
O139,142,26混合血清型大肠杆菌的O-抗原基因簇的破译
2010-07-09 01:36:54许亚卓杨春柳刘文鑫杨旭东李海滨师东方
东北农业大学学报 2010年2期
许亚卓,杨春柳,刘文鑫,杨旭东,李海滨,师东方
(东北农业大学动物医学学院,哈尔滨 150030)
大肠杆菌的不同血清型分类是由大肠杆菌O-抗原决定的。大肠杆菌O-抗原是大肠杆菌细胞最外侧结构,由多个重复的单糖单位组成,是构成大肠杆菌脂多糖的重要组成部分[1]。其单糖种类、数量以及单糖间键,寡糖单位之间键的不同构成了O-抗原的多样性,根据O-抗原进行分类,大肠杆菌现已有180多种不同血清型[2-4]。O-抗原种类虽多,但都有固定结构模式,O-抗原基因一般分成三部分:单糖合成酶基因;糖基转移酶基因和寡糖单位处理酶基因。这些结构基因都位于galF和gnd 2个看家基因之间。galF基因和gnd基因分别位于O-抗原基因簇的5′末端和3′末端,它们不属于O-抗原基因簇[5]。这两个看家基因在不同血清型大肠杆菌中都有相对保守的部分,比较稳定,可用来扩增O-抗原基因簇[3]。
师东方等从猪水肿病病死猪肠系膜淋巴结中分离到了一株O139,142,26混合血清型大肠杆菌[6],这与以往的研究中曾发现过血清型交叉反应现象相符,Lothar等发现大肠杆菌O123的两个H型分别与O4、O12发生交叉血清型反应[7],O142与O106在发生了一个羧基改变后也发生了交叉血清型反应现象[8],在牦牛体内[9]和试验兔体内[10]分离到混合血清型大肠杆菌,出现2个或3个血清型交叉反应,以及其他菌属与大肠杆菌之间都发生过血清型交叉反应现象[11]。本文通过鸟枪法扩增了O139,142,26混合血清型大肠杆菌分离株O-抗原基因,利用分子生物学软件对其O-抗原基因簇进行分析,为进一步研究混合血清型大肠杆菌分子生物学特性提供依据。
1 材料与方法
1.1 材料
1.1.1 菌种
O139,142,26混合血清型大肠杆菌,大肠杆菌TGⅠ由本实验室保存。
1.1.2 质粒和试剂
PMD18-T Vector、PCR纯化试剂盒购自Sigma公司、HiFiPCR酶、EasyTaq酶、dNTP购自北京全氏金公司;Eco R I、DNaseI购自日本TaKaRa公司;引物由上海生工生物技术公司合成;氨苄青霉素、氯仿、异戊醇、乙醚、醋酸钠等常规试剂购自天津市恒兴化学试剂制造有限公司。
1.2 方法
1.2.1 O-抗原基因簇的获得
根据galF基因设计上游引物P1:5′ATTGTG GCTGCAGGGATCAAAGAAAT 3′,根据gnd基因设计下游引物P2:5′TAGTCGCGTGNGCCTGGATTA AGTTCGC 3′。提取该大肠杆菌基因组DNA做为模板[1],用长距离PCR方法扩增O-抗原基因簇。PCR反应条件为:94℃预变性5 min,94℃变性30 s,56℃退火30 s,70℃延伸10 min,30个循环,终延伸为70℃,7 min,反应体系为50 μL。合并8管PCR产物,用PCR纯化试剂盒纯化。
1.2.2 O-抗原基因文库的构建
用DNaseI消化PCR纯化产物,1∶8 000稀释DNaseI,在10 μL体系中加入7 μL PCR纯化产物,1 μL 0.5 mol·L-1Tris-HCl(pH=7.5)、1 μL MnCl2(0.1 mol·L-1),反应 10 min,用 1 μL 5 mol·L-1EDTA,在65℃终止反应。使其酶切片段在500~3 000 bp之间,合并反应产物,用氯仿∶异戊醇和醋酸钠对酶切产物进行纯化。用PCR方法对酶切片段补平加A,再经氯仿∶异戊醇和乙醚纯化,产物与pMD18-T载体在16℃连接24 h,取连接产物转化感受态大肠杆菌TGⅠ,涂布在含有氨苄青霉素的LB固体培养基上,37℃过夜培养,以碱裂解法大规模提质粒,用Eco RⅠ酶切鉴定其中的插入片段的大小,挑选插入片段在500~3 000 bp的克隆。为了保证序列的准确性,避免PCR反应本身的错配,根据帕松公式(P=e-m,P-未测定碱基的概率;m-测定序列的覆盖率)[12],挑取了50个插入片段为500~3 000 bp的克隆进行测序拼接,使每个碱基至少有3个以上高质量(>90%)的序列覆盖,最终得到全基因序列。
2 结果与分析
2.1 O-抗原基因簇的获得
根据galF和gnd基因设计上下游引物,用长PCR方法扩增O-抗原基因簇。长PCR产物通过0.8%琼脂糖凝胶电泳检测,在接近15 kb的位置出现一条电泳带。将PCR产物酶切后,按前述方法挑取插入片段500~3 000 bp的克隆,测序拼接,最终获得了大肠杆菌O139,142,26的O-抗原基因簇的全序列,序列全长11 771 bp。
2.2 O-抗原基因簇分析
在获得混合O-抗原基因簇的全序列后,用ORF finder发现基因,共找到10个开放阅读框(ORF),都位于galF基因和gnd基因之间且具有相同的转录方向,都是从5′~3′方向。用Artemis软件完成基因注释后得到大肠杆菌O139,142,26的O-抗原基因簇结构及其(G+C)%含量,如图1所示。
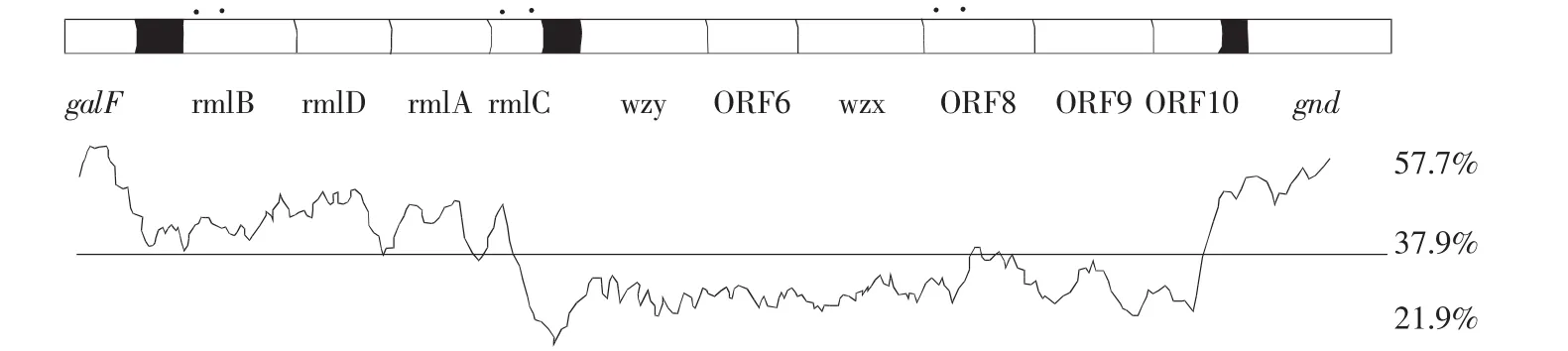
图1 大肠杆菌O139,142,26的O-抗原基因簇结构
galF基因和gnd基因分别位于O-抗原基因簇的5′末端和3′末端,它们不属于O-抗原基因簇[5]。除这两组基因外O-抗原基因簇还含有3种酶基因,即单糖合成基因,糖基转移酶基因和寡糖单位处理基因[3]。
2.2.1 单糖合成酶基因
通过进行BLAST比较发现,ORF1、ORF2、ORF3和ORF4都与许多菌的合成鼠李糖前体的4个合成酶基因有非常高的相似性,其中与大肠杆菌O139的rmlB、rmlD、rmlA、rmlC相似性最高,分别为97.63%、98.45%、99.66%、98.15%,并且都与鲍氏志贺氏菌的rml有97%以上相似性。可以确定 ORF1、ORF2、ORF3、ORF4分别为合成dTDP-鼠李糖的rmlB、rmlD、rmlA、rmlC基因,因此分别命名为rmlB、rmlD、rmlA、rmlC。
2.2.2 寡糖单位处理基因
通过TMHMM和SMART分析,ORF5和ORF7是仅有的2个具有多个以上跨膜蛋白的序列,ORF5有11个跨膜蛋白,并在第2和第3个跨膜蛋白之间有一个含92个氨基酸残基的大型胞质茎环,这是wzy跨膜蛋白的典型结构[13],经过BLAST保守蛋白分析,与大肠杆菌O139的O-抗原跨膜基因相似性达到99.75%;ORF7有9个跨膜蛋白,也具有高度特异性,与大肠杆菌O139O-抗原跨膜基因相似性达到99.43%。因此将ORF5与ORF7分别命名为wzy、wzx。
2.2.3 糖基转移酶基因
经过BLAST分析,ORF6、ORF8、ORF9、ORF10与大肠杆菌O139编码糖基转移酶wfaI、wfaJ、wfaK、wfaL,相似性分别为99.2%、98.88%、99.23%、98.7%,推测它们属于糖基转移酶家族类蛋白,但不能确定其作用的糖苷健类型,与其他菌也有一定的相似性,如ORF9与假单胞杆菌的wbpR糖基转移酶的相同性为36%,相似性为53%,所以根据其特点及与已发表的O139抗原基因有较高的同源性将ORF6、ORF8、ORF9、ORF10 命名为 wfaI、wfaJ、wfaK、wfaL[14]。
通过对每个ORF用BLAST系列软件与Gen-Bank中的基因比较预测出它们的功能如表1所示。
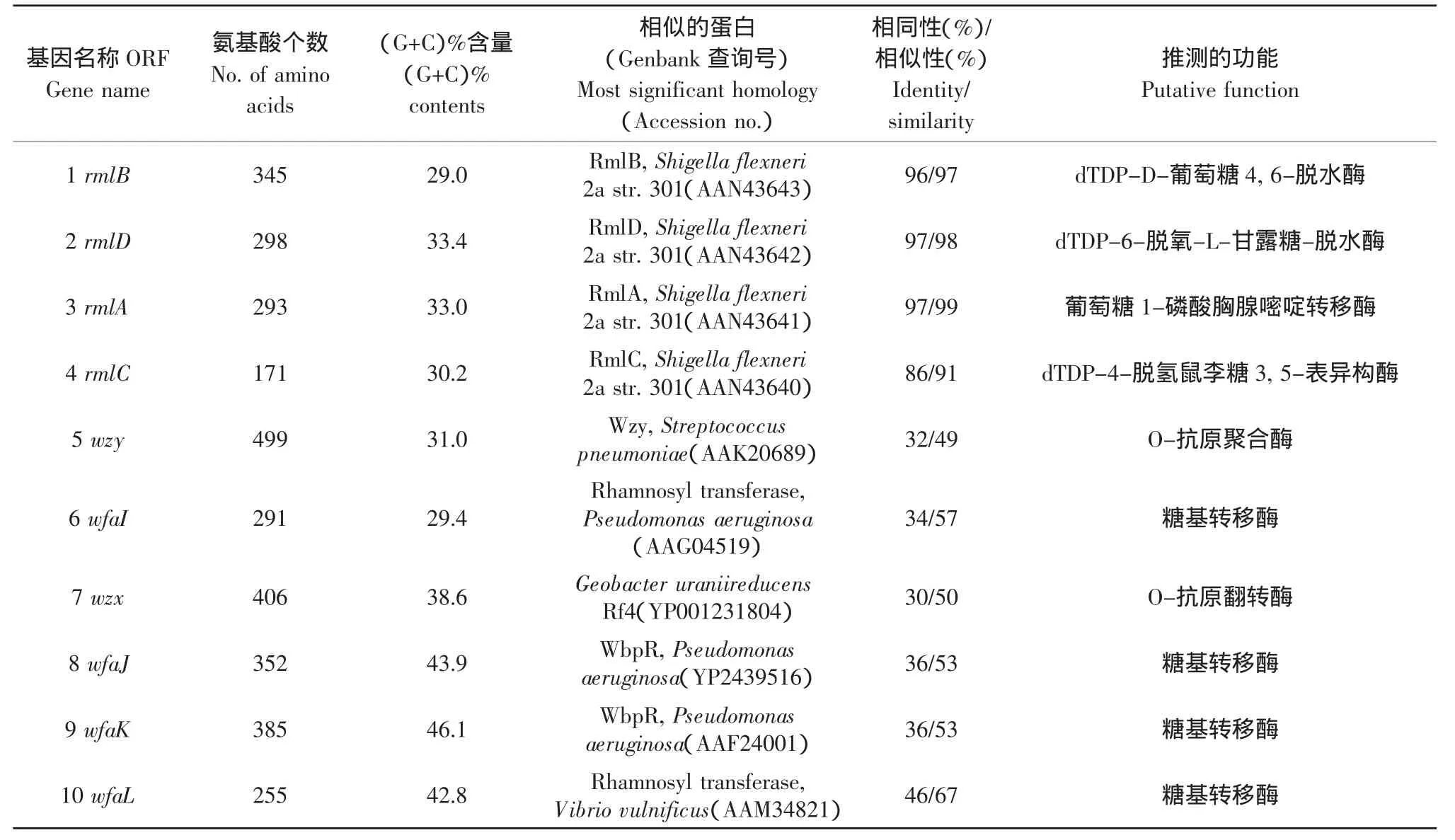
表1 O139,142,26大肠杆菌O-抗原基因簇中基因的功能Table 1 Function of genes on E.coli O139,142,26O-antigen gene cluster
2.3 大肠杆菌O139、O26及O142的O-抗原基因簇及糖基链的比较
根据GenBank中发表的 O139(序列号 DQ109 552)、O26(序列号AY763106)基因序列可知,除wzx与wzy 2个特殊的跨膜蛋白合成基因具有较高的特异性以外,其他开放阅读框都有部分相似性,如它们都含有合成鼠李糖前体的4个基因rmlB、rmlD、rmlA、rmlC,并且相似性都在93%以上,在其他的糖基转移酶中O139的wfaI、wfaJ糖基转移酶基因与O26的糖基转移酶基因fnl2、fnl1相似性为33.15%和33.5%,O139的wfaK、wfaL糖基转移酶基因与O26的糖基转移酶基因wbuB、wbuA相似性为30.95%和40.15%。
瑞典学者Landersjo等[15]对O142和O139的O-抗原外部糖基结构进行了测定,发现了它们重复的单糖结构。O142O-抗原含有 α-D-GalpNAc、α-L-Rhap、β-D-GlcpNAc;O139O-抗原含有 α-D-GlcpNAc、α-L-Rhap、α-D-GalpA、β-D-Glcp,它们的糖基大部分成分相同。比较O142O-抗原基因序列[16],发现大肠杆菌O142也含有鼠李糖前体合成基因rmlB、rmlD、rmlA、rmlC,并且与O139相似性很高。
通过DNAMAN比对基因相似性,大肠杆菌O139,142,26与O139(序列号DQ109552)相似性为99.7%,与O26(序列号AY763106)相似性仅为44.15%。与大肠杆菌O139相比,O139,142,26全基因序列共有32处不同,各段开放阅读框相似性都在99%以上,除了合成鼠李糖前体的单糖合成酶基因(rmlB、rmlD、rmlA、rmlC)有21处不同外,就只有O139,142,26糖基转移酶基因wfaL与O139的糖基转移酶基因wfaL段出现了最多的6处不同,是相对其他片段变异较大的。而与大肠杆菌O26相比,除合成鼠礼糖前体的单糖合成酶基因相似性在75%以上,其他片段比对结果同大肠杆菌O139与O26比对结果相同,其中O139,142,26的糖基转移酶基因wfaL与O26糖基转移酶基因wbuA的相似性为40.15%,是相对于其他片段相似性较高的。
2.4 wfaL基因的进化分析
由于大肠杆菌O139,142,26与O139,的wfaL片段具有相对较高的变异性,大肠杆菌O139,142,26的wfaL片段与O26的wbuA片段具有相对较高的相似性,所以预通过wfaL氨基酸序列的进化关系来分析大肠杆菌O139,142,26、O139,、O26O-抗原之间的进化关系。经BLAST protein分析寻找wfaL的相似序列,从这些相似序列中选择了包括O139和O26在内的value值在5e-18以上的共17个序列,用MEGA进化树分析它们的同源性(见图2)。将O139,142,26暂时命名为E.mix,其他命名都为简写。从图2中可以看出O139,142,26与O139,落在一个分支里,和R.JMP134(蛋白序列号为YP_294950)是同一个祖先,都是糖基转移酶家族蛋白的成员,而O26和Psychromonas ingrahamii 37(蛋白序列号为YP_944740)落在同一分支里,与O139,142,26相距较远,属于糖基转移酶类似蛋白。因此,大肠杆菌O139,142,26的wfaL很可能是由大肠杆菌O139的wfaL单独演变而来的。
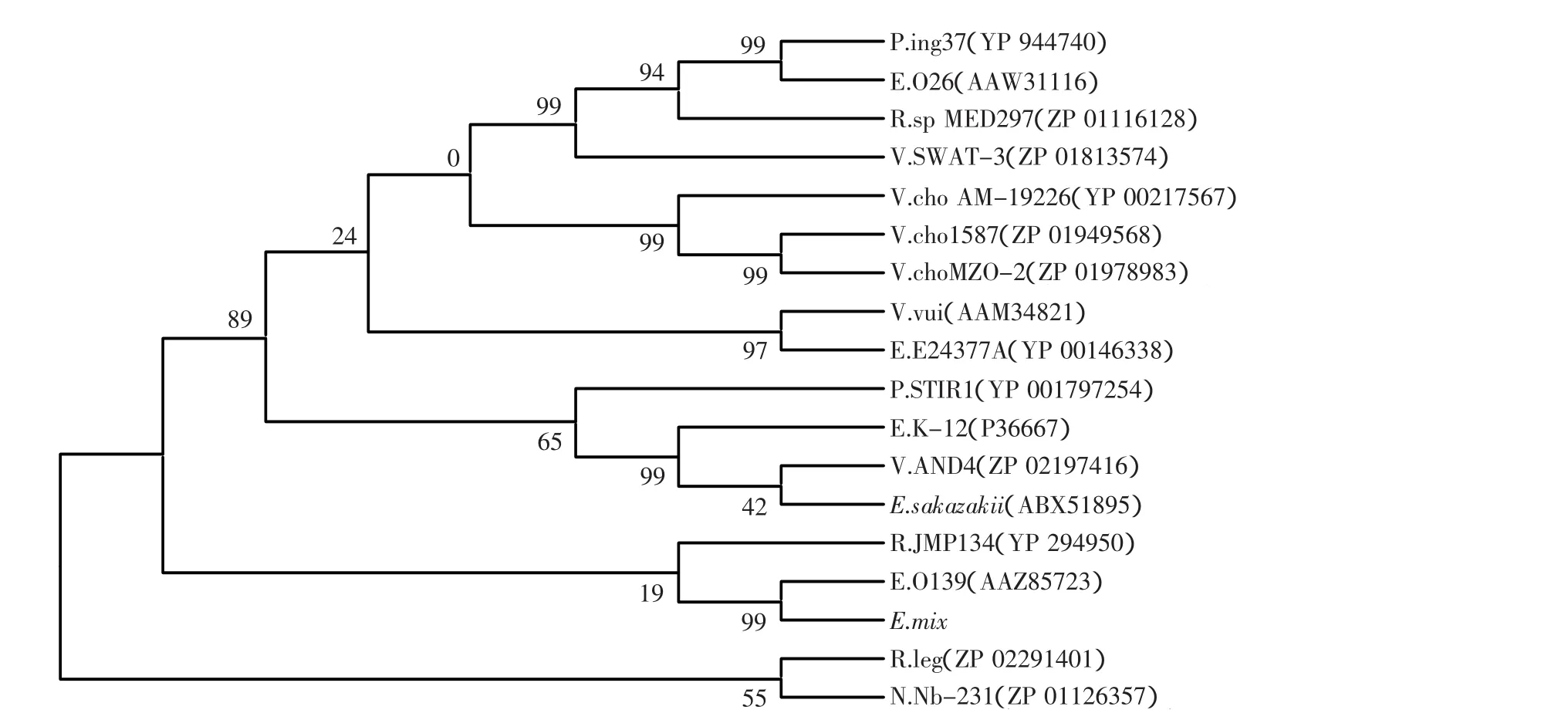
图2 根据相似蛋白序列构建的wfaL系统近化树Fig.2 Phylogenetic trees based on wfaL proteins sequences showing the relationships numbers in parentheses represent the accession number of the protain sequences in GenBank;the number at each branch points of the percentage supported by bootstrap
3 讨论与结论
在O-抗原基因簇中,寡糖单位处理酶基因包括O-抗原转运酶基因wzx和O-抗原聚合酶基因wzy,由于wzx、wzy跨膜基因的高度特异性,近年来有学者用分段设计wzx、wzy序列引物的PCR方法来快速准确地鉴定大肠杆菌的血清型[17-19],说明这两个基因片断对O-抗原免疫原性的形成具有决定性作用。但是由于O139,142,26和O139大肠杆菌的wzx、wzy跨膜基因相似性很高,所以用PCR方法很难将它们区分开。根据二级结构预测抗原决定簇方法[20],分析O139,142,26与O139跨膜基因,预测到wzy的第99位和第330位氨基酸的突变部位很可能处于抗原决定簇内,这些突变很可能改变wzy跨膜蛋白的性质,如拓扑学变化,解旋及亲水集团的转移等,从而使形成的糖前体在跨膜转运时发生变化,引起O-抗原糖基链的改变,从而出现混合血清型现象。以上对大肠杆菌O-抗原混合血清型产生的原因分析还需进一步研究证明。
[1]Bastin D A,Reeves P R.Sequence and analysis of the O antigen gene(rfb)cluster of Escherichia coli O111[J].Gene,1995,164(1):17-23.
[2]Whitfield C.Biosynthesis of lipopolysaccharide O antigens[J].Trends in Microbiology,1995,3:178-185.
[3]Reeves P R,Wang L.Genomic organization of LPS-specificloci[J].Curr Top Microbiol Immunol,2002,264(1):109-35.
[4]Stenutz R,Weintraub A,Widmalm G.The structures of Escherichia coli O-polysaccharide antigens[J].FEMS Microbiol Rev,2006,30:382-403.
[5]Marolda C L,Valvano M A.The GalF protein of Escherichia coli is not a UDP-glucose pyrophosphorylase but interacts with the GalU protein possibly to regulate cellular levels of UDP-glucose[J].Mol Microbiol,1996,22:827-840.
[6]师东方,葛俊伟,王明翠,等.一株混合O血清型猪水肿病大肠杆菌的分离与鉴定[J].中国预防兽医学报,2009(5):397-355.
[7]Lothar B,Wang Q,Dieter N,et al.Relationship between O-antigen subtypes,bacterial surface structures and O-antigen gene clusters in Escherichia coli O123strains carrying genes for Shiga toxins and intimin[J].Journal of Medical Microbiology,2007,56:177-1840.
[8]Hanna K,Jonsson M,Weintraub A,et al.Structural studies of the O-antigenic polysaccharides from Shigella dysenteriae type 3 and Escherichia coli O124,a reinvestigation[J].Carbohydrate Research,2006,341:2986-2989.
[9]索朗斯珠,曾群辉,查果,等.西藏牦牛大肠埃希氏菌的分离鉴定与毒力测定[J].西北农林科技大学学报,2005,33(9):19-23.
[10]王云峰,王翠兰,崔尚金.兔致病性大肠杆菌生物学特性的研究[J].中国预防兽医学报,1999,21(5):354-355.
[11]Guang Z,Perepelov A V,Senchenkova S N,et al.Structural relation of the antigenic poly-saccharides of Escherichia coli O40,Shigella dysenteriae type 9,and E.coli K47[J].Carbohydrate Research.2007,342:1275-1279.
[12]徐建国.分子医学细菌学[M].北京:科学出版社,2000.
[13]DanielsC,VindurampulleC,MoronaR.Overexpressionandtopology of the Shigella flexneri O-antigen polymerase(Rfc/Wzy)[J].Mol Microbiol,1998,28:1211-1222.
[14]Wang L,Liu B,Kong Q,et al.Molecular markers for detection of pathogenic Escherichia coli strains belonging to serogroups O138and O139[J].Veterinary Microbiology,2005,111:181-190.
[15]Landersjo C,Weintraub A,Widmalm G.Structural analysis of the O-antigenic polysaccharide from the enteropathogenic Escherichia coli O142[J].Eur J Biochem,1997,244:449-453.
[16]孔庆科.大肠杆菌O138O-抗原基因簇的破译及糖合成酶基因的功能鉴定[D].天津:南开大学,2004:79-80.
[17]Wang L,Reeves P R.Organization of Escherichiacoli O157O antigen gene cluster and identification of its specific genes[J].Infection and Immunity,1998(9):3545-3551.
[18]王威,彭霞,王荃,等.大肠杆菌O11O-抗原基因簇序列的破译及特异分子标识的鉴定[J].微生物学报,2006,46(3):341-346.
[19]程剑松,王威,王荃,等.大肠杆菌O23O-抗原基因簇序列的破译及UDP-N-乙酰葡萄糖-C4异构酶的鉴定[J].微生物学报,2006,46(5):702-708.
[20]孙瑛勋,栾晓,孙启鸿,等.合成多肽为半抗原制备抗SCF受体单克抗体[J].细胞与分子免疫学杂志,1998,14(2):126-129.
猜你喜欢
西北农林科技大学学报(自然科学版)(2019年8期)2019-07-17 02:43:32
网络安全和信息化(2018年4期)2018-11-09 12:01:52
哈尔滨医药(2016年3期)2016-12-01 03:58:34
癌变·畸变·突变(2015年4期)2015-02-27 06:15:17
现代检验医学杂志(2015年1期)2015-02-06 01:59:13
遗传(2015年5期)2015-02-04 03:06:55
海洋科学(2014年12期)2014-12-15 03:35:00
计算机与网络(2013年6期)2013-08-15 00:50:42
食品科学(2013年15期)2013-03-11 18:25:36
中国烟草学报(2012年5期)2012-04-12 06:21:21
