m的样方中,8种乔灌木种群中仅枇杷柴和盐节木种群的空间分布格局类型为极显著的聚集分布;而在面积10m×10m的样方中,8个种群的空间分布格局均为极显著的聚集分布,表明采用5m×5m的样方面积测量乔灌木种群空间分布格局的误差较大,而当样方面积增加至10m×10m,t检验的显著性增加。因此,在艾比湖湿地自然保护区的乔灌木种群分布格局的研究中,采用10m×10m的样方面积是相对合适的。
种群空间分布格局是种群生态生物学特征的重要方面,是种群本身生物学特性、种间关系和环境条件综合作用的结果[2]。在环境均一、生存条件良好的情况下,以母株为中心的繁殖方式可使低龄子代呈现聚集分布[25]。在实际的野外调查数据中,面积5 m ×5m的样方内中、小幼苗多,成株较少。在艾比湖湿地自然保护区面积5m×5 m的样方中,在8个乔灌木种群种间总体关联性为显著性的正联结(物种间很少存在资源的限制)条件下,有6个种群表现为聚集分布,1个种群表现为均匀分布。虽然8个乔灌木种群总体上很少存在对资源的竞争,但各个种群对空间的侵占和扩展程度是不同的,可以用负二项参数(K)作为衡量指标[26]。
艾比湖湿地自然保护区8个乔灌木种群的K值表明:在5m×5m的尺度上,白梭梭种群有最大的资源利用力,对空间的扩展能力强;灌木层植物的竞争性以及对环境的占据能力强于乔木,如枇杷柴、盐节木和盐穗木的K值均大于胡杨。胡杨虽然为艾比湖湿地自然保护区的建群种,但在5m×5 m的小尺度上,胡杨种群的竞争性和对资源的适用性较差,这是由于胡杨种群在小面积的样方内中、小幼苗的竞争能力较弱所致。
马万里等[1]、何恒斌等[9]和郭华等[27]的研究结果表明,在植物个体由小到大的生长过程中,由于对环境资源的需求增加,种内竞争增强,导致不同个体间产生分化,群落自疏,使种群的聚集性减弱、随机性增强。在艾比湖湿地自然保护区内,样方面积从5m×5m扩大至10m×10m,8个乔灌木种群总体从显著正联结转变为不显著负联结,从只有2个种群呈极显著聚集分布到8个种群全部呈极显著聚集分布,说明随着种群年龄的增加,种群内部开始出现竞争,但这种竞争不太激烈,还没有达到使种群自疏至均匀分布或随机分布的程度,即表明种群中的物种是稳定共存的。
不同于面积5m×5m的样方,在面积10m×10 m的样方内,胡杨种群的K值在8个乔灌木种群中较高,其值为0.36,仅小于盐穗木种群。说明随着种群年龄的增加,胡杨种群在样方中的竞争能力和对环境的适用性是持续增加的,也验证了胡杨在整个群落中处于建群种地位。
根据上述研究结果可得出以下结论:1)在面积5m×5m的中、小幼苗样方上,艾比湖湿地自然保护区的8个乔灌木种群总体间表现为显著正联结,物种间很少存在对资源的竞争;胡杨、枇杷柴、盐节木、白梭梭、盐豆木和盐穗木种群的空间分布格局为聚集分布,多枝柽柳种群为均匀分布,梭梭种群为随机分布。2)在面积10m×10m的成株样方上,8个乔灌木种群总体间表现为不显著的负联结,物种间存在对资源的竞争,但稳定共存,全部表现为极显著的聚集分布;在进行艾比湖湿地自然保护区种群空间分布格局的研究中,采用10m×10m的取样面积是相对合适的。3)随着样方面积的增加,艾比湖湿地自然保护区内的8个乔灌木种群的种间竞争能力和种内自疏能力有所变化,枇杷柴、盐穗木和梭梭的总体表现较强,多枝柽柳最差。
[1]马万里,荆 涛,罗菊春,等.长白山林区核桃楸种群分布格局研究[J].内蒙古师范大学学报:自然科学汉文版,2008,37(2):233-236.
[2]郭忠玲,马元丹,郑金萍,等.长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究 [J].应用生态学报,2004,15(11):2013-2018.
[3]张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[4]于传宗,慕宗杰,特日格勒.植物种群空间分布格局的研究方法[J].畜牧与饲料科学,2008(5):40-42.
[5]兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-21.
[6]张金屯.数量生态学[M].北京:科学出版社,2004:243-287.
[7]赵峰侠,尹林克.荒漠内陆河岸胡杨和多枝柽柳幼苗种群空间分布格局及种间关联性[J].生态学杂志,2007,26(7):972-977.
[8]李宗菊,张微思,张志国,等.鹅膏属种群分布格局研究[J].中国食用菌,2007,26(3):6-9.
[9]何恒斌,张惠娟,贾桂霞.磴口县沙冬青种群结构和空间分布格局的研究[J].林业科学,2006,42(10):13-18.
[10]杨君珑,王 辉,王 彬,等.子午岭油松林灌木层主要树种的空间分布格局和种间关联性研究[J].西北植物学报,2007,27(4):791-796.
[11]程 煜,闫淑君,洪 伟,等.檫树群落主要树种分布格局及其动态分析[J].植物资源与环境学报,2003,12(1):32-37.
[12]范海兰,洪 伟,吴承祯,等.福建大田栲树次生林优势种群结构与动态[J].植物资源与环境学报,2004,13(3):44-49.
[13]李艳红,楚新正,金海龙.新疆艾比湖流域水文特征分析[J].水文,2006,26(5):68-71.
[14]吴敬禄,刘建军,王苏民.近1500年来新疆艾比湖同位素记录的气候环境演化特征[J].第四纪研究,2004,24(5):585-590.
[15]杨晓东,吕光辉,田幼华,等.新疆艾比湖湿地自然保护区植物的生态分组[J].生态学杂志,2009,28(12):2489-2494.
[16]陈蜀江,侯 平,李文华,等.新疆艾比湖湿地自然保护区综合科学考察[M].乌鲁木齐:新疆科学技术出版社,2006:15-36.
[17]杜道林,刘玉成,李 睿.缙云山亚热带栲树林优势种群间联结性研究[J].植物生态学报,1995,19(2):149-157.
[18]徐 坤,谢应忠,李世忠.宁南黄土丘陵区退化草地群落主要植物种群空间分布格局对比研究[J].西北农业学报,2006,15(5):123-127.
[19]C lark P J,Evans FC.D istance to nearestneighborasameasure of spatial relationship s in populations[J].Eco logy,1954,35(4):445-453.
[20]Greig-Sm ith P.Pattern in vegetation[J].Journal of Eco logy,1979,67:755-779.
[21]L loyd M.Mean crow ding[J].Journal of Anim al Ecology,1967,36:1-30.
[22]Cassie R M.Frequency distribution models in the ecology of p lankton and other organism[J].Journal of Animal Eco logy,1962,31:65-95.
[23]王新功,洪 伟,吴承祯,等.武夷山米槠林群落优势种群空间格局的研究[J].河南农业大学学报,2003,37(1):44-48.
[24]郭志华,卓正大,陈 洁,等.庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J].植物生态学报,1997,21(5):424-432.
[25]郑元润.大青沟森林植物群落主要木本植物种群分布格局及动态的研究[J].植物学通报,1998,15(6):52-58.
[26]张继义,赵哈林.科尔沁沙地草地植被恢复演替进程中群落优势种群空间分布格局研究[J].生态学杂志,2004,23(2):1-6.
[27]郭 华,王孝安,肖娅萍.秦岭太白红杉种群空间分布格局动态及分形特征研究[J].应用生态学报,2005,16(2):227-232.
猜你喜欢
哈哈画报(2022年4期)2022-04-19 11:11:54
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
科学大众(2020年17期)2020-10-27 02:48:48
水利规划与设计(2020年6期)2020-06-23 11:41:48
林业调查规划(2020年3期)2020-06-03 07:02:36
中学生天地(B版)(2019年4期)2019-05-08 03:22:54
广东农业科学(2017年9期)2018-01-16 18:38:40
绿色中国(2015年6期)2015-07-03 01:13:10
