
下沙新城区城市化过程中不同生境斑块荠菜种群数量特征及生活史变异
2010-11-23 00:53:47黄瑞建张岳恒
杭州师范大学学报(自然科学版) 2010年6期
黄瑞建,张岳恒,陈 波
(杭州师范大学 生命与环境科学学院,浙江 杭州 310036)
加快城市化进程,是我国走向现代化的必由之路,但城市化过程中对土地的大量需求引起土地利用的严重矛盾.滩涂作为我国重要的后备土地资源,对解决这一矛盾起到了重要的作用.然而,这种土地利用方式造成了生境破碎化,所形成的众多生境岛屿[1],加之各类人工绿地,组成了城市逆境中特殊的斑块环境.下沙新城区位于杭州经济开发区内,是典型的滩涂开垦地.该区在目前的城市化扩张过程中,原有的滩涂环境发生改变,取而代之的是大片的人工绿地(大多以草坪和模型色块为主)和许多零星分散的废弃地(闲置地),生境由均质走向异质.
城市化的进程改变着城市植被的生境——大量的地表被水泥地和建筑物覆盖,气候因子也发生改变.随之而来的是城市生物尤其是城市植物种类的变化,如新城区常出现大量的伴人植物.伴人植物常被定义为靠自身的适应及竞争能力而在人为活动干扰过的生境中得以繁衍的一类植物[2].这些植物由于适应城市逆境常得到迅速发展,并对环境产生重要影响,具有耐干旱、贫瘠、污染以及抗干扰等特征,因此被认为是城市逆境中的先锋植物.城市伴人植物不但构筑了城市地区物种、群落、生态系统以及景观多样性的基础,而且具有指示环境的作用,可在一定程度上反映人为干扰程度[3].
不同的生境条件常常是导致植物种群生活史变异的外界因素.虽然植物的生活史受到种的遗传而表现出相对稳定性,但有时外界因子所起的塑造作用更为重要[4].植物种群的生活史,一般用生活史特征来描述.生境的不同,导致了植物种群的可塑性生长,并表现为外部数量特征以及繁殖上的变异.
下沙特殊的斑块环境中同样也分布着大量伴人植物,许多种类如荠菜(Capsellabursapastoris)、繁缕(Stellariamedia)等常在早春季节大量繁生.这类杂草一般靠城建用料以及城市绿化无意带进,其种群何以能在这些斑块生境中生存?其动态如何?生活史如何?与脆弱的斑块环境有着什么样的联系?该研究试图分析新城区典型的早春伴人杂草——荠菜的种群动态、繁殖分配和生活史特征,以揭示这类植物成功入侵城市斑块生境的机制,以及与这些生境的关系.
1 研究区概况
根据陈征海[5]对浙江天然湿地的分类,下沙新城区原先应属于钱塘江典型的泛洪平原湿地,是由钱塘江冲刷而成的滩涂地.经过高强度的开垦后,目前下沙大部分已建成居民区、公路、校园、公园等场所,成为杭州的三大副城之一,是以杭州经济技术开发区和高教园区为骨干的综合性新城区.新城区所在地年均气温16~19 ℃,年均降雨1 100~1 200 mm,年均湿度80%~85%.其土壤为潮土化盐土,母质为近期浅海相沉积物,pH为7.7~8.0,含盐量0.1%~0.4%,质地为沙壤土,以粗粉沙为主(80%),粘粒含量较少(10%),一些耐碱性、耐盐性的植物分布较多.另外,城区周围无山脉和大面积防护林,受风的影响较大.
研究区内典型的早春杂草种类有加拿大一枝黄花(SolidagoCanadensis)、鼠麴草(Gnaphaliumaffine)、一年蓬(Erigeronannuus)、青蒿(Artemisiacaruifolia)、小蓬草(ConyzaCanadensis)、泥胡菜(Hemisteptalyrata)、酸模叶蓼(Polygonumlapathifolium)、杠板归(Polygonumperfoliatum)、齿果酸模(Rumexdentatus)、马唐(Digitariahall)、棒头草(Polypogonfugax)、通泉草(Mazusjaponicus)、婆婆纳(Veronicadidyma)、葎草(Humulusscandens)、大巢菜(Viciasativa)、田菁(Sesbaniacannabina)、荠菜、碎米荠(Cardaminehirsute)、藜(Chenopodiumalbum)、繁缕、节节草(Hippochaeteramosissima)、野老鹳草(Geraniumcarolinianum)、石龙芮(Ranunculussceleratus)、井栏边草(Pterismultifida)、龙葵(Solanumnigrum)、酢浆草(Oxaliscorniculata)、空心莲子草(Alternantheraphiloxeroides)、乌蔹莓(Cayratiajaponica)等,许多种类如荠菜、繁缕、井栏边草等为典型的伴人植物.
2 材料与方法
研究以野生荠菜为实验对象.该植物是典型的早春伴人植物,为一或二年生草本,一般花期为3~4月,果期6~7月[6].该种生长势及适应能力很强,能耐旱、脊薄等恶劣条件,在下沙新城区的斑块生境中已大量出现.该实验主要采用野外调查和实验室分析相结合的方法进行研究.
2.1 野外固定样地设置
下沙新城区土地基本建成学校、公园、公路等,另外零星分散着未开发的荒野地,该研究以杭州师范大学为典型地(可代表下沙校园生境),在校园内选择9个生境取样,在校园外附近选择2个生境取样,各生境的具体位置及特点见表1.
2.2 实验室内分析
于2006年4月与5月(重要生长期或繁殖期)对样地中的植物种群进行连续观察,并于月初各采集一次样品进行分析.该实验采用收获法在所设样地中各选取约15株大小整齐的荠菜,分别测定其主根长(cm)、株高(cm)、单叶长(cm)宽(cm)重(g)、各构件干重生物量(g)(放置在85 ℃的烘箱内烘48 h至恒重后,用Mettler Toledo AL204型电子天平测定)、生殖分配(RA%)(用公式RA=繁殖构件生物量/总生物量计算)等参数;同时在各个样地中随机选取大小一致的5株荠菜叶片(每株2片),测定叶绿素相对含量(CCI)(CCM-200手持式叶绿素仪);同步测定各小生境内的生态因子,主要包括大气温(℃)湿度(RH%)(HI93640便携式温湿度计)、光量子(μmol·m-2·s-1)(BQM光量子计)、土壤pH(HI98128酸度计)和含水率(%)(烘干法).
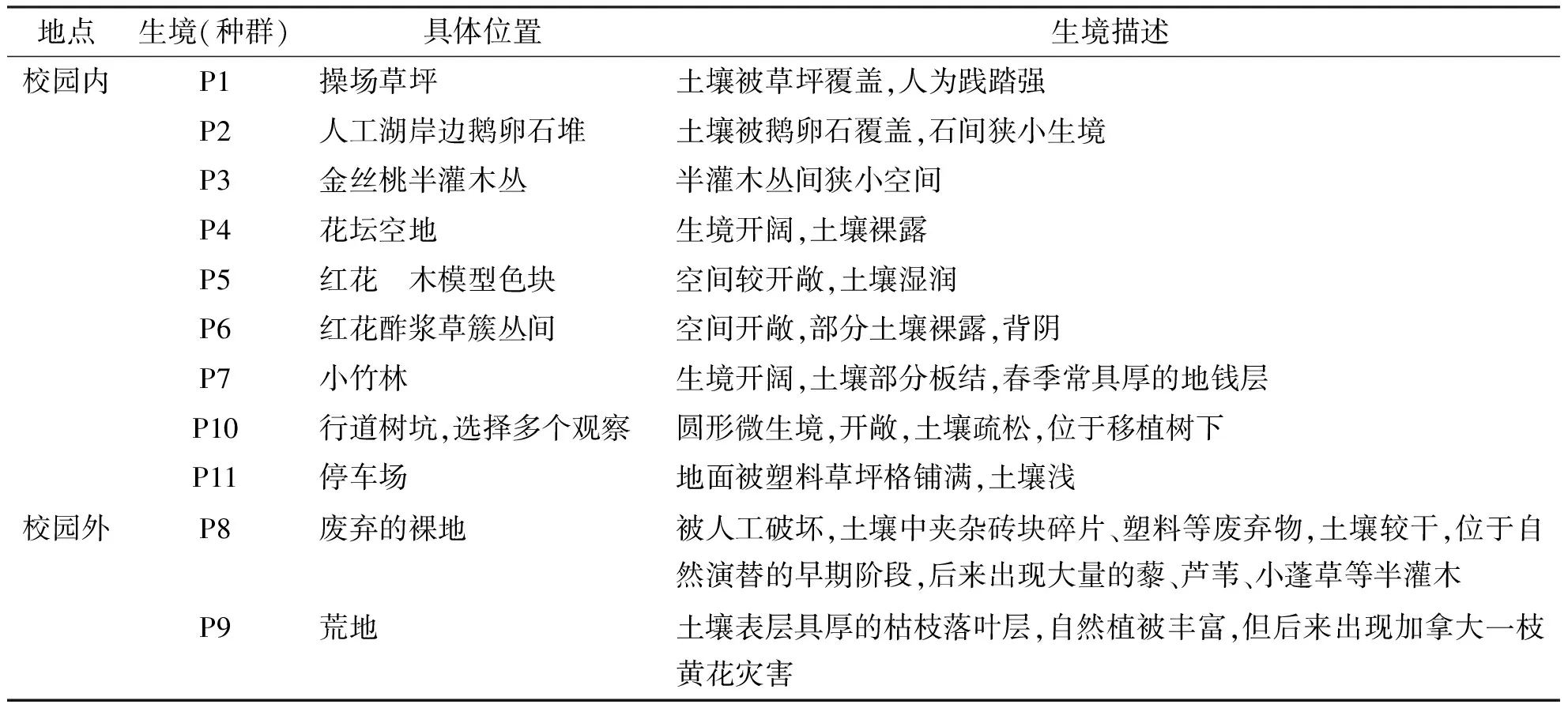
表1 取样生境具体位置及生境描述
2.3 数据分析
结合SPSS和Excel软件对数据进行转化和处理.用单因素方差分析、非参数分析、独立样本t检验、LSD和Duncan多重比较等统计学方法分析各类环境因子与荠菜种群数量特征的时间和空间差异.用Spearson相关系数法检验叶重与叶长、叶宽以及生殖分配与多种环境因子的相关性,并运用多元线性回归法模拟方程.利用SPSS统计软件绘制荠菜种群生殖分配柱形图,并依据4月份的生殖分配将各种生境进行分层聚类,绘制聚类树形图.
3 结果与分析
3.1 环境因子的时空格局
研究区域的微生境变化体现出了明显的时空差异:4、5月份晴朗中午各生境的平均气温、相对湿度、光量子的变化因微生境的特殊而有所不同(p<0.05);空间水平上,生境土壤pH的变化范围为7.0~8.5,含水率为23.7%~35.3%.
3.2 荠菜种群主要数量性状的变异
同一时间不同生境及同一生境不同时间的荠菜种群发生了表型上的变异(表2),体现出时空差异.

表2 4、5月份不同种群性状的变异
3.2.1 主根长、株高、株生物量的变异
3.2.2 叶构件的生境和月份差异
4月份的荠菜种群单叶长、宽、重均达到了极显著的水平(X2=226.075,173.214,77.302,p<0.01),其中P5的叶长、P3的叶宽、P9的叶重显著高于其他生境,P2的叶长、叶重及P4的叶宽在各生境中最低.5月份的3项指标也达到了极显著的差异(X2=106.486,102.246,77.302,p<0.01),P9的叶长和叶宽、P10的叶重在各生境中最大,P11的3项指标均最低.月份间3种指标的变异系数也有很大差别,4月份分别为30.4%~109.3%,38.3%~99.0%,56.1%~171.6%,5月份为29.2%~46.7%,34.7~71.0%,71.0%~152.2%.相关分析表明叶重和叶长、叶宽具有极显著的相关,其Spearson相关系数R分别为0.902,0.809和0.749,0.778(p<0.01);但叶重和长宽比在各种群间却普遍不存在相关(P>0.05).荠菜叶重与叶长、叶宽的回归方程为:W=-0.006 5+0.004 6L+0.001 7B,其中W为叶重(g),L为叶长(cm),B为叶宽(cm).
4月份测定的一组叶绿素相对含量也存在着生境差异(p<0.01),P8的平均含量为(21.084±6.71)CCI,在生境中居首位,Duncan多重比较发现该种群与其他种群均存在显著差异(p<0.05).
3.3 生殖分配调节的生活史变异
生殖分配是种群生活史的一个重要内容,它是一定时间下植物个体权衡生殖和营养功能的一种表现[7].生活史一旦发生变异,其繁殖时间和物候期就发生改变,最终导致同一时间水平上种群生殖分配的改变.
3.3.1 依赖物候期的生殖配置
草本植物的生殖分配是将植物干重区分为生殖部分和非生殖部分,对荠菜而言其生殖分配的实质就是其伞房花序和果实占株生物量的比例,即繁殖构件的分配比.表3是种群在同一月份所处的各个物候期,图1是4、5月各生境荠菜营养构件(根、茎、叶)和繁殖构件的相对分配比.总体来看,不论是4月还是5月,营养构件的比例始终高于繁殖构件.4月大多种群处在花果共存期,种群生殖分配的空间波动为20%~30%,但P2的生殖分配高达40%,而P10只有15.3%.5月,各种群转入另个物候期.与4月相比,P1,P4,P6的生殖分配明显下降,P1由27.7%降到5.6%,在各生境中处于最低;P10的生殖分配有所升高;P3,P9的生殖分配只下降了0.3%,0.7%;P11于5月才开始进入花期,其生殖分配为24.0%;P8的生殖分配达到了41.6%.
通过人机交互的技术将抽象的数据表现为直观可视的图形或图像,加强了用户对于数据的理解能力,使得复杂的数据更加容易被人们所理解。
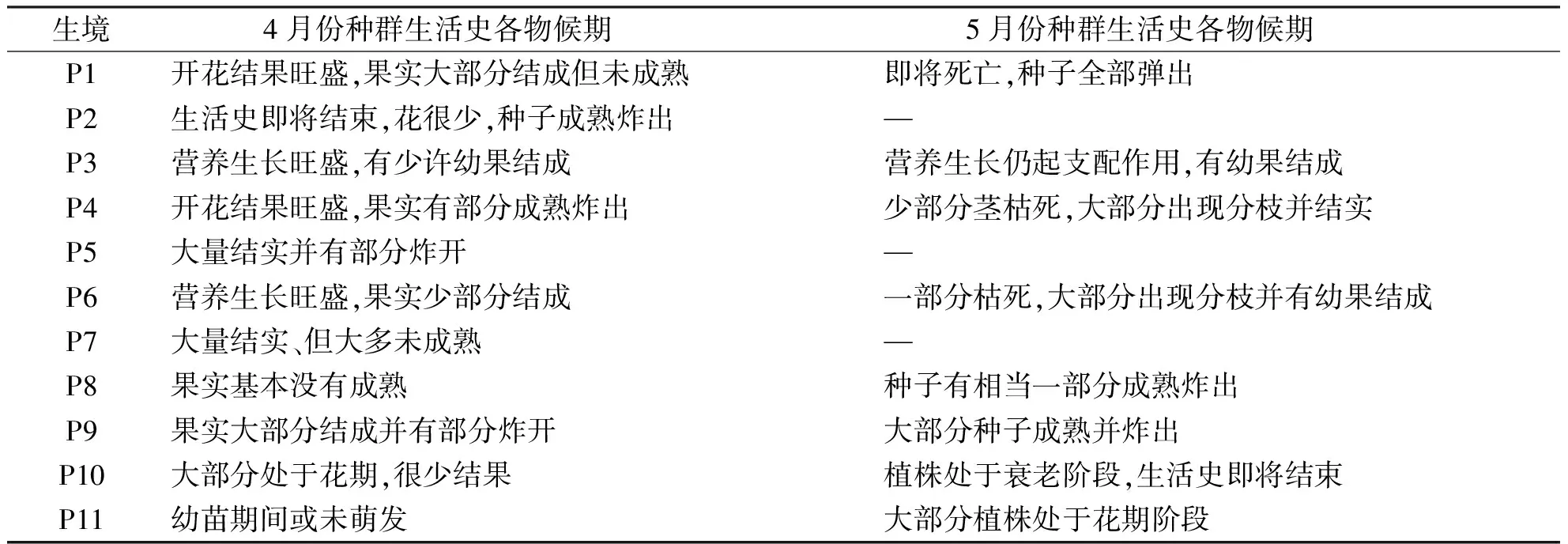
表3 4、5月份不同种群的物候阶段
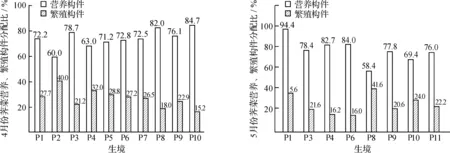
图1 荠菜种群的繁殖分配Fig. 1 Reproduction allocation of Capsella bursapastoris population
3.3.2 生殖分配与环境因子的关系
同一时间上种群生殖分配的可塑性是种群对外界多种因子综合作用的灵敏响应.利用4月份的一组数据模拟生殖分配与多个环境因子的多元线性回归方程:
RA=-1.87+0.044t+0.024R-0.000 031L-0.046pH+0.71C
其中,RA为繁殖分配(%),t为大气温度(℃),R为大气湿度(RH%),L为光量子(μmol·m-2·s-1),C为土壤含水率(%).
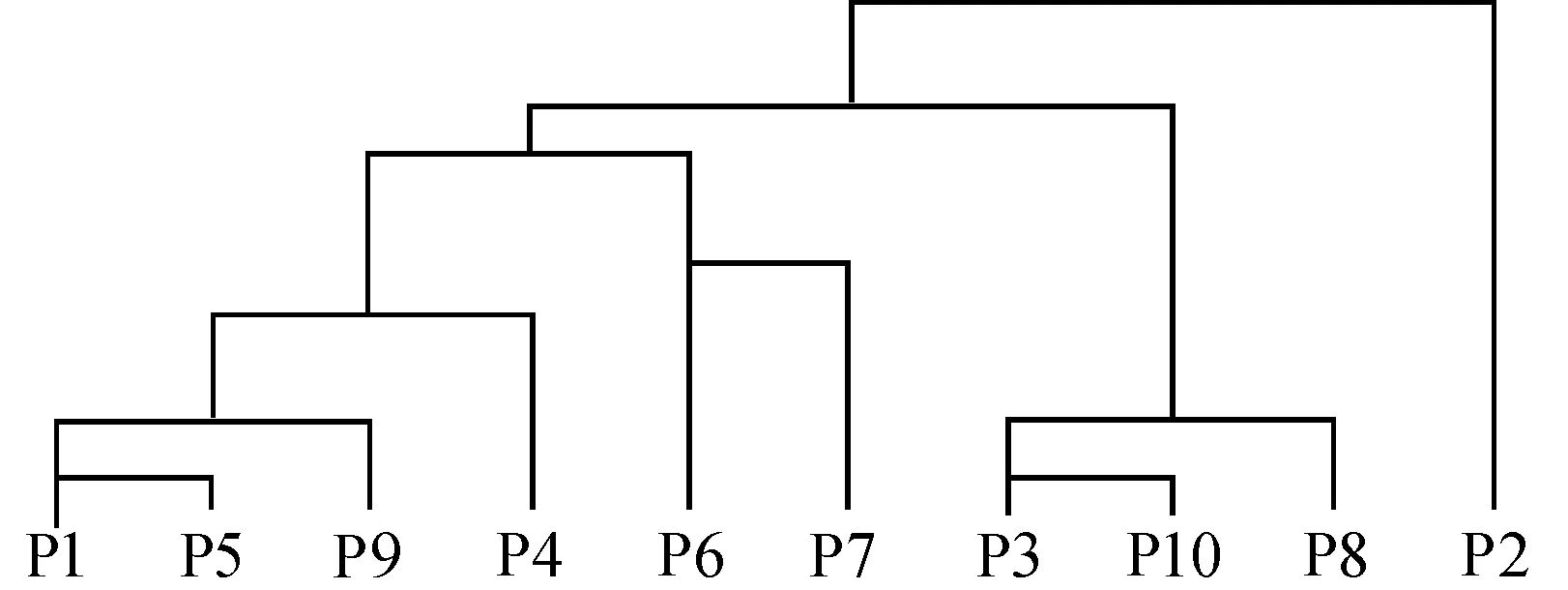
图2 生境聚类树形图Fig. 2 Dendrogram of cluster analysis of habitats
3.3.3 依据荠菜繁殖分配的生境分类
利用分层聚类的统计学方法并依据4月份荠菜种群的生殖分配绘制聚类树形图(图2),将10个生境分为3类:第一类包含P1,P5,P9,P4,P6,P7,该类生境中种群的生殖分配处于23.9%~32.0%之间,物候为花果共存期;第二类包含P3,P10,P8,繁殖分配在15.3%~21.3%之间,这些种群处于盛花期;第三类只包含一个生境,即P2,生殖分配可达40%,种群到达盛果期.
4 讨 论
下沙新城区在城建过程中形成了诸如杭州师范大学校园等类似生境,因此在一定程度上可认为该校园的校内及校外生境能代表整个下沙生境水平.这些斑块生境在城市化过程中形成,其环境是极其复杂多变的,气温、相对湿度、光量子、土壤pH和含水率等外界变量都有可能造成微生境差异.早春季节,伴人植物荠菜大量生长,以其作为优势种的种群一般由去年秋季萌发的植株和本年刚出土的植株共同组成.2、3月初,这些植株就有可能完成营养生长而进入盛花期,但该期持续时间不长,花序呈伞房状;荠菜花一凋谢便结成幼果,伞房花序伸长成总状花序,由于花序的无限伸长,盛花期结束并不意味着花期的结束,而是转入到花果共存期,且持续时间长;到了5、6月份,花期结束过渡到盛果期,这时植物营养体接近死亡,短角果大量生成,持续时间短;果实成熟后将大量种子弹出导致植株的死亡.下沙新城区特殊斑块内的生态因子以及人为干扰是极其不稳定的,生活于其中的物种对这种生境或多或少都会表现出一定的反应[7-8].荠菜对该类生境的响应往往体现在表型数量特征的变异及对以上生活周期(主要在繁殖方面)的调节上.
4.1 荠菜种群数量特征的变化
数量性状的变异对种群而言最为直接.当微生境不同时,主根长、株高、株生物量、叶形状等在生境间有巨大差异:如4月份,P5所在生境气温最低(荠菜喜冷凉气候)、湿度最大(荠菜喜湿润气候)、所接纳的光量子最多、土壤含水率最高,则相应的主根长、株高、株生物量就达到最高.月份间由于斑块内种群的动态,个别数量性状也可能有一些变化:如P10的主根长和株高具有月份间差异,而株生物量却没有这种差异;但此时P4的3项指标都有时间上的差异.变异系数的比较证明株生物量不论在时间还是空间上都具有更大的可塑性,因此在数量特征中具有更大的变异潜能.叶的变化极其复杂,但主要表现在叶长、叶宽、叶重、叶形状以及叶绿素等指标上,这些指标在月份和生境间均有显著差异.变异系数证明了生境之间存在大的波动,这可能是荠菜的生理异型叶(基生叶和茎生叶、羽状叶和全缘叶)造成的.叶长、叶宽、叶重构成了一定的叶形状,直接影响其变异系数.变异系数小的种群叶型比较整齐,即以一种叶型为主,而变异系数大的种群叶型则较为混杂.叶型的丰富变化与环境直接相关,如4月份P2的叶长、叶宽、叶重的变异系数分别为30.41%,42.84%,87.10%,在所有种群中最低.该类生境相对恶劣,干扰大,种群具有小植株性状,其叶形结构小,这些小叶趋于同一,从而使各类指标的变异系数变小.
4.2 生殖分配调节的生活史变异
繁殖分配是生活史对策的另一个重要方面,是指植物个体在特定时间内用于生殖的净同化产物占总产物的比例[7,9-10].植物在不同的生境下,可通过权衡营养与生殖功能的相对资源配置而提高适合度[10-12],从而演变出最佳生殖对策并成功繁殖.图3所显示的营养体比例始终大于繁殖体的现象说明荠菜种群的营养分配要优于生殖分配.出现这种现象的原因是:尽管生殖在维持种群繁衍方面极其重要,但只有生存才能保证这种活动的进行,因此对营养体的绝对投入具有积极意义.虽然繁殖体在这方面不及营养体重要,但在生殖时仍可能占据一定的比例,如花果共存期常在20%~30%之间,成为高生殖分配的植物,这种现象可用一维选择对策[13-14]中的r-选择理论来解释:在新环境及暂时生境中,特殊种群可能大量出现并繁衍,但也容易受环境干扰而突然灭绝,只有高生殖分配的类群才有可能产生大量后代并靠机遇而存活下来.Grime的杂草对策[13,15]同样也可以解释荠菜的生殖对策:在干扰强烈的生境中往往会发展出潜在生产力高的植物种群.荠菜种群生活于城市逆境的高干扰生境中,资源转向种子的生产,则对应的生殖分配往往就被提高.
生活史理论假说[16]认为,在不利的环境中植物生殖早,但个体小.该假说同样适合荠菜种群:如P2所处的生境具有很高的死亡危险,其种群在3月就进入繁殖期,4月进入生活史的结束阶段,种子大量散布,且最终的个体最小;而比P2优越的其他生境中,4月荠菜一般刚进入繁殖期,并处于花果共存阶段,但个体明显比P2大(如P8).该假说还认为营养生长与生殖存在负相关,这点也普遍存在于荠菜种群中.营养构件比例越大,则生殖分配越低(图3),如P2种群在4月初就已经停止生长,接近生活史的死亡阶段,表现为基生叶的枯萎脱落(叶构件只占8.3%),花期结束,果实大量成熟(但种子未弹出),从而使生殖分配高达40%,在所有生境类型中处于最高;而在同一时间P10才刚进入花期,生殖分配仅为15.3%,在这之前植株一直处于营养生长,物质和能量明显积累在叶中(叶构件高达45.8%).因此在不同生境条件下,同一时间各种群所处的物候期不一致,则相应的生殖分配就表现出不同.这些现象是种群对微环境生态因子的灵敏反映[17-19],具有很大的生态可塑性.另外,5月再次测定种群的生殖分配时发现,这些值与4月相比有明显的变化:有些种群下降了,但另一些却升高,还有的保持基本不变.对这种现象可能的解释是:经过一个月的种群动态,种群可能进入或不进入另个物候期,并通过营养与生殖功能的再次权衡而重新划分营养构件与生殖构件的分配比.
4.3 种群数量特征、生活史与斑块生境的联系
荠菜种群的数量特征以及生活史的变异具有一定的生物学和生态学意义,体现了斑块水平上种群内部的微小进化[4].同时,在城市斑块水平上种群的这些变异可在一定程度上反映城市化过程中的环境变化和人为干扰,因此可作为指示城市环境的一种优良伴人植物,如根据针对4月荠菜种群的生殖分配而进行的聚类分析,可推测出下沙新城区在城市化进程中斑块生境的异质程度和干扰程度,该文对此仅进行了初步的分析.
[1] 马克明,傅伯杰.北京东灵山地区景观格局及破碎化评价[J].植物生态学报,2000,24(3):320-326.
[2] 蒋高明.城市中的伴人植物[J].植物学通报,1989,6(2):116-120.
[3] 蒋高明.城市植被:特点、类型与功能[J].植物学通报,1993,10(3):21-27.
[4] 钟章成,曾波.植物种群生态研究进展[J].西南师范大学学报:自然科学版,2001,26(2):230-236.
[5] 陈征海.浙江天然湿地类型研究[J].浙江大学学报:农业与生命科学版,2002,28(2):156-160.
[6] 韦制直,何业祺.浙江植物志第三卷[M].杭州:浙江科学技术出版社,1993:36-37.
[7] 操国兴,谢德体,钟章成,等.植物种群的生殖分配[J].四川林业科技,2003,24(2):25-29.
[8] 覃凤飞,安树青,卓元午,等.景观破碎化对植物种群的影响[J].生态学杂志,2003,22(3):43-48.
[9] 郭力华,杨允菲,李建东.松嫩平原不同生境寸草苔种群生殖分株的数量特征及生殖分配动态[J].草业学报,2005,14(2):63-68.
[10] 张景光,王新平,李新荣,等.荒漠植物生活史对策研究进展与展望[J].中国沙漠,2005,25(3):306-314.
[11] 吕玲,谭敦炎.旱麦草属4种短命植物的结实特性及生殖包装[J].新疆农业大学学报,2005,28(3):21-25.
[12] 姚红,谭敦炎.胡卢巴属4种短命植物的生长特点及繁殖分配动态[J].新疆农业大学学报,2005,28(3):26-29.
[13] 班勇.植物生活史对策的进化[J].生态学杂志.1995,14(3):33-39.
[14] MacArthur R H. Some generalized theorems of natural selection[J]. Proc Natl Acad Sci USA,1962,48(1):1893-1897.
[15] Grime J P. Plant strategies and vegetation processes[M]. London: Wiley,1979:120-154.
[16] Kozlowski J, Uchmanski J. Optimal individual growth and reproduction in perennial species with indeterminate growth[J]. Evolutionary Ecology,1987,1:214-230.
[17] 包国章,康春莉,李向林.不同放牧强度对人工草地牧草生殖分配及种子重量的影响[J].生态学报,2002,22(8):1362-1366.
[18] 王仁忠,祖元刚.羊草种群生物量和能量生殖分配的研究[J].植物研究,2001,21(2):299-303.
[19] 王仁忠.放牧影响下羊草种群生殖生态学的研究[J].应用生态学报,2000,11(3):399-402.
猜你喜欢
小学生优秀作文(低年级)(2024年4期)2024-05-07 03:13:40
保健医苑(2022年4期)2022-05-05 06:11:28
语文周报·教研版(2022年2期)2022-04-27 00:58:07
小学生优秀作文(低年级)(2021年3期)2021-03-15 07:44:26
科学(2020年5期)2020-11-26 08:19:22
小学生优秀作文(低年级)(2020年9期)2020-10-26 09:10:34
语文世界(小学版)(2020年5期)2020-04-20 10:42:41
基层中医药(2018年4期)2018-08-29 01:26:00
中学历史教学(2016年4期)2016-11-11 07:11:35
长江师范学院学报(2016年1期)2016-08-23 11:40:08
