
两种体色三疣梭子蟹线粒体DNA部分片段序列的比较分析
2010-09-24 08:09:30迟大利沈颂东阎斌伦
海洋科学 2010年11期
迟大利, 高 焕 沈颂东, 阎斌伦
(1. 淮海工学院江苏省海洋生物技术重点建设实验室, 江苏 连云港 222005; 2. 苏州大学生命科学院, 江苏苏州 215123)
两种体色三疣梭子蟹线粒体DNA部分片段序列的比较分析
迟大利1,2, 高 焕1, 沈颂东2, 阎斌伦1
(1. 淮海工学院江苏省海洋生物技术重点建设实验室, 江苏 连云港 222005; 2. 苏州大学生命科学院, 江苏苏州 215123)
三疣梭子蟹(Portunus trituberculatus)作为重要的海洋经济动物, 其常见体色(头胸甲)为茶绿色。近年来在我国沿海海域海捕三疣梭子蟹中开始出现体色为紫色的一类梭子蟹, 除体色不同外, 二者表型特征并无显著差异。对两种体色三疣梭子蟹不同个体的线粒体COI和16S rRNA基因进行了序列比对分析以判定二者之间的亲缘关系。结果发现COI和16S rRNA基因在两种体色三疣梭子蟹群体中的核苷酸序列一致性分别为99.87%和99.88%, 表明紫色三疣梭子蟹并未发生亚种的分化, 即紫色和茶绿色群体属于同一个种。并利用GenBank数据库检索了梭子蟹科其他15种海产蟹的16S rRNA序列, 进行了序列同源性比对分析和分子系统学方面的探讨, 为梭子蟹种属分类学研究提供分子依据。
三疣梭子蟹; 体色; 线粒体DNA; 16S rRNA; COI
三疣梭子蟹(Portunus trituberculatus)隶属于甲壳纲(Crustacea), 十足目(Decapoda), 梭子蟹科(Portunidae), 梭子蟹属(Portunus), 又称梭子蟹、枪蟹、海螃蟹、海蟹、门蟹等, 是一种大型海洋经济动物, 广泛分布于中国、日本、朝鲜等西太平洋沿岸海域[1], 具有肉质好、食物链短、生长快、产量高等优点[2], 是我国重要的海洋渔业资源。鉴于其较高的经济价值及其在生态环境中的重要作用, 自20世纪50年代起, 国内外学者相继对三疣梭子蟹生活习性、生理生化、胚胎发育、养殖模式、群体遗传结构、超微结构观察和核型分析等方面做了深入的研究, 而对三疣梭子蟹分子生物学展开的研究工作尚处于起步阶段, 且主要集中在分子标记研究领域[3]。
常见三疣梭子蟹体色(头胸甲)为茶绿色, 近年来, 在我国从渤海到东海广大海域的海捕野生群体中发现一类体色为紫色的梭子蟹。从形态上观察, 除了身体颜色存在差异外, 两种体色梭子蟹其他表型特征相似。对于此类紫色梭子蟹的来源有不同的说法: 有的认为雌性梭子蟹为紫色, 雄性蟹为茶绿色[4];有的认为它们属于两个不同的种[5]。因此, 有必要了解不同体色梭子蟹之间的关系及其与其他蟹类的系统发生学关系。
由于母性遗传、进化率低以及缺乏分子间遗传重组等特点, 线粒体DNA已经广泛应用于群体遗传结构分析和不同分类水平物种间亲缘关系的研究[6]。迄今为止, 线粒体两个保守序列基因COI(细胞色素c氧化酶亚基I基因)和16S rRNA基因已经成功应用于甲壳类动物的生物学亲缘关系鉴定[7]。通过应用长PCR技术, Yamauchi等[7]首次对三疣梭子蟹线粒体基因组进行全序列分析研究。郭天慧等[2]对三疣梭子蟹线粒体DNA COI和16S rRNA基因片段序列进行比较研究发现, 16S rRNA基因较 COI基因更为保守,表明 COI基因适于种内遗传多态性研究, 而 16S rRNA基因适于种属间遗传多态性分析。
Place等[8]对可口美青蟹(Callinectes sapidus)整个线粒体基因组进行了测序和基因定位, 发现可口美青蟹和三疣梭子蟹线粒体基因组长度(分别为16 263 bp 和16 026 bp)十分接近, 且二者线粒体基因组内基因序列及相关功能基因的定位均十分相似,表明三疣梭子蟹和可口美青蟹的亲缘关系相近, 但是, 三疣梭子蟹和可口美青蟹分别生活在太平洋和大西洋沿岸[8], 处于地理隔离状态, 分类学上分别隶属于梭子蟹属和青蟹属(Callinectes), 且二者体色亦有不同, 因此有必要了解二者在分子系统分类学中的关系。
本文期望通过线粒体DNA COI和16S rRNA部分基因片段在茶绿色和紫色梭子蟹间的序列比对分析, 以阐释清楚两种体色梭子蟹的亲缘关系, 即二者是否达到亚种的分化水平; 并对三疣梭子蟹与梭子蟹属其他 14种梭子蟹以及青蟹属可口美青蟹的16S rRNA基因进行序列同源性比对分析, 以探讨三疣梭子蟹与其他海产蟹类的分子系统发育关系。
1 材料与方法
1.1 材料
实验所用紫色和茶绿色三疣梭子蟹购自连云港水产品市场, 时间为2007年10月至2008年3月。
1.2 样品DNA的提取
取三疣梭子蟹附肢肌肉, 使用上海赛百盛基因技术有限公司生产的树脂型TM基因组 DNA提取试剂盒提取基因组DNA, 经Gene QuantPro核酸定量仪(通用公司, 美国)测定基因组DNA浓度, -20 ℃保存备用。
1.3 PCR扩增及目的基因序列测定分析
三疣梭子蟹线粒体DNA COI和16S rRNA基因片段扩增所用特异性引物由上海生工生物工程技术服务有限公司合成。引物序列参见郭天慧等[2]的文献,分别为 COI-H/COI-L(正向: 5’-taaacttcagggtgaccaaaaaatca-3’, 反 向 : 5’-ggtcaacaaatcataaagatattgg-3’)和16S-AR/16S-BR(正向: 5’-cgcctgtttatcaaaaacat-3’, 反向: 5’-ccggtctgaactcagatcacgt-3’)。
PCR 反应体系(25.0 μL)为: ddH2O 12.3 μL, 10×PCR 缓冲液 2.5 μL, 25 mmol/L MgCl22.0 μL, 10 μmol/L dNTPs 2.0 μL, 5 U/μLTaqDNA 聚合酶 0.2 μL,10 μmol/L 上下游引物各 2.0 μL, 500 μg/L DNA 模板2.0 μL。PCR扩增条件为: 94 ℃预变性5 min; 94 ℃变性45 s, 48 ℃退火1 min, 72 ℃延伸1 min, 循环30次; 最后72 ℃延伸5 min。PCR试剂均购自上海赛百盛基因技术有限公司。
PCR扩增所得目的DNA片段直接送至上海生工生物工程技术服务有限公司进行序列测定, 再通过DNAMAN(Version 6.0.40)软件进行紫色和茶绿色梭子蟹线粒体DNA COI和16S rRNA基因片段序列比对分析。
1.4 16种海产蟹16S rRNA基因片段的序列分析及分子系统学探讨
根据三疣梭子蟹的种属特异性及序列分析结果,在GenBank数据库中有选择性地检索出同属于梭子蟹科(Portunidae)的、梭子蟹属其他 14种梭子蟹(P.anceps;P. bahamensis;P. binoculus;P. depressifrons;P.floridanus;P. gibbesii;P. ordwayi;P. rufiremus;P. sayi;P. sebae;P. spinicarpus;P. spinimanus;P. ventralis;P.vossi)及青蟹属可口美青蟹(C. sapidus)的 16S rRNA基因片段, 共15种海产蟹类型。利用DNAMAN软件对三疣梭子蟹与这15种海产蟹的线粒体DNA 16S rRNA基因片段核苷酸序列进行结构分析, 并以可口美青蟹作为外类群进行分子系统分析。邻接法(Neighbor-joining, NJ)分析使用Kimura 2-paramater,系统树分支的置信度采用重复抽样分析(Booststrap analysis)的方法, 重复抽样的次数为1000次。
2 结果
2.1 线粒体DNA COI和16S rRNA基因片段PCR扩增结果
本实验采用三疣梭子蟹基因组DNA为模板对其线粒体 COI和 16S rRNA基因部分片段分别进行PCR扩增及电泳检测。结果表明, 两个基因片段的PCR扩增产物均产生了清晰可辨的扩增条带, 且二者扩增条带分子量大小与参考文献[2]相一致。
2.2 线粒体DNA COI和16S rRNA基因的序列分析
从COI和16S rRNA基因PCR扩增样品中分别随机选取4个紫色和4个茶绿色个体的扩增样品进行测序分析(图1和图2)。经测定分析后得到了长度为658 bp的COI基因序列和长度为524 bp的16S rRNA基因序列。相关序列分析结果为: 紫色和茶绿色梭子蟹个体中COI基因(G+C)摩尔分数(DNA碱基比)分别为46.8%和46.7%; 两类个体中16S rRNA基因(G+C)摩尔分数均为53.15%; 经DNAMAN软件分析合并的COI和16S rRNA基因中(A+T)含量分别为63%(A=36.6%; C=16.4%; G=20.5%; T=26.4%)和69.7%(A=34.5%; C=12.4%; G=18.0%; T=35.2%), 这与三疣梭子蟹整个线粒体DNA序列中70.2%的(A+T)含量相近[7], 而且, COI和16S rRNA基因在紫色和茶绿色群体间的核苷酸序列一致性(sequence identity)分别达到了99.87%和99.88%。
2.3 三疣梭子蟹 16S rRNA基因与其他海产蟹的同源性比较
经 GenBank数据库选择性地检索, 得到用于同源性比对分析的梭子蟹科共15种海产蟹16S rRNA基因片段, 相应分类和GenBank登录号见表1。

图1 紫色(P)和茶绿色(G)梭子蟹个体COI基因片段的序列比对Fig. 1 Multiple sequence alignment of COI gene fragments of tea-green (G) and purple (P) crab samples
由于选取的 16种海产蟹属于同一科(梭子蟹科)内不同属间的类型, 其中 15种为同一属(梭子蟹属)内不同种间的类别, 本文通过常用于系统发生比较的线粒体DNA 16S rRNA基因对16种海产蟹的核苷酸序列同源性进行分析比较。DNAMAN软件进行序列同源性比对分析所得结果见表2。

图2 紫色(P)和茶绿色(G)梭子蟹个体16S rRNA基因片段的序列比对Fig. 2 Multiple sequence alignment of 16S rRNA gene fragments of tea-green (G) and purple (P) crab samples

表1 用于16S rRNA基因比对分析的梭子蟹科15种海产蟹的分类学位置及GenBank登录号Tab. 1 Classification of 15 species of Portunidae and the GenBank accession numbers for 16S rRNA analysis
16种海产蟹之间的16S rRNA基因核苷酸序列一致性(sequence identity)为 88.06%, 序列同源性(sequence homology)范围为 76.2%~100%。梭子蟹P.anceps和P. rufiremus之间的16S rRNA基因核苷酸序列同源性最低, 为 76.2%; 梭子蟹P. bahamensis和P.depressifrons以及P. spinimanus和P. vossi之间的16S rRNA基因核苷酸序列同源性最高, 均达到100%。与其他15种海产蟹16S rRNA基因同源性比对分析结果表明, 三疣梭子蟹和其他不同种属海产蟹间的核苷酸序列的同源性为81.7%~91.7%, 其中, 与赛氏梭子蟹(P.sayi)同源性最高, 达到91.7%; 与P. rufiremus的同源性最低, 为81.7%; 而与可口美青蟹(C. sapidus)的同源性仅次于赛氏梭子蟹, 达到87.7%。

表2 16种海产蟹线粒体DNA 16S rRNA基因核苷酸部分序列同源性比较Tab. 2 Comparison of sequence homology (%) among 16S rRNA genes from 16 species of Portunidae
2.4 16S rRNA基因在系统学分析中的应用
基于线粒体 DNA 16S rRNA基因核苷酸序列,以可口美青蟹为外群对16种海产蟹进行系统发育分析。通过应用邻接法(NJ法)构建的的分子系统发育关系树(图 3), 枝长表示分歧度, 枝上的数值是 1000次重复抽样检验的置信度值。聚类分析结果表明, 三疣梭子蟹首先与赛氏梭子蟹(P. sayi)聚为一类, 再与可口美青蟹聚在一起。另外, 梭子蟹P. bahamensis和P. depressifrons以及P. spinimanus和P. vossi之间的 16S rRNA基因核苷酸序列的高同源性在分子系统树的聚类结果中也反映了这一点。由于针对梭子蟹种属内的研究主要集中在三疣梭子蟹等重要经济种类, 且天然地理分布阻隔也造成对梭子蟹属内各种之间的分类学研究较为匮乏, 该聚类分析结果可为梭子蟹种属分类学研究提供分子依据。
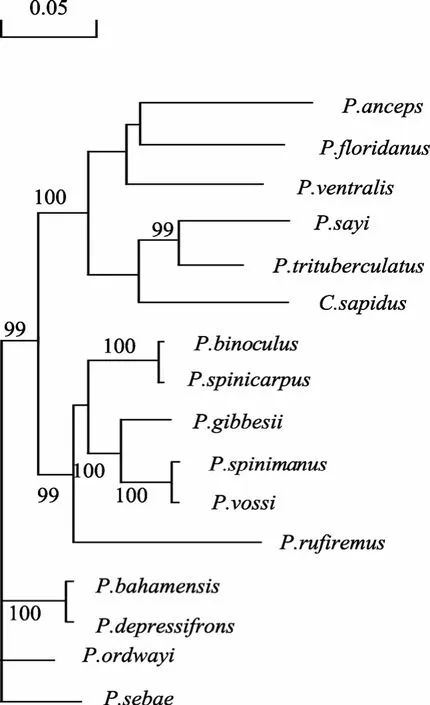
图3 基于16种海产蟹线粒体DNA 16S rRNA基因核苷酸序列通过邻接法构建的分子系统树(NJ树)Fig. 3 Molecular phylogenetic tree on the nucleotide sequence of 16S rRNA gene from 16 species of Portuni dae constructed by Neighbour-Joining method (NJ tree)
3 讨论
3.1 线粒体保守序列基因分析在系统分类学中的应用探讨
当前, 动物分子分类学研究成果多建立在线粒体DNA相关基因序列分析的基础之上。线粒体中核糖体 RNA基因(rRNAs)和细胞色素 c氧化酶亚基 I基因(COI)的核苷酸序列比对, 是分子系统亲缘分析行之有效的方法[9]。陈丽梅等[10]对 3种蛏类 16S rRNA和 COI基因片段序列进行了比较并对其系统学进行了初步研究, 发现COI基因的进化速率与16S rRNA基因相比要快, 16S rRNA基因则相对保守一些。徐琰等[11]通过运用16S rRNA基因序列初步探讨了部分真虾类的系统发育关系, 结果表明, 16S rRNA基因片段的序列很适合研究真虾类属间系统发育关系, 而在研究属以上高级阶元或种以下阶元间的系统发生关系时较不敏感, 因其变异对于种级水平显得过于保守, 而对于科级以上阶元又显得过快。蒋钦杨等[12]在 3个不同群体罗氏沼虾(Macrobrachium rosenbergii)线粒体16S rRNA基因序列分析及遗传差异比较中也指出, 线粒体16S rRNA基因是动物种内个体间较为保守的序列, 主要用于种以上水平的变异分析。在细菌分类学鉴定方面,16S rRNA基因序列分析是当前细菌分类中最精确的一种技术[13]。刘佳文等[14]指出通过分子生物学技术分析16S rRNA基因, 可将分枝杆菌(Mycobacterium)鉴定到种的水平。由于具有较强的中间解析和鉴别能力, 线粒体16S rRNA基因正广泛应用于脊椎动物和无脊椎动物的系统与进化学及种类鉴别中[15]。
目的基因序列测定及比对分析可以直观反映两个群体之间的亲缘关系。以细菌鉴定为例, 当两DNA间同源性为 100%, 即可认为是源于同一个体;同源性为80%~90%则被认源于为同一种内或同一亚种; 同源性为 60%~70%被认为源于同一种内的不同亚种, 同源性为 20%~60%则被认为是源于同一属的不同种(http://space.biox.cn/3776/viewspace-4586.html)。对不同个体或群体间DNA碱基比(G+C)摩尔分数的差异比较的分析方法已经广泛应用于微生物的分类学的亲缘关系鉴定的研究中。一般认为, (G+C) 摩尔分数相差超过5%就不可能属于同一个种, 相差超过10%就可考虑是不同属[16]。本实验选取的线粒体DNA两个保守基因的序列比对分析结果显示了COI和16S rRNA在两个群体中均接近100%的核苷酸序列一致性, 且紫色和茶绿色梭子蟹群体中 COI基因(G+C)摩尔分数差别为 0.1%; 16S rRNA基因(G+C)摩尔分数则完全相同, 说明紫色和茶绿色梭子蟹群体间亲缘关系十分相近, 二者同属于一个种, 紫色三疣梭子蟹并未发生亚种分化。
在 COI基因序列比对分析中, 紫色梭子蟹个体间仅发现单碱基差异, 序列歧异为转换(G-A); 而茶绿色梭子蟹不同个体间有较多的(6个)碱基突变, 序列歧异基本上是转换(T-C, G-A)多于颠换(G-C), 且其中有一处单碱基序列差异(G-A)与紫色梭子蟹个体相同, 这说明紫色梭子蟹群体的线粒体DNA序列在进化关系上较为保守(图1); 在16S rRNA基因序列比对分析中, 紫色和茶绿色梭子蟹群体内不同个体间均只出现单碱基突变, 而且仅为单碱基 T的重复序列数目不同(图2), 反映了16S rRNA基因高度保守的特性, 说明16S rRNA基因更适用于亲缘关系较远物种之间的分类学研究鉴定。
在利用线粒体DNA 16S rRNA基因对16种海产蟹的核苷酸序列同源性进行分析中, 三疣梭子蟹与可口美青蟹的同源性仅次于赛氏梭子蟹(91.7%), 达到87.7%, 且DNAMAN软件分析表明三疣梭子蟹与可口美青蟹间的 16S rRNA基因核苷酸序列一致性达到 83.00%, 表明二者相近的亲缘关系。以上结果表明, 海产蟹线粒体DNA 16S rRNA基因是相对保守的, 适合用于梭子蟹科内不同种间同源性比较及遗传多样性的分析。
鉴于本实验所用线粒体DNA COI和16S rRNA基因序列均未检测到可用于鉴定两种体色三疣梭子蟹间的差异, 可进一步对线粒体DNA中核苷酸序列变化较大的区域(如, 线粒体 DNA 控制区[17,18]等)进行序列比较分析, 以得到基于碱基序列差异的不同体色三疣梭子蟹间的具体亲缘关系。
3.2 三疣梭子蟹体色成因分析
体色在海洋蟹类的生活史中扮演重要的角色。首先它可以作为一种保护色, 与周围生活环境融为一体, 有利于规避天敌和捕食猎物; 其次, 体色可以作为一种交流信号, 与其特定的生活生理特征相联系; 再次, 体色的改变可能与其生活环境的变化有关, 如招潮蟹(Uca pugilator)便具有因环境温度高低和光强的强弱而改变身体颜色的特性[19]。
自然界中, 同一物种内具有不同体色个体或群体的现象十分普遍, 针对体色遗传变异等方面的研究也比较深入。郑怀平等[20]根据贝壳颜色的差异对海湾扇贝(Argopecten irradians)家系的建立及其成长发育过程进行了观察研究, 发现体色(壳色)作为一个可遗传的质量性状, 它与表型性状的联系可能与贝类的遗传、生理特性以及生态环境等有关。在本研究所观察到的三疣梭子蟹既然同属于一个种, 那么其体色的变化很可能是由环境因素或者遗传因素引起的。同时在实际观察中了解到, 雌蟹和雄蟹中都有紫色和茶绿色个体的存在, 因此否定了雌蟹为紫色, 雄蟹为茶绿色的说法[4]。另外, 在养殖中虽然有个别紫色个体体色会褪色为接近茶绿色个体体色的现象, 但其与茶绿色体色仍存在着明显的差异, 且同等条件下绝大多数紫色个体颜色不会发生明显变化。因此, 基于以上分析, 可以认为三疣梭子蟹产生紫色和茶绿色两种明显体色差异的原因是遗传和环境因素共同作用的结果, 产生体色差异的具体原因有待进一步地深入研究。
[1] Hamasaki K, Fukunaga K, Kitada S. Batch fecundity of the swimming crab Portunus trituberculatus (Brachyura: Portunidae)[J]. Aquaculture, 2006, 253: 59-365.
[2] 郭天慧, 孔晓瑜, 陈四清, 等. 三疣梭子蟹线粒体DNA 16S rRNA和COI基因片段序列的比较研究[J].中国海洋大学学报, 2004, 34(1): 22-28.
[3] 迟大利, 阎斌伦, 高焕, 等. 三疣梭子蟹分子生物学研究进展[J]. 水产养殖, 2008, 29(4):11-15.
[4] 苏秀榕, 李太武, 欧阳芬, 等. 三疣梭子蟹营养成分的研究[J]. 营养学报, 1996, 18(3): 342-346.
[5] 孙先交. 三疣梭子蟹越冬、育苗及养殖[J]. 齐鲁渔业,2001, 18(4): 18-20.
[6] Avise J C. Phylogeography[M]. London: Harvard Univesity Press, 2000. 447.
[7] Yamauchi M M, Miya M U, Nishida M. Complete mitochondrial DNA sequence of the swimming crab,Portunus trituberculatus (Crustacea: Decapoda:Brachyura)[J]. Gene, 2003, 311: 129-135.
[8] Place A R, Feng X J, Steven C R, et al. Genetic markers in blue crabs (Callinectes sapidus) II: Complete mitochondrial genome sequence and characterization of genetic variation[J]. Journal of Experimental Marine Biology and Ecology, 2005, 319: 15-27.
[9] Bandyopadhyay P K, Stevenson B J, Cady M T, et al.Complete mitochondrial DNA sequence of a Conoidean gastropod, Lophiotoma (Xenuroturris) cerithiformis:Gene order and gastropod phylogeny[J]. Toxicon, 2006,48: 29-43.
[10] 陈丽梅, 孔晓瑜, 喻子牛, 等. 3种蛏类线粒体 16S rRNA和COI基因片段的序列比较及其系统学初步研究[J]. 海洋科学, 2005, 29(8): 27-32.
[11] 徐琰, 宋林生, 李新正. 用 16S rDNA序列初步探讨部分真虾类的系统发育关系[J]. 海洋科学, 2005,29(9): 36-41.
[12] 蒋钦杨, 杨学明, 郭亚芬, 等. 3个不同群体罗氏沼虾线粒体16S rRNA基因序列分析及遗传差异[J]. 水产科学, 2005, 24(10): 28-31.
[13] 郭亚辉, 郭坚华, 李斌. 根据 16S rRNA序列对假单胞菌属分类学的研究进展[J]. 微生物学杂志, 2004,24(2): 38-41.
[14] 刘佳文, 吕红艳, 吴雪琼, 等. 16S rRNA基因序列在分枝杆菌分类鉴定的研究进展[J]. 国外医学临床生物化学与检验学分册, 2001, 22(5): 265-268.
[15] 孔晓瑜, 姜艳艳, 相建海, 等. 魁蚶线粒体16S rRNA和 COI基因片段序列测定及其应用前景[J]. 海洋科学, 2001, 25(12): 46-48.
[16] 张文治. 微生物学[M]. 北京: 高等教育出版社, 2005:p240.
[17] 朱世华, 郑文娟, 邹记兴, 等. 科鱼类线粒体DNA控制区结构及系统发育关系[J]. 动物学研究, 2007,28(6): 606-614.
[18] 郭学光, 郭乔楠. 线粒体 DNA 控制区研究进展[J].肿瘤防治杂志, 2005, 12(22): 1747-1750.
[19] Silbiger N, Munguia P. Carapace color change in Uca pugilator as a response to temperature[J]. Journal of Experimental Marine Biology and Ecology, 2008,335: 41-46.
[20] 郑怀平, 张国范, 刘晓, 等. 不同贝壳颜色海湾扇贝(Argopecten irradians)家系的建立及生长发育研究[J].海洋与湖沼, 2003, 34(6): 632-639.
Received: Sep., 10, 2009
Key words:Portunus trituberculatus; boby-color, mitochondrial DNA; 16S rRNA; COI
Abstract:The blue swimming crabs, an important commercial species, with different body-colors were discovered along the coast of East China in recent years. Both purple and tea-green crabs have similar, even the same physical characteristics except for the carapace color. In order to identify the relationships between them,we amplified and compared two conserved mitochondrial gene fragments of color-different crabs through DNA sequencing analyses. The sequence alignment analysis of color-different crabs presented 99.87% and 99.88% nucleotide sequence identity for the COI and 16S rRNA genes, respectively, indicating that there was no species or subspecies differentiation between them. Based on the alignment of nucleotide sequences, a phylogenetic tree of 16S rRNA genes from 16 species of Portunidae was constructed. It may serve as a basis for the taxonomy of portunid crabs.
(本文编辑:张培新)
Comparison analysis between purple and tea-green individuals of Portunus trituberculatus using mitochondrial partial genes
CHI Da-li1,2, GAO Huan1, SHEN Song-dong2, YAN Bin-lun1
(1. Jiangsu Key Laboratory of Marine Biotechnology, Huaihai Institute of Technology, Lianyungang 222005,China; 2. Life Sciences College, Suzhou University, Suzhou 215123, China)
Q178.532
A
1000-3096(2010)11-0027-08
2009-09-10;
2009-11-12
“十一五”国家科技支撑计划重大项目(2006BAD09A01)作者简介: 迟大利(1983-), 男, 山东龙口人, 硕士研究生, 从事三疣梭子蟹体色相关分子标记研究, E-mail: charliedl@163.com; 通信作者: 阎斌伦, 教授, 从事虾蟹类养殖工作, 电话: 0518-85895252, E-mail:
yanbinlun@yahoo.com.cn
猜你喜欢
科学养鱼(2022年9期)2022-10-18 01:49:54
当代水产(2019年7期)2019-09-03 01:02:22
学生天地(2017年24期)2017-11-07 01:47:43
红蜻蜓·低年级(2016年10期)2016-12-03 10:45:28
江苏农业科学(2016年5期)2016-07-23 19:00:27
新作文·小学低年级版(2015年5期)2015-05-18 04:57:51
热带农业科学(2014年8期)2014-09-23 18:59:14
少儿科学周刊·儿童版(2014年1期)2014-03-10 06:21:21
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28 12:21:38
餐饮世界(2013年3期)2013-08-16 07:48:36
