
典型性微量金属元素对藻华的激励趋势探讨
2010-11-20 08:12:33陈仕光王志红
华南师范大学学报(自然科学版) 2010年2期
陈仕光, 王志红, 曹 欣
(广东工业大学土木与交通工程学院,广东广州 510006)
典型性微量金属元素对藻华的激励趋势探讨
陈仕光, 王志红*, 曹 欣
(广东工业大学土木与交通工程学院,广东广州 510006)
Fe、Mn、Zn等典型性微量营养元素对藻类增殖的影响是探讨藻华成因分析的重要内容. 通过设计系列正交实验,研究代表性微量元素Fe、Mn、Zn对常见藻华生物(蓝藻、绿藻)的影响,结果表明:P、Fe、Mn、Zn的质量浓度变化对藻生长速率激励效果明显,显著性顺序依次为:Zn>P>Fe>Mn. 通过单独添加Fe、Mn、Zn的方法进一步进行了藻增长潜力实验. 结果表明:在Fe、Mn、Zn质量浓度分别为0.20、0.13、0.033 mg/L时,藻类生长速率达到最大值,低于该质量浓度时,金属离子质量浓度与藻类生长速率、生物量呈正相关;高于该质量浓度时,金属离子质量浓度与藻生长水平呈反相关;拟合Fe、Mn、Zn质量浓度与藻生长速率之间的数值关系,分别得到相关系数为0.93、0.87、0.90的回归方程;根据微量元素与藻类生长速率之间的相关性,可以对藻华的爆发时间或程度进行一定精度的预测. 正交实验中,锰的激励作用受微量元素之间的竞争性抑制而被消弱,反映出锰的作用机理比较复杂,有待于深入研究.
微量金属元素; 藻华; 生长速率; 生物量; 激励
有害藻华的发生对当地的经济、生态、人类身体健康造成严重危害,寻找有害藻华的诱发因素成为人们共同努力的目标. 长期以来,关于藻华(赤潮)成因的研究均侧重于N、P及N∶P比对藻类生长速率及最终生物量的影响评估方面,直到上世纪80年代,MARTIN等人在北太平洋发现在高营养低叶绿素(HNLC)的水体,浮游生物产量与铁(Fe)相关,以及后来在北太平洋、赤道地区、瑞典内陆湖泊等地区陆续发现铁对初级生产力有所限制后,人们越发清晰地认识到微量元素在藻华(赤潮)形成过程中的重要性[1-4].
最近20年来,人们在探寻微量元素与藻华因果关系方面取得了一系列的成果,对可能诱发藻类增殖的微量元素种类及每种元素的作用机理进行了细致的分析和深入的揭示,目前的研究成果主要体现在生物学的微观角度,已经发现:铁是叶绿素合成中某些酶或酶辅基的活化剂,补充铁可以提高藻类各种酶的活性;适量锰可加快藻类光合速率;Cu2+作为藻类呼吸和光合作用中多种酶的辅助因子,能提高酶的表达量;锌可促进各种酶的活性[5-13].
这些研究在水处理研究领域则意味着:金属离子可能在藻华爆发中发挥积极的作用. 但关于这些微量营养发挥作用的程度,与常规营养因素影响的比较,以及各种金属对藻类产生不同生物促进作用(促进或抑制)的浓度限值还很少报道,而这方面的研究对于深化认识藻华诱发机理,为藻华的防控提供科学依据至关重要.
Fe、Mn和Zn等3种元素因分布广泛及高度可生物利用,本研究中作为微量元素中的代表,结合常规营养磷(P),设计正交实验,比较P与Fe、Mn、Zn影响的显著关系,并通过单因素实验分析不同的Fe、Mn、Zn对不区分藻种的混合群体的作用效应,为富营养化水源藻华的预警和防控提供技术保障.
1 材料与方法
1.1试验材料
本研究的藻种取自于广州大学城广东工业大学人工湖(湖心N 23°02′,E113°23′). 该湖原水藻类密度在2×103~6×104个/L不等,种类主要为蓝藻和绿藻(藻种群中蓝藻、绿藻为优势藻种). 实验期间藻种分布情况及比例较稳定. 培养液采用朱氏改良10号[14]. 营养成分和项目分析所用试剂均为分析纯. 实验确定的因素有:P(Na3PO4·12H2O), Fe(FeCl3), Mn(MnSO4·H2O), Zn(ZnSO4·7H2O). Na3PO4采用蒸馏水配制,为避免金属离子发生沉淀反应,FeCl3、MnSO4、ZnSO4采用0.75 mmol/L EDTA溶液配制.
1.2实验方法
为了比较P、Fe、Mn、Zn对藻生长影响的相对大小,粗略判断各种元素对藻生长作用范围,先进行第1阶段的P、Fe、Mn、Zn正交试验,采用L16(45)正交表安排实验顺序. 实验水平的选取以各自元素在水体中的波动范围及藻类对金属离子的耐受度为依据[15-16]. 为了在有限的实验次数下,元素水平能覆盖金属元素对藻类的有效作用范围,第4水平均采用较高值. 因素水平如表1所示.

表1 因素水平表Tab.1 Factors and levels of orthogonal tests
第2阶段,Fe、Mn、Zn单因素实验,单因素实验的水平阈值根据正交实验中有效浓度水平来确定,以0.618法设计下一系列的因素水平[17],得到的实验顺序及各组水平如表2所示.
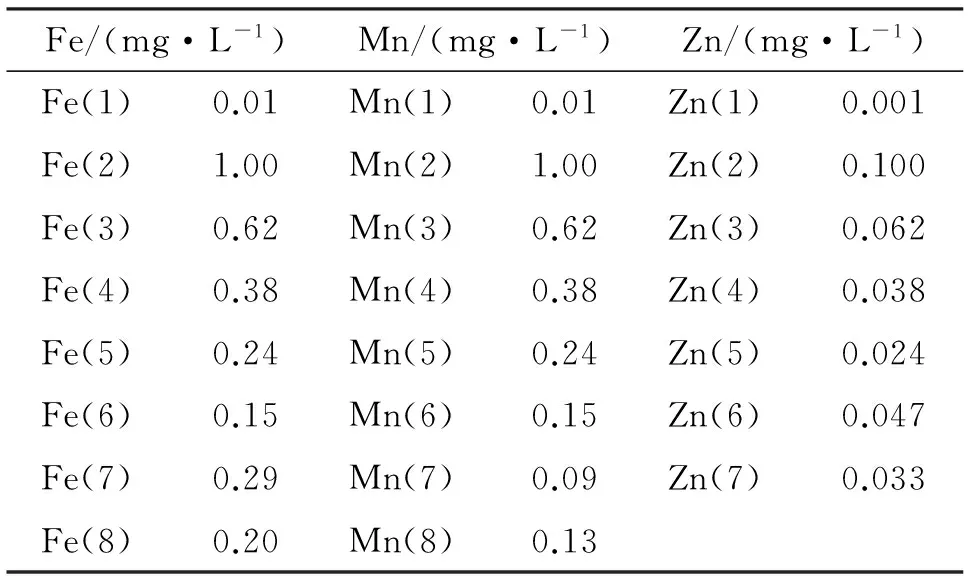
表2 单因子实验因素水平表Tab.2 Levels of single factor tests
试验前,先对原水藻种扩大培养1周,取一定体积的含藻原水用0.2 μm 微孔滤膜在负压下过滤,再以5 000 r/min的速度离心15 min,弃掉上清液,然后用0.1 mmol/L EDTA重悬以洗去细胞表面附带的微量金属离子,重复离心3次,使得待试验的藻细胞含微量金属不超过10-9mol/L. 经过上述程序后,将洁净藻细胞用无菌水稀释接种于不含P、Fe、Mn、Zn的培养基中进行饥饿培养,2 d以后加入P、Fe、Mn、Zn元素,开始实验周期,实验初始藻密度为1.3×104~3.4×104个/L. 当生物量连续3 d出现下降时,即认为已达到饱和值,随即停止实验.
试验采用自行设计的微循环藻类培养箱作为培养容器. 于室温条件静置培养,每天接受光照强度4 000~5 000 Lux,光暗比为14 ∶10,每日定时混匀3~4次,以促进营养物质循环,减少器壁效应,实验时间为2009年5月—9月,水温日变幅1.5~2.5 ℃,整个实验期间水温变化范围为25.6~31.5 ℃.
1.3测试方法
实验以藻细胞生长速率和峰值细胞产量为评价指标.
每天8:30定时取样,取样前先将培养箱水体混合均匀,准确吸取10 mL样品加入适量福尔马林固定藻细胞,然后用滴管吸取l滴含藻细胞的液体置于载玻片上,在10×20倍显微镜下,对藻细胞进行计数,计数2片取其平均值,绘制生长曲线,并计算藻峰值(个/L)及生长速率μ(d-1). 其中藻生长速率μ(d-1)的计算方法为:

(1)
式(1)中:X1为t1时的细胞数(个/L);X2为t2时的细胞数(个/L);t为培养时间(d).
总磷(TP):钼锑抗分光光度法;铁(Fe):邻菲罗琳分光光度法;锰(Mn):焦磷酸钾-乙酸钠分光光度法;锌(Zn):双硫腙分光光度法[18]. 使用的仪器有:重庆奥特光学仪器有限公司的BK系列生物显微镜;上海华岩仪器设备有限公司生产的HL-JS藻类计数框;北京瑞利分析仪器公司的VIS-723G可见分光光度计.
2 结果与分析
2.1一阶段正交实验结果分析
正交实验在于预估Fe、Mn和Zn对藻类生长产生不同激励作用(促进或抑制)的敏感浓度区间,为单因素实验水平设计提供参考. 同时也可以对这4种元素的影响显著程度进行比较. 由于磷是已证实的对藻类有显著影响的营养因子,实验以P作为对照物来评价3种微量元素对藻类影响的显著程度. 各组初始状态及藻生长效果见表3.
为能直接反应各质量浓度水平对藻生长速率的影响,通过式(1)计算得到表4. 表4中的平均生长速率是指每个因素的同一水平所在培养周期的生长速率均值.
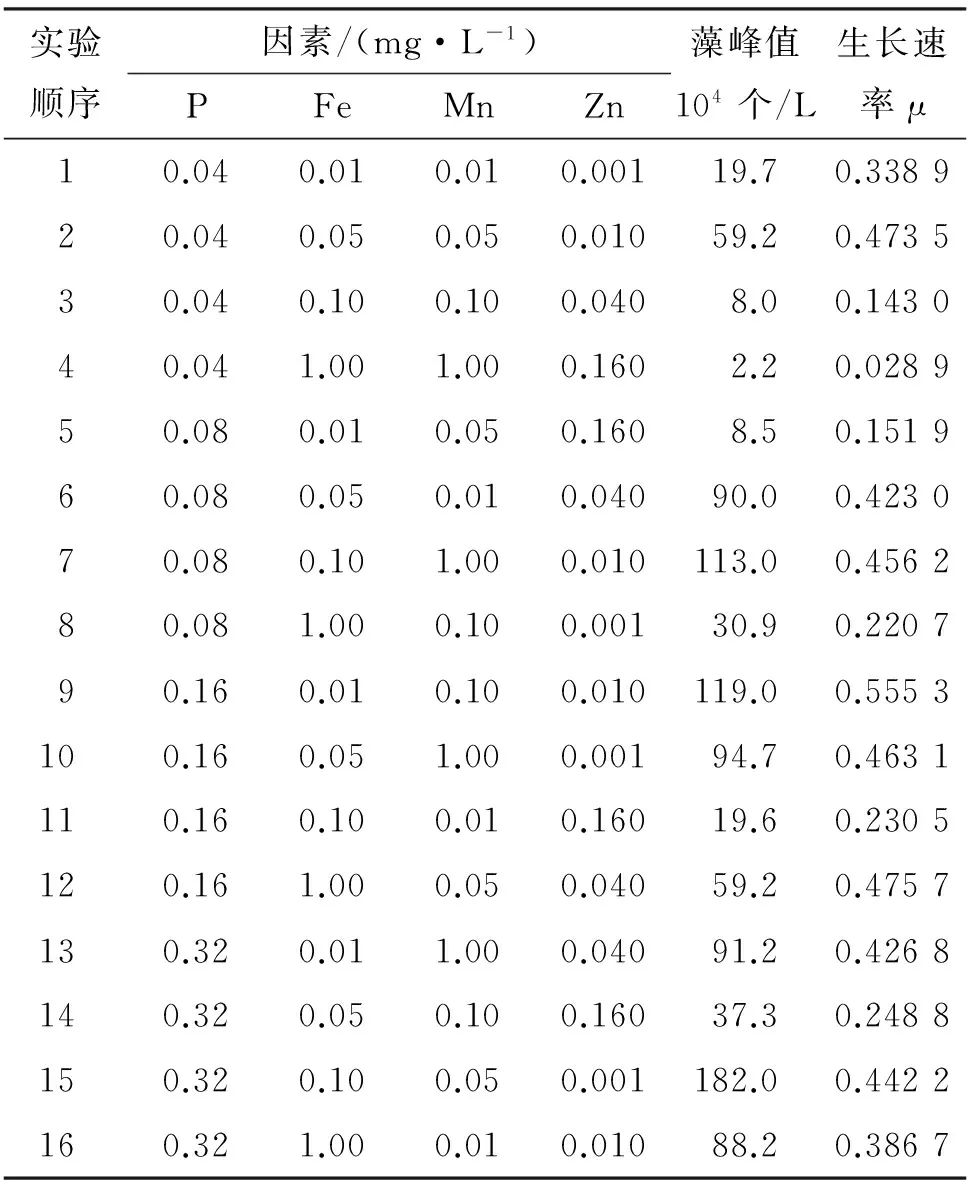
表3 正交实验结果及直观分析表Tab.3 Results of orthogonal tests and direct analysis
从表4可看出,Fe、Zn质量浓度分别对应为0.05、0.010 mg/L时,藻平均生长速率较0.01、0.001 mg/L条件下都有增加,表明增加的金属质量浓度有刺激藻生长的效果.然而,当Fe、Zn含量分别达到0.10 mg/L、0.040 mg/L时,藻生长速率出现显著下降,到1.00、0.160 mg/L时进一步降低;锰在0.01~0.05 mg/L区间内时,藻生长速率表现为增加趋势,在0.05~0.10 mg/L区间内时,表现为减少趋势,而在0.10~1.00 mg/L区间内时,藻生长速率又表现出了增加趋势. 依据各因素水平的生长速率极差判断:4种物质的显著性顺序依次是:Zn>P>Fe>Mn.
正交实验中,尽管可以反应出一定的趋势性关系,如在Fe:0.01~0.05 mg/L; Zn:0.001~0.010 mg/L区间变化时,表现为藻生长速率的增加趋势,在Fe:0.05~1.00 mg/L; Zn:0.010~0.160 mg/L区间变化时,表现为减少的趋势,但无法确切地了解铁、锰、锌质量浓度与藻生长速率之间的对应关系,藻类生长的最适质量浓度点也无法确定. 为了得出Fe、Mn、Zn质量浓度与藻生长态势之间的数值关系,采用水平梯度更精密的单因素实验,依次确定3种元素对藻类生长的激励规律.

表4 各因素各水平的平均生长速率Tab.4 Mean growth rate of each factor level
从表4看出,藻类在4个Fe、Mn、Zn质量浓度区间内,平均生长速率差异明显,即铁、锰、锌不同的质量浓度状态会对藻类产生不同的激励效果,表明正交实验中铁、锰、锌的4个水平皆处于对藻类产生激励作用的敏感浓度区间内. 根据正交实验结果,选定铁的质量浓度阈值为0.01~1.00 mg/L;锰为0.01~1.00 mg/L;锌为0.001~0.100 mg/L,见表2所示.
2.2二阶段单因素实验结果分析
2.2.1 Fe的激励规律 为了充分体现铁的生物活性,采用无铁朱氏培养液作为培养基质. 在添加不同Fe(避免发生价态转化和沉淀,采用有机络合形式Fe3+)条件下的藻生长曲线如图1所示,生长速率和最终藻高峰产量分别如图2、图3所示.

图1 不同Fe质量浓度下藻生长曲线Fig.1 Growth curve of algae under different concentration of Fe

图2 不同Fe质量浓度对生长速率的影响Fig.2 Influence of different concentration of Fe on growth rate
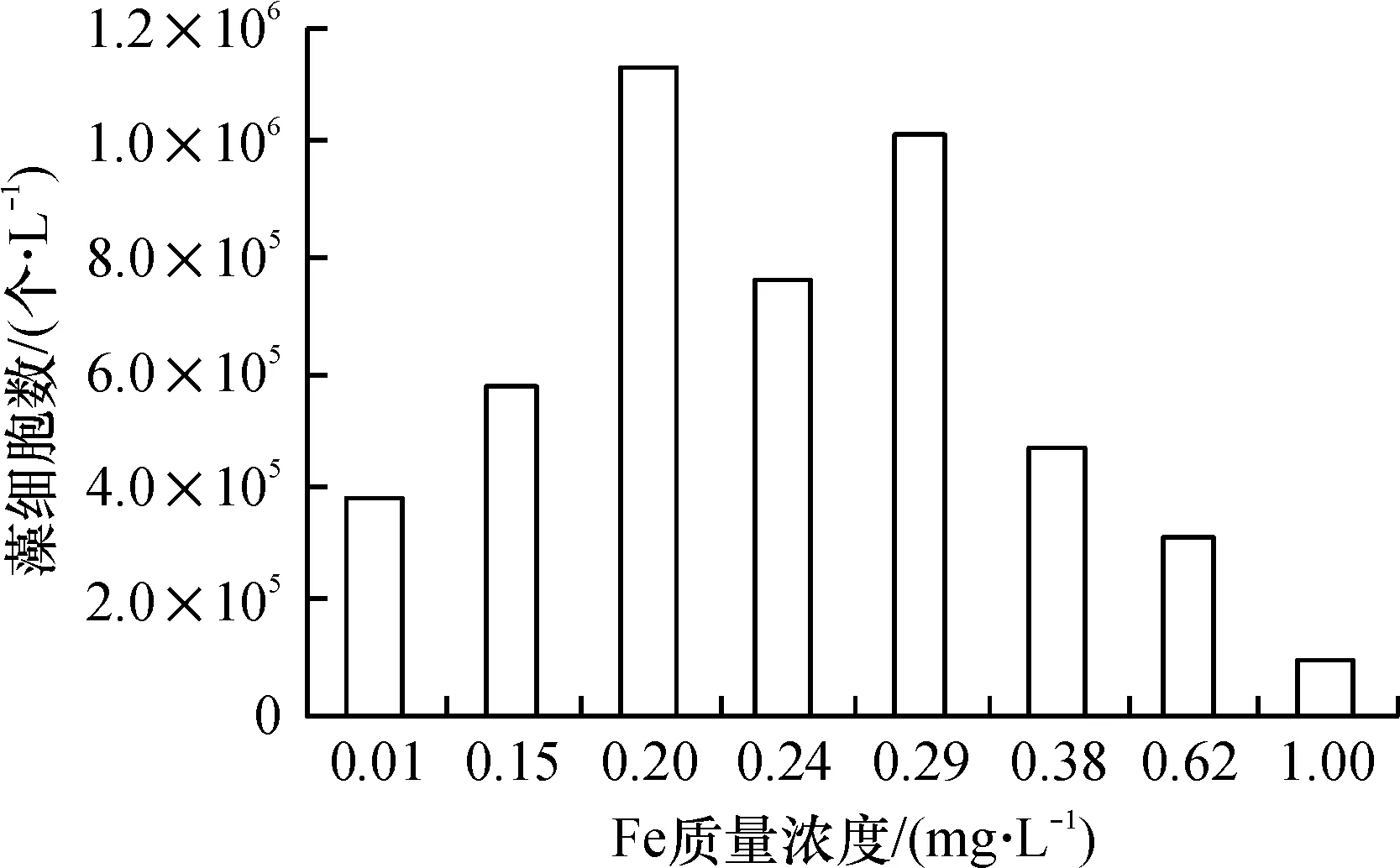
图3 不同Fe质量浓度对藻生物量高峰值的影响
Fig.3 Influence of different concentration of Fe on algae biomass maximum.
从实验结果来看,铁对藻类生长的影响分3种情况:
(1)缺铁条件:在0.01 mg/L时,初期细胞增长缓慢,第7 d以后,开始出现快速增长,到第10 d时细胞密度不再增加,最大生物量止于40×104个/L,为实验平均峰值的61.4%. 表明0.01 mg/L的铁含量尚不能满足藻类生长潜力的充分发挥. 原因在于,铁是藻生物体内光合反应和呼吸反应中不可或缺的电子载体,缺铁直接影响藻细胞的新陈代谢活动. 因此,藻类生长潜力的发挥必需足够量的可利用铁来保证.
(2)适量铁条件:从图1看出,在初始铁投加量达到0.15 mg/L时,藻细胞在第8 d出现了明显对数增长迹象,而0.20 mg/L和0.29 mg/L等2个系列从第4 d起就出现了对数增长趋势.这是由于,当有足够铁可供藻类利用时,藻类生命活动恢复正常,使藻类迅速进入旺盛的繁殖阶段. 随着铁质量浓度的提高,细胞峰值显著增加,在铁为0.15 mg/L时,藻生物量到第12 d达到最高峰,密度为50×104个/L左右,当铁上升至0.20 mg/L后,生物量高峰超过100×104个/L. 表明铁是藻类生物量的重要主导因素. 如图3所示,当铁质量浓度从0.01 mg/L提高到0.20 mg/L时,细胞密度增加近4倍.
(3)铁过量条件:从图2、图3看出,在初始投加的铁质量浓度高于0.38 mg/L后,生长速率和峰值细胞密度明显不如几个低铁系列. 尤其是0.62 mg/L和1.00 mg/L 2个组,在培养的初期,藻细胞密度不但没有增加,反而有下降趋势,在经历了5 d的调整以后虽然恢复了生长,但极其缓慢,表明藻类受到了铁的抑制作用. 这可以从藻生命活动的特点进行分析,藻细胞对金属离子的吸收缺乏主动选择性,当生长环境出现铁富余时,大量的铁离子聚集在藻细胞表面并源源不断地进入藻细胞内,超过了藻类的代谢能力,过量的铁离子与带负电荷的藻细胞壁发生电荷吸引,以及与一些含硫、氮、氧的官能团发生鳌和反应,造成对藻细胞的毒害作用[4],而藻类细胞受到的抑制作用与富余的铁含量呈正比.
本次实验各批次均在同温同光照条件下进行,同一对照组里使用的培养基质完全相同,因此,藻类生长的差异完全可以看做是由Fe质量浓度的变化引起的. 根据图3、图4的结果,Fe质量浓度按细胞生长密度排序依次是:0.20>0.29>0.24>0.15>0.38>0.01>0.62>1.0(mg/L),根据Fe质量浓度与藻类生长速率数值上的对应关系,采用最小二乘法拟合藻生长速率与铁质量浓度之间的函数关系,得到回归方程如下:
y=0.001x3-0.0234x2+0.1248x+0.1997
(R2=0.93),
(2)
式(2)中x为Fe3+质量浓度(mg/L),y为藻生长速率.
2.2.2 Mn的激励规律 试验所用的藻细胞经过了细致的洁净处理(反复离心,并用0.75 mmol/L EDTA溶液冲洗). 试验用的MnSO4采用0.75 mmol/L EDTA溶液配制,使细胞表面及培养基质中锰的含量不超过5.6×10-5mg/L. 因此,可认为初始添加的锰即是藻类唯一的外部锰源.
实验结果如图4、图5、图6所示.
结合图4、图5进行观察,藻类在锰质量浓度为0.13 mg/L的培养环境里生长最好,当初始锰含量在0.24 mg/L及以上时,生长速率与细胞数量均已开始下降. 到了0.62 mg/L条件时,图4显示,藻细胞直到第10 d才出现对数增长,而当锰为1.00 mg/L时,生长曲线接近一条水平直线,始终没有出现对数增长. 生物量与生长速率只有最高值的13.0%和47.4%.
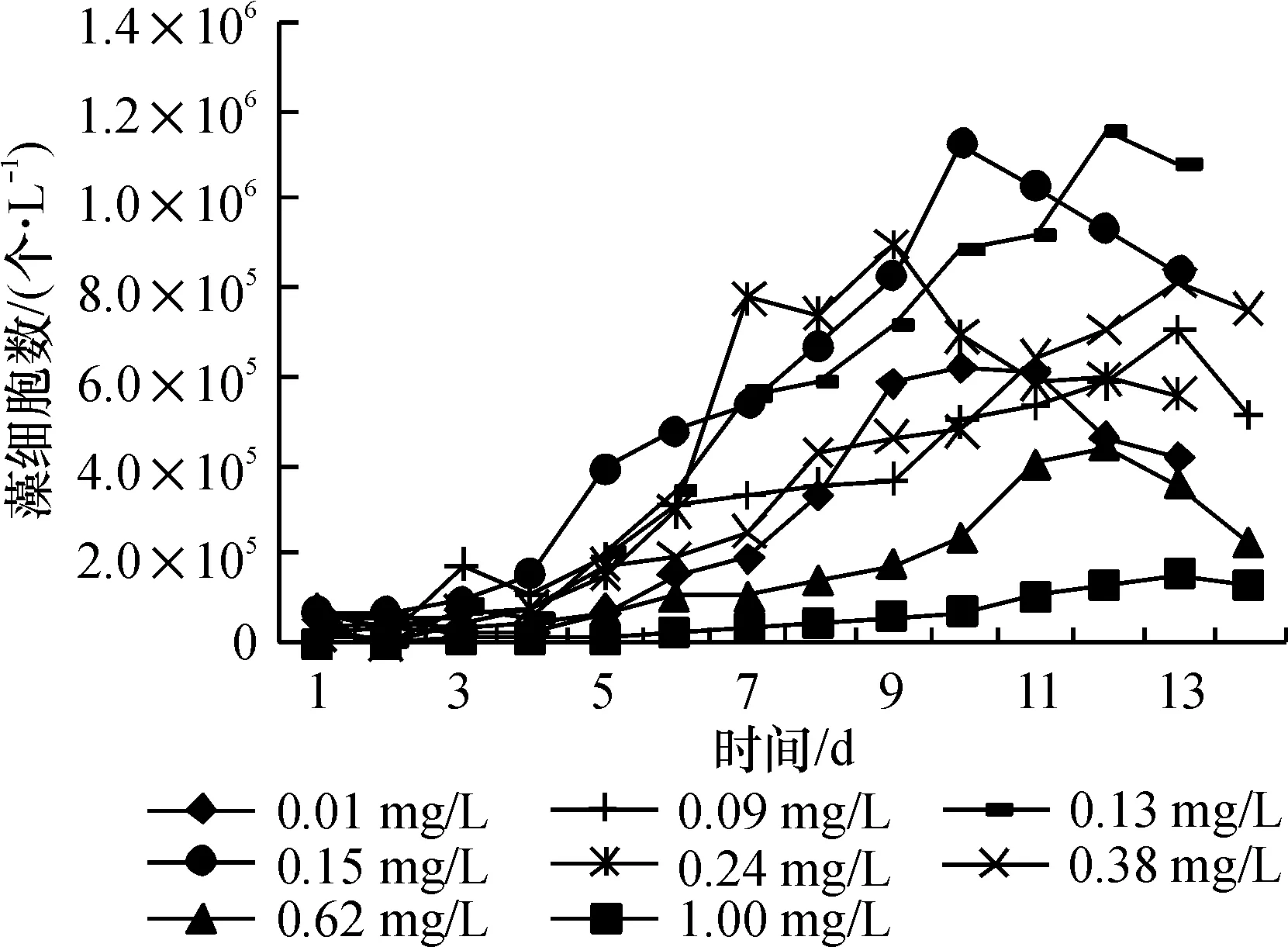
图4 不同Mn质量浓度下藻的生长曲线
Fig.4 Growth curve of algae under different concentration of Mn
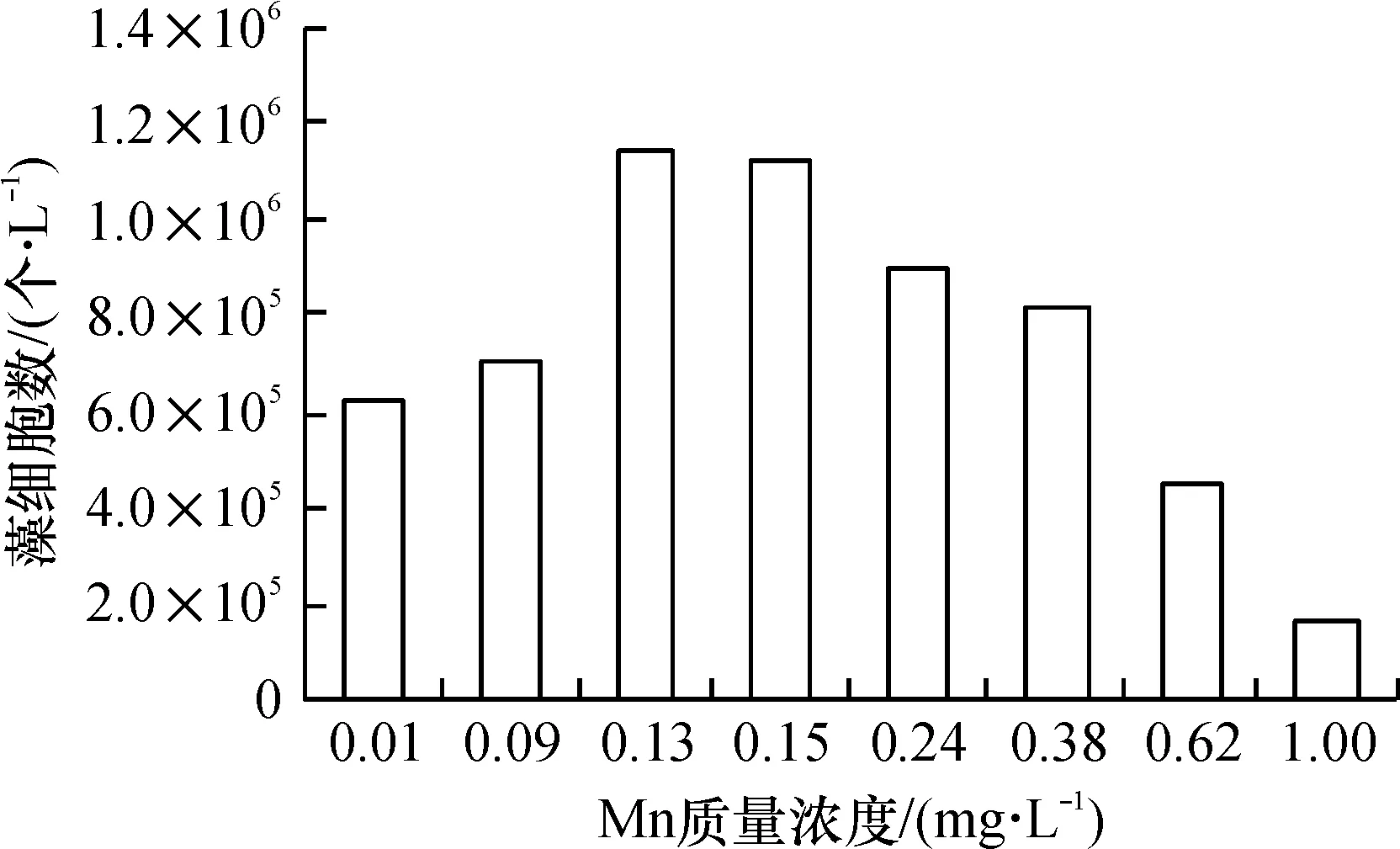
图5 不同Mn质量浓度对生长速率的影响
Fig.5 Influence of different concentration of Mn on growth rate
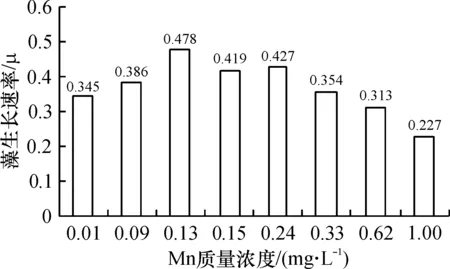
图6 不同Mn质量浓度对藻生物量的影响
Fig.6 Influence of different concentration of Mn on algae biomass
在0.13~0.24 mg/L之间时,属于藻生长的最佳质量浓度范围.
在0.24~1.00 mg/L之间时,锰对藻类的生长表现出明显抑制作用,藻生物量与质量浓度呈反相关关系. 认为抑制作用与藻生物体内的一种叶绿素降解酶有关,当锰离子的含量超过一定水平后,这种酶的活性就会超过叶绿素抗氧化酶的活性,原有的平衡被打破,造成叶绿素的含量下降[12].
实验结果表明:在控制营养、光照为最佳状态时,锰质量浓度的变化有可能使藻获得非常高的细胞比增长速率(0.478)和细胞密度(118×104个/L),产生严重藻华.
按生物量促进效果(细胞峰值)大小排序的锰质量浓度依次为0.13>0.15>0.24>0.38>0.09>0.01>0.62>1.0 (mg/L),同样采用最小二乘法拟合藻生长速率与Mn质量浓度关系,可得到方程式为:
y=-0.0138x2+0.1093x+0.2418(R2=0.87),
(3)
式(3)中x为Mn2+质量浓度(mg/L),y为藻生长速率.
根据本次实验结果,可以认为0.13~0.24 mg/L是藻类对锰需求的饱和质量浓度.
2.2.3 Zn的激励规律 藻类生物体中锌含量极少(每克藻干细胞含Zn 3.64~64.8 μg)[19],因而针对锌试验的精度要求也相应提高,本实验采用0.75 mmol/L EDTA配制溶解Zn2+溶液,尽量避免Zn2+的吸附效应,使得外加的Zn质量浓度变化梯度得以很好的体现. 实验结果如图7、图8、图9所示.
从图7看出,藻类生长在细微的Zn梯度差异条件下表现出了显著的生长差异. 整体来看,低Zn的3个组(0.033 mg/L及以下)藻类长势明显优于高Zn组(0.038 mg/L及以上). 以往关于Zn抑制浮游初级生产力的研究相对较少,一是难以检测到痕量Zn的变化,二是因为Zn的部分功能可以由其它元素如Co、Cu取代[20],因此,即使在缺Zn状态,藻类也不一定表现出生长受限. 本次实验,在前3个锌递增的系列中,藻类还是表现出明显的增加趋势(锌为0.033 mg/L时,藻类生长速率和生物量比在0.001 mg/L时分别增加了17%和15%),表明微量的Zn(0.001~0.033 mg/L)增加对藻生长会起到一定正向刺激作用.
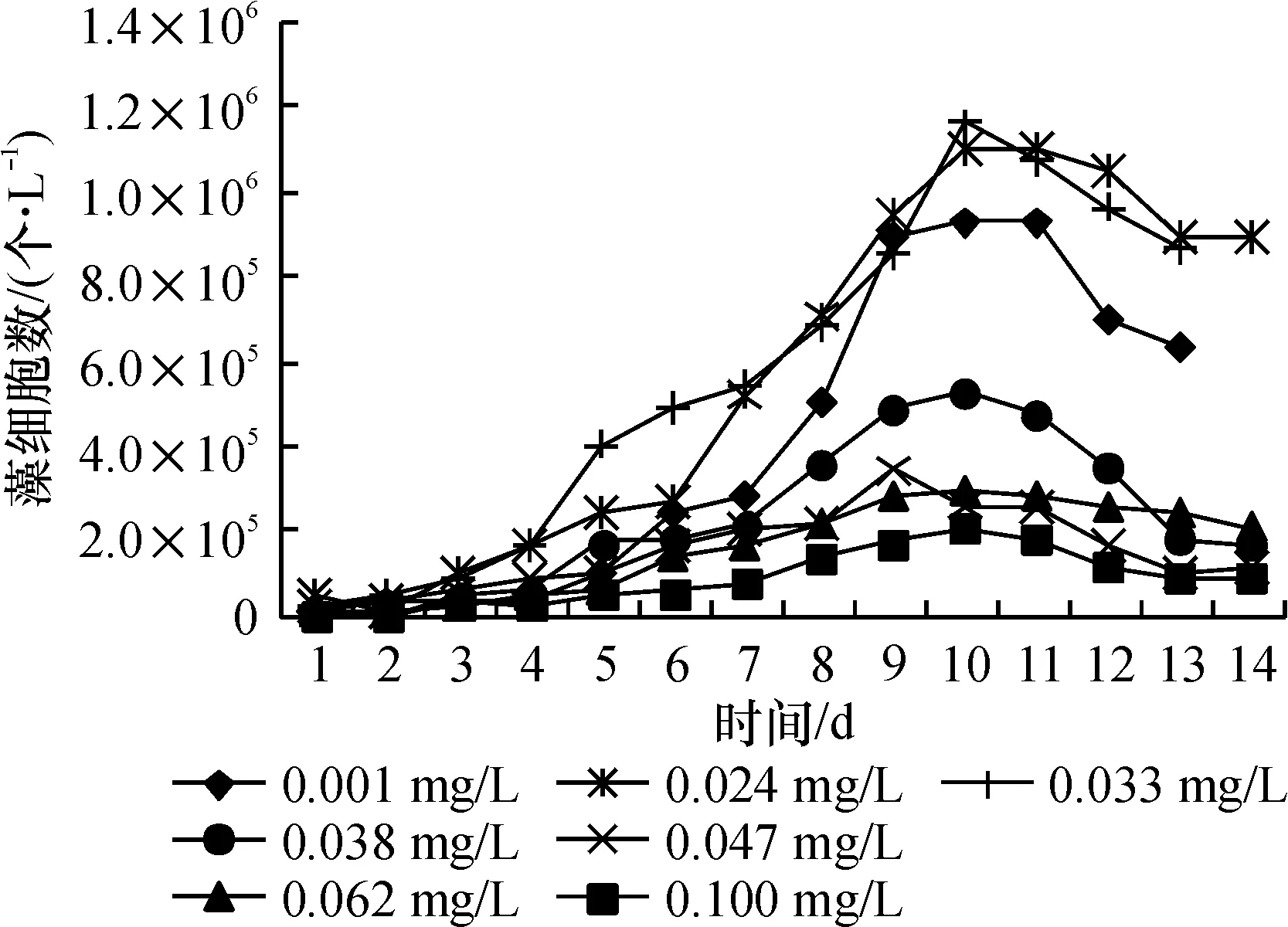
图7 不同Zn质量浓度下藻的生长曲线
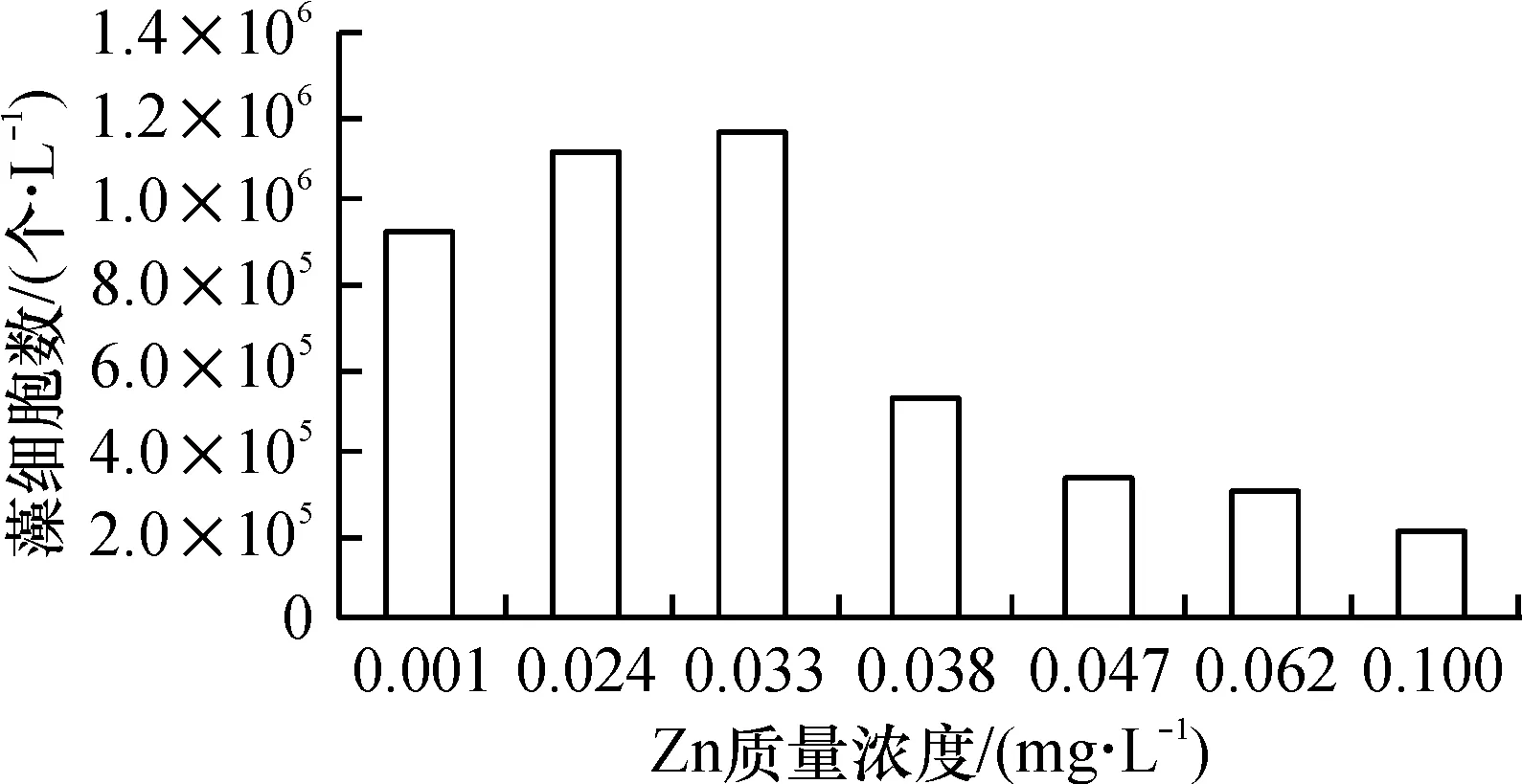
图8 不同Zn质量浓度对生长速率的影响
Fig.8 Influence of different concentration of Zn on growth rate
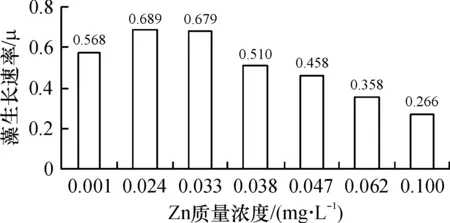
图9 不同Zn质量浓度对藻生物量的影响
Fig.9 Influence of different concentration of Zn on algae biomass
然而,当初始锌超过0.033 mg/L后,藻峰值生物量急剧下降(见图8),由此得出结论,对藻类造成抑制效应的锌质量浓度介于0.030~0.040 mg/L左右,超过此质量浓度后,藻类即处于低生长率状态.
使用重金属(包括Zn)抑制藻类,是目前普遍采用的方法,值得注意的是,不同的藻种群对Zn的耐受范围有异,但大多数藻类处于6.5×10-4mg/L到6.5×10-2mg/L以内[6]. 根据本次实验结果,对实验中的藻类群体,其最低负效应质量浓度出现在0.038 mg/L处,虽然有个别藻种在0.100 mg/L时还有生长,但大多数种类在锌高于0.047 mg/L后已死亡或进入长时间的迟滞期.
根据上述实验结果,锌元素的特点可总结为需求量少,但敏感度高,拟合方程为:
y=-0.0152x2+0.0581x+0.5766(R2=0.899),
(4)
式(4)中x为Zn2+质量浓度(mg/L),y为藻生长速率.
3 讨论
正交实验中,在元素质量浓度为:P(0.16 mg/L), Fe(0.05 mg/L), Mn(0.05 mg/L), Zn(0.010 mg/L)时,藻生长速率出现最大值,而在单因素实验中,在金属质量浓度分别为Fe:0.20~0.29 mg/L;Mn:0.13~0.24 mg/L;Zn:0.033~0.038 mg/L处藻生长最快. 相同质量浓度下,正交实验所反应出的金属元素激励趋势与单因素实验有些许差异,对此认为有以下几种可能:
首先是由于正交实验中,水平设计跨度大,无法反应出确切的最适质量浓度点;其次是正交实验时添加的金属元素种类较多,作用机理复杂,金属总量较单因素实验时要高,故藻类对单一金属所需的质量浓度值偏低;另外,各种元素之间的相互作用可能导致有些元素的作用效果减弱或增强,从而产生实验结果的差异. 如锰在单因素实验条件下对藻生长速率的影响极差为0.251,远比正交试验(0.094)显著,究其原因认为:与正交实验时各种金属元素之间的竞争性抑制作用有关,有研究表明,有一定量的Cu或Zn时,藻类对Mn的需求将减少,即藻类对金属离子具有选择性吸收特性,一种金属元素会替代藻类对另一种金属的需求[11,13]. 这是由于金属离子在与藻细胞分子结合时,具有相似功能的金属之间会发生相互取代,结合能力强的元素优先被利用[6]. 这也意味着,在寻找藻华的诱导因素时,不能只考虑与藻类生长直接相关的物质,还需考虑可能与这些物质发生相互关系的其它因子的变化情况.
二组实验的不同意义在于,正交实验反应了P、Fe、Mn、Zn各次影响的大小、主次关系,确定了金属元素对藻生长产生激励作用的敏感浓度区间,单因素实验分别确定了Fe、Mn、Zn质量浓度变化与藻生长速率之间的相关关系. 当水体只有一种元素变化时,可运用单因素实验结果对藻类的生长态势进行预测,当有多种元素变化时,可以运用正交实验的结果,对其影响大小进行比较,找出主要因素.
4 结论
(1)当Fe质量浓度低于0.20 mg/L时,藻生长速率、生物量与Fe质量浓度呈正相关,高于0.29 mg/L时,呈反相关;Mn含量低于0.13 mg/L时,Mn质量浓度与藻生长速率、生物量呈正相关,Mn高于0.24 mg/L时,呈反相关;在Zn不超过0.033 mg/L的条件下,增加Zn质量浓度促进藻类生长,超过0.038 mg/L后,抑制藻类生长. 最适藻类生长的Fe、Mn、Zn质量浓度分别为0.20、0.13、0.033 mg/L.
(2)对藻生长速率影响的显著性顺序依次是:Zn>P>Fe>Mn;
(3)用回归趋势来描述Fe、Mn、Zn对藻生长速率的激励函数,得到的方程分别为:
Fe:y=0.001x3-0.0234x2+0.1248x+0.1997
(R2=0.93);
Mn:y=-0.0138x2+0.1093x+0.2418(R2=0.87);
Zn:y=-0.0152x2+0.0581x+0.5766(R2=0.899).
(4)藻类最大生长速率在单因素状态下比在正交实验状态下需要的金属元素质量浓度要高,因此对藻类生长的预测需考虑在单一因素的变化和多种因素变化条件下的异同.
[1] MARTIN J H, FITZWATER S E, GORDON R M. Iron deficiency limits phytoplankton growth in Antarctic waters[J]. Global Biogeochemical Cycles,1990, 4: 52-55.
[2] TOBIAS V, LARS J T. Iron constraints on planktonic primary production in oligotrophic lakes[J].Ecosystems,2006 (9):1094-1105.
[3] SUN B K,YASUNORI T J, UNNO H J. Influences of iron and humic acid on the growth of the cyanobacterium anabaena circinalis biochemical[J]. Engineering Journal,2005,24:195-201.
[4] KATHLEEN S A, COLIN R A, JAMES W U. Nutrient additions generate prolific growth ofLyngbyamajuscule(cyanobacteria) in field and bioassay experiments[J]. Harmful Algae, 2007,6: 134-151.
[5] PATRICIA P R, PATRICIA E V,RICARDO B. Effect of copper on the photochemical efficiency, growth,and chlorophyll a biomass of natural phytoplankton assemblages Environmental[J]. Toxicology and Chemistry,2006,25(1):137-143.
[6] GAISINA L A, KHAIBULLINA L S. Influence of heavy metals on the morphology of the soil algaeXanthonemaexile(Klebs)Silva[J]. Eurasian Soil Science, 2007, 40(3):313-317.
[7] KANAKO N, MASAKAZU M, ICHIRO I. Ability of marine eukaryotic red tide microalgae to utilize insoluble iron[J]. Harmful Algae, 2005, 4: 1021-1032.
[8] KATHLEEN S A,COLIN R A,JAMES W U. In situ field experiment showsLyngbyamajuscula(cyanobacterium) growth stimulated by added iron, phosphorus and nitrogen[J].Harmful Algae,2008,7:389-404.
[9] LANCELOT C, HANNON E, BECQEVORT S, et al. Modeling phytoplankton blooms and carbon export production in the Southern Ocean: dominant controls by light and iron in the Atlantic sector in Austral spring[J]. Deep-Sea Research,Part I:Oceanographic Research Papers,2000,47(9):1621-1662.
[10] MAURICE L, MICHAEL G S, SONIA M. DMSP and DMS dynamics during a mesoscale iron fertilization experiment in the Northeast Pacific-Part I:Temporal and vertical distributions[J]. Deep-Sea Research, Part II:Topical Studies in Oceanography, 2006,53: 2353-2369.
[11] WILLIAM G S,SUSAN A H. Interactive effects of external manganese,the toxic metals copper and zinc, and light in controlling cellular manganese and growth in a coastal diatom[J]. Sundu and Huntsman,1998,43(7):1467-1475.
[12] KNAUER K, JABUSCH T, SIGG L. Manganese uptake and Mn(II) oxidation by the alga Scenedesmus subspicatus[J]. Aquat Sci,1999, 61: 44-58.
[13] JEFFREY M R, MILYON R S.Effect of manganese and nickel on growth of selected algae in pH buffered medium[J]. Wat Res,1999,33(10): 2448-2454.
[14] 沈韫芬,章宗涉,龚循矩. 微型生物监测新技术[M].北京:中国建筑工业出版社,1990:146-148.
[15] 雷衍之. 养殖水环境化学[M]. 北京: 中国农业出版社,2008:140-156.
[16] 张铁明, 杜桂森, 杨忠山,等. 锌对2 种淡水浮游藻类增殖的影响[J]. 西北植物学报,2006,26(8):1722-1726.
ZHANG Tieming, DU Guisen,YANG Zhongshan,et al. Effects of zinc on two phytoplanktons in fresh water[J]. Journal of northwestern plants, 2006,26(8):1722-1726.
[17] 吴俊奇,李燕城. 水处理实验技术[M]. 北京:中国建筑工业出版社,2009:6-13.
[18] 国家环境保护总局. 水和废水监测分析方法[M]. 4版. 北京:中国环境科学出版社,2002:101-385.
[19] SHI Y J, HU H H, MA R Y, et al.Improved use of organic phosphate by skeletonema costatum through regulation of Zn2+concentrations[J]. Biotechnology Letters,2004,26:747-751.
[20] MIAO A J, WANG W X, PHILIPPE J.Comparison of Cd,Cu,and Zn toxic effects on four marinephytoplankton by pulse-amplitude-modulated fluorometry[J]. Environmental Toxicology and Chemistry,2008,24(10):2603-2611.
Keywords: trace metal elements; algae bloom; growth rate; biomass; motivation
【责任编辑 成 文】
RESEARCHONTHEMOTIVATIONTRENDOFTYPICALTRACEELEMENTSONALGAEBLOOM
CHEN Shiguang, WANG Zhihong*, CAO Xin
(Faculty of Civil and Transportation Engineering, Guangdong University of Technology, Guangzhou 510006,China)
The effect of micro-nutrients (Fe, Mn, Zn) on the proliferation of planktonic algae colony is one of the most important contents in the research of cause analysis of algae blooms. Experiments on the impacts of typical micro-nutrients (P, Fe, Mn, Zn) on common algal blooms organisms (cyanobacteria, chlorophyta) were carried out by a series of orthogonal experiments. The results show that these nutrients have significant impacts on algae growth and stimulated cell-specific growth rates with Zinc>Phosphorus>Iron>Manganese. AGP (algae growth potential) experiments by separate addition of iron, manganese and zinc were further performed through single-factor experiment. The data obtained show that maximum algae growth rate appeared when concentration of Fe,Mn,Zn were 0.20 mg/L, 0.13 mg/L, 0.033 mg/L,respectively. Furthermore, a positive motivation to the growth rate and biomass of algae was emerged with the concentrations of Fe, Mn and Zn range from 0.01~0.20, 0.01~0.13, 0.001~0.033 mg/L, while there is a negative motivation with the increase of the concentrations of Fe, Mn and Zn above 0.20, 0.13, 0.033 mg/L, respectively. By fitting the numerical relationships between concentrations of Fe, Mn, Zn and algal cell-specific growth rate, regression equations with correlation coefficient of 0.93, 0.87, 0.90 were obtained respectively. The results make it be able to predict the time and severity of outbreak of algae blooming to a certain precision extent according to the correlation between trace elements and algae growth rate or biomass. Experiment consequence also suggested that the significant levels of Manganese to algae had been severely weakened by the inhibitory effect among multi-microelements during orthogonal experiments. These results reveal that the mechanism of Manganese is rather complicated that further researches are needed to be performed.
2010-01-11
广东省科技攻关资助项目(2007B031700007);广东省自然科学基金资助项目(9151009001000048)
陈仕光(1982—),男,湖南邵阳人,广东工业大学2007级硕士研究生, Email:luyi813929@163.com;王志红(1975—),女,江西赣州人,博士,广东工业大学教授,主要研究方向:微污染水源预警及处理技术,Email:gdwzhihong@126.com.
* 通讯作者
1000-5463(2010)02-0082-08
TU991
A
猜你喜欢
当代水产(2021年8期)2021-11-04 08:49:00
今日农业(2020年19期)2020-12-14 14:16:52
军事文摘(2020年20期)2020-11-16 00:31:40
中学生数理化·高一版(2020年6期)2020-07-25 01:47:24
今日农业(2019年10期)2019-01-04 04:28:15
中学生数理化·八年级物理人教版(2017年3期)2017-11-09 03:05:33
环境科技(2016年2期)2016-11-08 12:18:22
中国科技信息(2015年2期)2015-11-16 08:18:32
上海金属(2014年5期)2014-12-20 07:58:39
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
