
患灰霉病仙客来植株的解剖观察
2010-09-20 00:24:02王继佳李凤兰孙丽丽
东北农业大学学报 2010年2期
王继佳,李凤兰,韩 峰,孙丽丽,金 峰
(东北农业大学生命科学学院,哈尔滨 150030)
仙客来(Cyclamen premium Mill.)属报春花科仙客来属的多年生球根草本植物,是世界著名的节日小盆花之一。仙客来灰霉病是温室中常见的真菌病害[1],其病原为富克尔核盘菌,属子囊菌门真菌,由气流传播,主要通过伤口侵入植物体。该病主要发生在叶片、叶柄、花梗和花瓣上[2]。据2005年调查,仙客来灰霉病在1~3月低温时发病率高达90%,对花卉种子的产量和品质均有很大影响[3]。近几年来,由于连年栽培、菌源积累等原因,仙客来的灰霉病病害越来越重,已成为影响仙客来欣赏价值和商品价值的关键因素。有关仙客来灰霉病的研究主要集中在病原鉴定及诊断和防治等方面[3-5],而对患病植株营养器官内部变化的研究则少有报道。
本文运用石蜡切片技术对患灰霉病仙客来植株的叶片、叶柄、花、花梗进行了解剖学观察,为进一步研究仙客来灰霉病的起源、发生、传播及其防治提供解剖学依据。
1 材料与方法
1.1 材料
仙客来的正常植株和染灰霉病的典型病株为同一品种,其来源于东北农业大学植物学实验室。
1.2 方法
选取仙客来染病植株具有代表性的叶片(长0.5 cm,宽0.3 cm)、叶柄(0.5 cm)、花(长0.5 cm,宽0.3 cm)、花梗(0.5 cm),用FAA固定24 h以上,常规石蜡切片法制片,切片厚度为l0 μm,番红-固绿双重染色,封片后,于OLYMPUS光学显微镜下观察并拍照。
本研究选择6种组织内的细胞,在显微镜下每种组织随机选取5个视野,观察每个视野中含有菌丝或孢子的细胞比例,并计算出在此组织中的平均数。
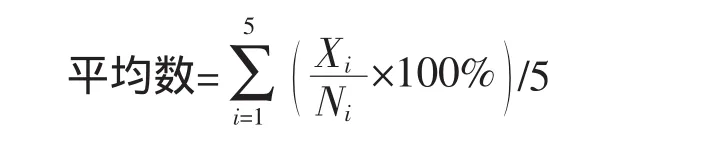
2 结果与分析
2.1 患病植株的外部形态特征
对正常仙客来植株和患灰霉病植株外观形态的对比观察,发现灰霉病主要危害叶片、叶柄及花冠(见图1、2)。由观察结果可以看出,灰霉病主要危害叶片、叶柄及花冠。染病叶片表面有暗绿色水渍状病斑,干燥条件下叶斑呈“V”字形褐斑,看不见灰霉;叶柄和花梗呈水渍状腐烂;花瓣发病时变色,白色品种花瓣变成淡褐色,红色品种的花瓣退色,并出现水渍状圆斑,病重时花瓣腐烂,密生灰色霉层,即病原菌分生孢子梗或分生孢子,影响观赏性,这些特点与已报道结果一致[4]。

图1 正常仙客来植株的叶片和花瓣Fig.1 Leaves and flowers of normal cyclamen plant

图2 感病仙客来植株的叶片和花瓣Fig.2 Leaves and flowers of cyclamen plant with Botrytis cinerea infection
2.2 灰霉病病原菌对仙客来器官的侵染方式
在番红-固绿的染色条件下,灰霉病病原菌的菌丝被染成绿色,分生孢子被染成红色[6]。对患病的不同器官进行解剖学观察发现,病原菌主要通过器官的表皮进行侵染,但不同器官的侵染过程有所差别。从患病叶片来看,病原菌大部分是以菌丝直接通过叶片下表皮细胞进入叶片组织,并在表皮细胞中产生大量分生孢子,从而破坏表皮细胞(见图3)。
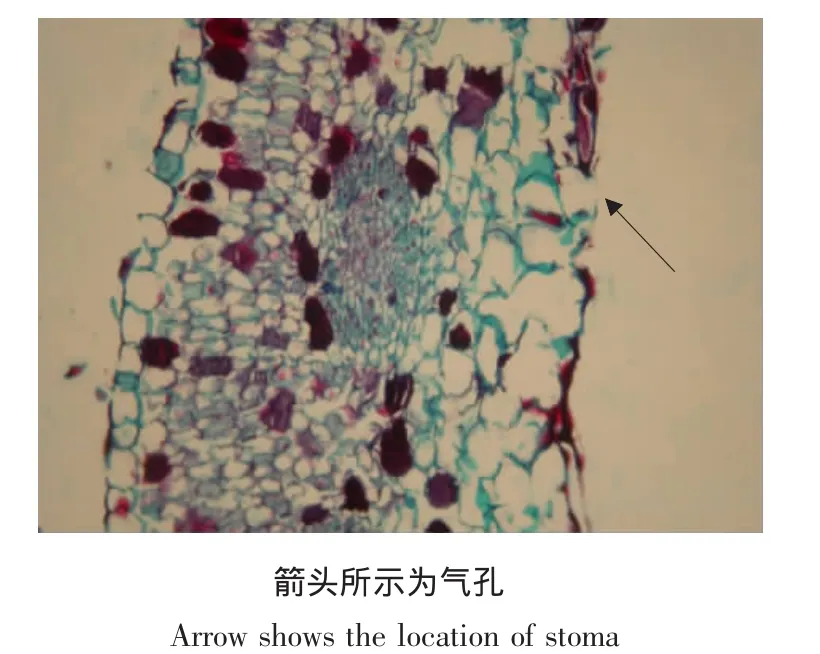
图3 感病仙客来叶片横切面(×100)Fig.3 Transverse section of cyclamen leaves with Botrytis cinerea infection(×100)
病原菌对花瓣的侵染过程与之相似,也是首先在表皮细胞中产生了大量菌丝,继而侵染内部组织细胞(见图4)。病原菌对仙客来叶柄和花梗的侵染,都是通过表皮进行的,其破坏表皮细胞的方式同叶片相似,即在表皮细胞中产生大量分生孢子(见图5)。

图4 感病仙客来花瓣横切面(×400)Fig.4 Transverse section of cyclamen flowers with Botrytis cinerea infection(×400)
2.3 患灰霉病仙客来植株中的病原菌分布及其存在状态
2.3.1 灰霉病病原菌在叶片中的分布及其存在状态

图5 感病仙客来花梗纵切面(×100)Fig.5 Vertical section of cyclamen pedicels with Botrytis cinerea infection(×100)
由表1可知,灰霉病病原菌的菌丝及分生孢子在叶片不同组织中的分布有很大差异。上下表皮中菌丝分布量较多,而其他组织中菌丝分布较少,尤其在韧皮部中,几乎没有菌丝存在;分生孢子在叶片的下表皮中分布量较大,并且密集度达到最高,为44.0%,而上表皮中的分生孢子量却很少,只有6.0%,这与解剖结构观察到病原菌通过叶片下表皮进行侵染的结论相一致。在叶脉的木质部和韧皮部中也有较多的分生孢子存在(见图3)。

表1 灰霉病病原菌在叶片中的分布及其存在状态Table1 Distribution and state of pathogens in cyclamen leaves (%)
2.3.2 灰霉病病原菌在叶柄中的分布及其存在状态
由表2可知,菌丝在叶柄中主要分布于表皮、皮层和韧皮部,其中在韧皮部中的含量最多,达到了极显著的水平,而木质部和髓中几乎没有菌丝的存在。在叶柄的不同组织细胞中都有病原菌的分生孢子,分生孢子呈圆形。叶柄表皮细胞中的分生孢子数量最多,达到40.6%,明显高于其他组织,其中木质部中的含量最少。叶柄髓部受病原菌感染的程度最低(见图6)。

表2 灰霉病病原菌在叶柄中的分布及其存在状态Table2 Distribution and state of pathogens in the cyclamen petioles (%)
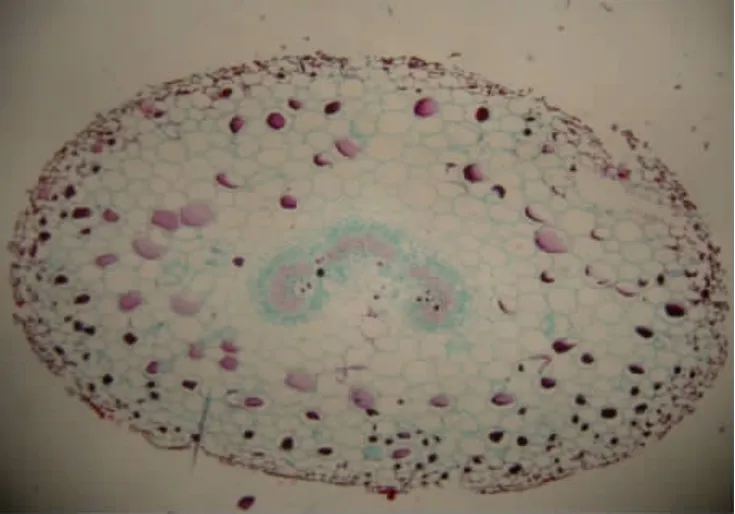
图6 感病仙客来叶柄横切面(×40)Fig.6 Transverse section of cyclamen petioles with Botrytis cinerea infection(×40)
2.3.3 灰霉病病原菌在花瓣中的分布及其存在状态
由表3可知,花瓣表皮细胞中存在着较多的丝状菌丝和分生孢子,而薄壁细胞中分生孢子和菌丝的含量都较少。在花瓣维管束中分生孢子最多,为41.3%,但菌丝的含量最少,只有11.0%(见图7)。这同灰霉病病原菌在叶片和叶柄各组织中的分布趋势相一致。

表3 灰霉病病原菌在花瓣中的分布及其存在状态Table3 Distribution and state of pathogens in cyclamen flowers (%)
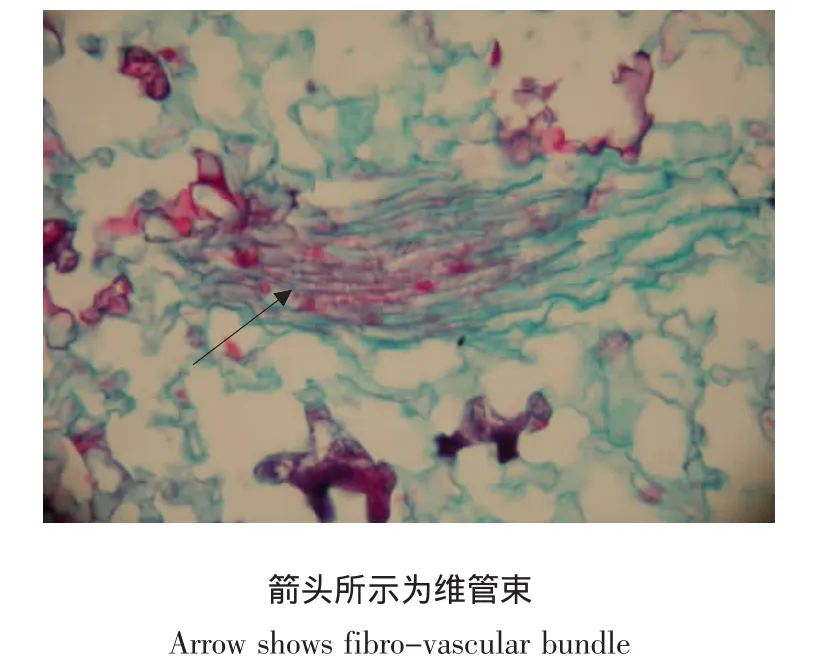
图7 感病仙客来花瓣横切面(×400)Fig.7 Transverse section of cyclamen flowers with Botrytis cinerea infection(×400)
2.3.4 灰霉病病原菌在花梗中的分布及其存在状态
由表4可知,在花梗中,几乎没有菌丝,病原菌主要以分生孢子的形式存在,其中表皮细胞中的分生孢子数量最多,达49.8%,而韧皮部中的量最低,仅为2.0%(见图8)。
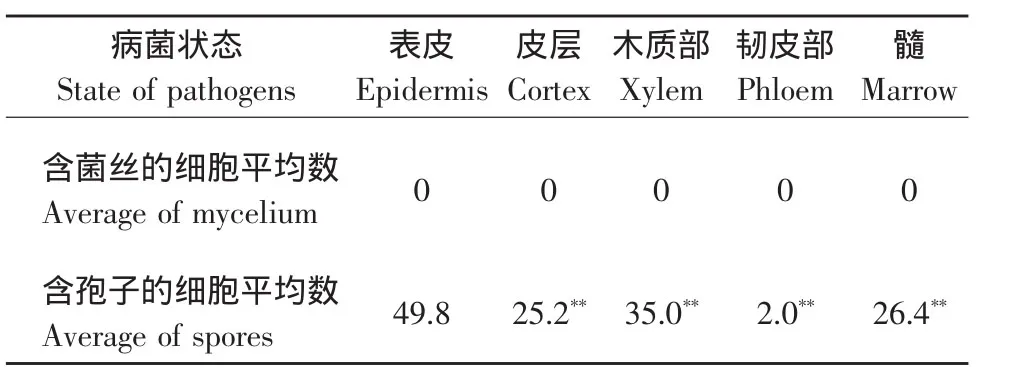
表4 灰霉病病原菌在花梗中的分布及其存在状态Table4 Distribution and state of pathogens in cyclamen pedicels (%)
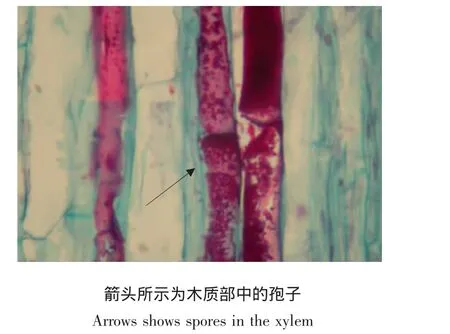
图8 感病仙客来花梗纵切面(×400)Fig.8 Vertical section of cyclamen pedicels with Botrytis cinerea infection(×400)
3 讨论与结论
3.1 石蜡切片技术在患灰霉病仙客来器官观察中的应用
对受到真菌病害侵染的植株内部变化的观察方法主要有细胞、亚细胞水平、生理生化以及基因分子水平的方法[7]。本试验采用石蜡切片技术结合番红-固绿染色方法对仙客来患病植株的器官进行解剖学观察,主要是从细胞水平上揭示仙客来感染灰霉病后其感病器官的解剖结构特征。由于被真菌感染的细胞壁角质化加厚[8],经番红-固绿染色后呈红色,因此有利于对感病细胞的确定和观察;此外,细胞中的分生孢子和菌丝在染色中呈现出不同的颜色反应,其中分生孢子易被番红染成红色,而菌丝则被固绿染成绿色,也便于试验结果的观察,因此,石蜡切片方法可以用于受真菌侵染植株的内部变化的观察。
3.2 仙客来植株被灰霉病病原菌侵染方式和位置
植物病原真菌侵入寄主并在寄主组织中生长是其致病的先决条件[8]。病原菌在入侵寄主时,通常会产生一些由特化的菌丝形成的侵染结构,如:侵染垫[9-10]、附着胞[11]、侵染钉和吸器[12-13]等来帮助入侵并与寄主建立寄生关系。但是,侵染垫和附着胞并不是侵入所必需的[14]。所以病原菌的侵染可归纳为以侵染垫或裂片状附着胞侵染、不产生专门侵染结构由菌丝顶端直接穿透植物表面或从自然孔口(主要是气孔)及伤口侵入几种情况[15-16]。在本试验中,感病仙客来的不同器官中都没有发现菌丝的特化结构,可能是由于所选取试验材料已经过了感染的最初期,但是在观察过程中发现,灰霉病病原菌在叶片、叶柄、花瓣和花梗中的分布特点比较一致,即表皮部位病原菌的密集度最高,表明灰霉病病原菌可能是通过表皮进行侵染,被侵染的叶片中可以观察到有很多病原体的菌丝是直接通过叶片下表皮的气孔进入植株组织的。在真菌侵染的表皮部位,都含有大量分生孢子,这种分布既有利于真菌侵染组织细胞,也有利于孢子向外传播。至于在侵染初期是否产生了其他特化结构,还需进一步观察和研究。
3.3 灰霉病病原菌在仙客来器官中的传播
蒋军喜等对真菌侵染植物的研究发现,游动孢子在侵入植物时先回缩鞭毛,接着将自身固定在植物表面,然后侵入寄主细胞质中[17]。本试验中有些患病器官的解剖结构显示,当孢子从表皮进入皮层后,先在皮层细胞内定殖并形成大量菌丝,然后进一步向皮层和其他组织细胞扩展,而且真菌菌丝在皮层内的扩展方向一般是先横向后纵向。导管中的分生孢子可沿导管纵向扩展,同时分生孢子还可穿越导管进行横向扩展,因此,在重病植株体内,分生孢子量比较大的导管周围的细胞中,往往也存在着大量分生孢子,说明病原菌在病株体内的纵、横向扩展,可能是由分生孢子本身不断生长发育来完成的,但具体的机制有待于进一步研究。分生孢子纵横向扩展时,在植株体内的某一部位还能因其大量产生而使导管填塞,进而导致花梗和花瓣枯萎死亡。
[1]原国辉,高一凤,程玉琴,等.仙客来几种主要病害的识别与防治[J].农业科技通讯,2001(2):26-27.
[2]孙文英,周丽.仙客来主要病害诊断与防控措施[J].辽宁农业科学,2007(5):45-46.
[3]张蕾,王福银,李阳春,等.仙客来灰霉病的病原鉴定及其生物学特性研究[J].安徽农学通报,2008,14(1):168-171.
[4]王利平.仙客来灰霉病的诊断及防治[J].技术与市场,2006(11):32.
[5]雷玉明.仙客来灰霉病的诊断及防治[J].中国花卉盆景,2002(5):41.
[6]王建明,郑经武,贺运春,等.西瓜枯萎病菌在植株体内的存在状态、数量分布及扩展规律的研究[J].中国农业科学,1993,26(3):69-74.
[7]郑玲,吴小芹.植物病原真菌侵染结构研究进展[J].南京林业大学学报:自然科学版,2007(1):90-94.
[8]谭小明,郭顺星.红豆杉根的显微结构及其内生真菌分布[J].中国医学科学院学报,2006(6):372-374.
[9]Dodman R L,Barker K R,Walker J C.A detailed study of the different modes of penetration by Rhizoctonia sulani[J].Phytopathology,1968,58:1271-1276.
[10]Dodman R L,Flentje N T.The mechanism and physiology of plant penetration by Rhizoctonia solani[J].Biology and Pathology,1970(5):149-160.
[11]Emmett R W,Parbery D G.Appressoria[J].Ann Rev Phytopathol,1975,13:147-167.
[12]Riopel J L,Timko M P.Haustorial initiation and differentiation[M]//Press M C,Graves J D.Parasitic plants.New York:Chapman and Hall Press,1995:39-79.
[13]黄国红,康振生,朱之,等.小麦叶锈菌在感病寄主上发育的组织病理学和超微结构研究[J].植物病理学报,2003,33(1):52-56.
[14]张红生,朱立宏,沙学延,等.水稻纹枯病抗性机理的初步研究[M]//朱立宏.主要农作物抗病性研究进展.南京:江苏科技出版社,1990:153-164.
[16]Deising H B,Wnener S.The role of fungal appressoriain plant infection[J].Microbes and in Infection,2000,2(13):1631-1641.
[16]Marshall D S,Rush M C.Relation between infection by Rhizoctonia sulani and R.oryzae and disease severity in rice[J].Phytopathology,1980,70:941-946.
[17]蒋军喜,蔡祝南.植物病毒的真菌传播及传毒介体油壶菌研究进展[J].江西农业大学学报,2003,25(3):362-365.
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31 06:38:50
小天使·四年级语数英综合(2022年6期)2022-06-21 12:49:36
今日农业(2021年21期)2022-01-12 06:31:36
蔬菜(2021年3期)2021-11-26 09:36:08
作文小学中年级(2021年4期)2021-05-07 01:17:50
文苑·经典美文(2019年9期)2019-09-09 05:56:44
疯狂英语·新读写(2018年2期)2018-09-07 09:32:10
新农业(2016年15期)2016-08-16 03:40:40
农家科技中旬版(2016年12期)2016-04-16 03:41:23
现代农业(2016年5期)2016-02-28 18:42:37
