
大鼠空间学习记忆能力测试的时反应量-效关系
2010-09-20 08:20:08汪萌芽
皖南医学院学报 2010年6期
秦 雯,曲 睿,汪萌芽
(皖南医学院 细胞电生理研究室,安徽 芜湖 241002)
时反应量-效关系的概念及其相关数学模型[1,2],通常用于药物作用的分析[3,4],非药物性作用过程的时反应量-效关系少见研究报道。我们实验室前期曾在 Y型迷宫试验中检测了正确反应潜伏期等指标,观察到类双曲线型时反应量-效关系的存在[5]。Morris水迷宫(Morris water maze)是较为理想的经典空间学习记忆能力检测方式[6],主要包括定位航行(place navigation)实验和空间探索(spatial probe)实验两个部分,前者用逃避潜伏期(escape latency)来反映学习能力,后者以通过虚拟平台的次数反映记忆能力,显然都是与时间相关的时反应指标[1],但其是否存在时反应量-效关系尚未见分析报道。另外,学习、记忆功能是相互交叉、并存、关联的神经活动过程,而在重复测试过程中,既有表现为刺激强度的递增规律,又有强化或消退作用的参与,这些学习记忆能力检测指标的时反应量-效关系能否反映这些作用特征,也是值得研究的问题。为此,本文应用 Morris水迷宫测试法进行大鼠空间学习记忆能力的检测,并对各项指标[7~9]进行观察和分析,以寻找具有时反应量-效关系的有效时反应指标。
1 材料和方法
1.1 动物 雄性 SD大鼠 14只,动物均由南京青龙山动物中心提供。实验动物到达实验室适应性饲养 1周后的体重为(257±16)g。
1.2 仪器 Morris水迷宫水池由本实验室参照文献[9]自制,即一直径 120 cm,高 50 cm的不锈钢圆形水池,分 4个象限,配有 2个直径 10 cm、高 30 cm的平台,一个为不锈钢可视平台,另一个为亚克力材料制成的透明不可视平台。摄像设备为 Smart视频追踪系统(San Diego Instuments,USA)中的红外摄像机(悬挂于水迷宫水池的正上方),大鼠行为Morris实验系统由中国科学院心理健康重点实验室隋南实验室馈赠。
1.3 实验方法
1.3.1 实验条件 水迷宫实验在安静、视觉线索充足的房间进行,房间里的一切物品位置保持固定不变(实验时,实验者的位置也固定不变),以作视觉参照系。间接照明,避免光线直射水面。水温控制在 20~25℃,并在实验过程中保持水温恒定。实验前先进行寻找平台训练,正式实验包括连续 7 d的定位航行实验和随后的 1次空间探索实验两个部分。实验于每日上午 9∶00开始进行。
1.3.2 寻找平台训练方法 大鼠在进行正式测试前,先进行游泳以及寻找可视平台的训练,目的是筛除游泳能力缺陷或有视力障碍的大鼠,并使大鼠知道水迷宫池内有一平台,可以作为逃生的目的地。训练时,将不锈钢可视平台置于迷宫水池的中心,台面高出水面 1 cm,每只大鼠先置于平台上适应 20 s,随后训练 3次,随机从不同象限的入水点投入水中,学习寻找平台 60 s,若超过 60 s仍未找到平台,由实验者引导至平台上停留15 s,然后取出擦干放回饲养笼。训练结束后,筛除有缺陷的大鼠。
1.3.3 定位航行实验 将透明平台置于 4个象限中任一象限 1/2弧度,池壁到池中心距离 1/2的位置,水面高于台面1 cm。每日测试一次,含分别从4个象限(顺序随机)面向池壁入水的 4个测试,每个测试间隔 30 s。软件的最长检测时间设为 60 s,驻台时间 5 s(作为找到平台的标准)。若大鼠能找到并登上平台,在平台上停留 5 s后停止记录,将软件记录的大鼠在水中游泳的时间记为逃避潜伏期;若大鼠 60 s内未找到平台,则在软件停止计时后由实验者引导其登上平台并停留 15 s,此时的逃避潜伏期计为 60 s。Morris水迷宫实验软件除记录大鼠游泳的运动轨迹图外,还实时记录运动时间、平台驻留时间、运动距离、入水角度、各象限的活动时间等,并分析记录游泳速度、分析策略等指标。由此将 4个象限入水的 4个检测求出平均结果,记为当天的检测结果。
1.3.4 空间探索实验 第 7天定位航行实验结束1 h后,将平台撤离,大鼠从原平台所在象限的对侧象限入水,测试空间探索实验一次。大鼠行为 Morris实验系统软件除记录大鼠游泳的运动轨迹图外,还分析记录大鼠在 60 s内穿过原平台所在位置的次数。同时观察首次通过虚拟平台时间、游泳速度、靶象限活动时间百分比等指标。
1.4 统计分析 结果以均数±标准差(¯x±s)表示,两组均数差异的比较用 t检验,多组均数差异的比较用方差分析(考虑大鼠个体间差异时用双因素方差分析),若具有显著性意义则用 Newman-Keuls两两对比测验,多组构成比的比较用 χ2检验,P<0.05为差异具有统计学意义。
2 结果
2.1 定位航行实验的指标分析
2.1.1 逃避潜伏期 分析 14只大鼠第 1~7天的数据(图 1 Escape latency,d1~7),其双因素方差分析结果:动物因素 F=1.5879,P=0.1067,表明大鼠间差异没有显著性统计学意义;测试日因素 F=8.5541,P=3.80E-07,表明测试日间差异具有非常显著性意义,即随着测试(或称学习、刺激)次数的增加,逃避潜伏期逐渐缩短,存在时反应量-效关系(图 1),参照文献[1]的分型方法,理论上属于 T型双曲线关系(呈反变关系),但因本实验采用了上限性数据,即有一固定的最长时间 60 s,应归于 P型双曲线关系。测试日因素的日间 Newman-Keuls两两对比测验显示,仅第 1天的逃避潜伏期与第 2~7天的差异有非常显著性统计学意义(P<0.001),提示重复测试(学习)可提高大鼠的空间学习能力,并在第 3天即可接近最好成绩。
2.1.2 靶象限活动时间百分比 分析 14只大鼠第 1~7天的数据(图 1 Time in target quadrant,d1~7),其双因素方差分析结果:动物因素 F=1.4842,P=0.1424,表明大鼠间差异无显著性意义;测试日因素 F=10.1308,P=2.9167E-08,表明测试日间差异具有非常显著性意义,即随着测试次数的增加,靶象限活动时间百分比逐渐增加,存在时反应量-效关系(图 1),可称为反 H型双曲线关系(呈正变关系)[1]。测试日因素的日间 Newman-Keuls两两对比测验显示,第 1天的靶象限活动时间百分比与第 2~7天,第 2天与第 4~7天的差异有非常显著性意义(P<0.01),提示重复测试(学习)可提高大鼠在靶象限的定位航行时间,提高空间学习能力,并在第 3天即可接近最好水平。由此可见,靶象限活动时间百分比的观察结果,与同时检测的逃避潜伏期结果是相互吻合的。
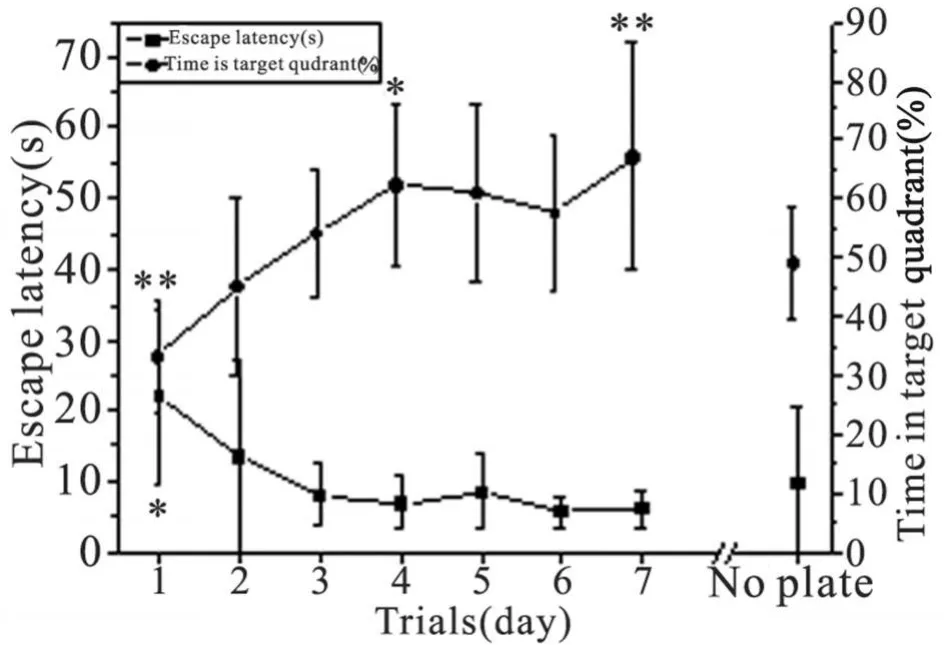
图 1 大鼠Morris水迷宫实验测试日与逃避潜伏期、靶象限活动时间百分比的时反应量-效关系d 1~7:定位航行实验;No plate:空间探索实验;¯x±s,n=14;与 No plate比较的配对 t测验:*P<0.05,**P<0.01Fig 1 Timed dose-response relationship of escape latencys,percentage of swimming time in target quadrant(%)at trial days by place navigation test or spatial probe test d 1~7:place navigation test;No plate:spatial probe test;¯x±s,n=14;Paired t test vs No plate:*P<0.05,**P<0.01
2.1.3 速度分析14只大鼠第 1~7天的数据(图 2,d1~7),其双因素方差分析结果:动物因素F=2.5801,P=0.0051,表明大鼠间差异有非常显著性意义;测试日因素 F=0.7895,P=0.5809,表明测试日间差异没有显著性意义。提示动物的游泳速度在测试的时间内保持相对稳定。
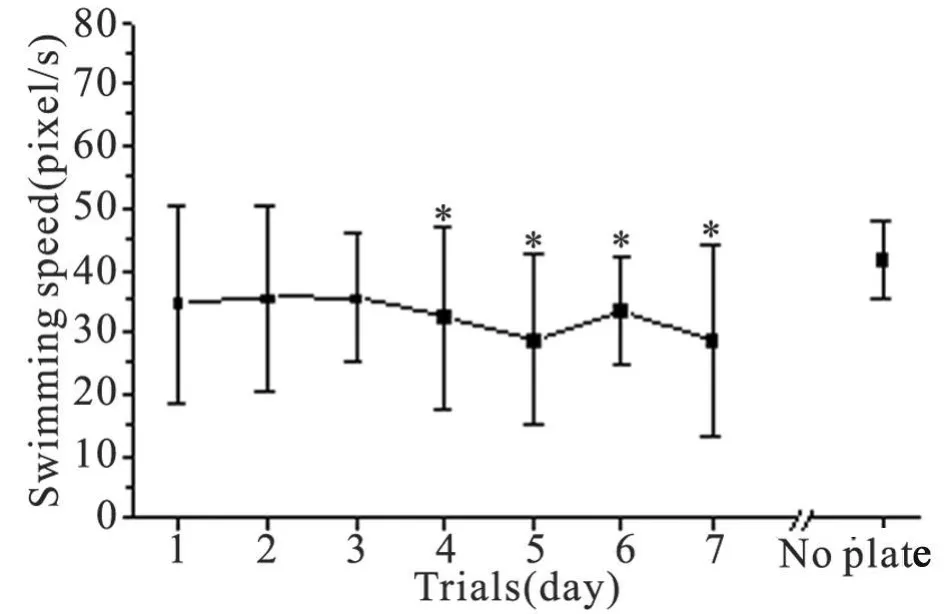
图 2 Morris水迷宫实验中大鼠的游泳速度d 1 ~ d 7:定位航行实验;No plate:空间探索实验;¯x±s,n=14;与No plate比较的配对t测验:*P<0.05Fig 2 Swimming speed of rat in Morris water maze trailsd 1 ~ d 7:place navigation test;No plate:spatial probe test;¯x±s,n=14;Paired t test vs No plate:*P<0.05
2.1.4 分析策略 观察 14只大鼠第 1~7天测试的分析策略构成比,根据软件设定的分析策略为 4种 ,即直线式 (straight)、趋向式(tendency)、边缘式(marginal)和随机式(random),结果见图 3。统计分析结果:χ2=46.83,P=0.0002,表明随着测试次数增加,分析策略构成比差异具有非常显著性统计学意义,即直线式策略逐渐增多,在第 3天即已占优势,到第 6天达 60.71%。趋向式策略仅在第 2天占优势,其后随着直线式策略使用的增多而下降。随机式策略只在第 1天占优势,随着训练增加而逐渐减少;而边缘式搜索策略到第 3 d之后几乎不再使用。
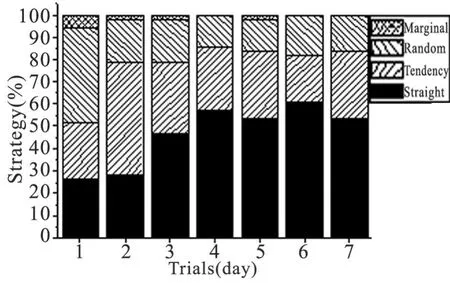
图 3 大鼠定位航行实验中使用的分析策略构成比Fig 3 A percentage of strategies used in place navigation test
2.2 空间探索实验的指标分析 在空间探索实验中,14只大鼠在 60 s内的穿台次数为(5.07±2.62)次,其中首次穿台时间为(9.71±10.87)s(图1 No plate),可视为首次逃避潜伏期,与第 1天的逃避潜伏期(21.95±12.20)s相比显著缩短(配对 t测验,P<0.05),与第 2~7天的逃避潜伏期相比无明显差异,提示空间探索实验的开始,大鼠对平台位置的空间记忆能力大致维持在经多次定位航行训练后的水平。60 s测试时间内,大鼠的靶象限活动时间百分比为(48.81±9.53)%(图 1 No plate),与第 1天和第 7天的靶象限活动时间百分比差异均有非常显著性统计学意义(配对 t测验,P<0.01),且游泳速度(图 2 No plate)为(41.50±6.39)pixel/s,与第7天相比显著性加快(P<0.05)。将探索实验的 60 s等分为两段,进行分析比较(图 4),前段穿台次数明显多于后段(配对 t测验,P<0.05),后段游泳速度显著快于前段(配对 t测验,P<0.01)。
3 讨论
本实验观察到 Morris水迷宫实验的重复定位航行实验,可提高大鼠空间学习能力,随学习天数增加,逃避潜伏期逐渐缩短,在靶象限活动时间的百分比逐渐增加,与报道结果相吻合[7,8]。由于逃避潜伏期是时反应指标[1,2],而重复测试的过程,也就是重复刺激或学习的过程,呈现为刺激强度的累积倍增,可以看作是刺激量的增加,类似于药效学中的药物剂量,其与逃避潜伏期的关系,也就类似于药物的时反应量效关系,经方差分析显示测试日间差异具有显著性意义,表明了时反应量-效关系的存在[2]。至于逃避潜伏期的时反应量-效关系类型,由于时反应指标的特殊性,应当有不确定结果出现,也就是在测试的观察时间内动物不能找到平台,潜伏期趋向于无穷大;而当测试次数增大时,也不是无限缩短,总有一趋向的最短潜伏期存在,理论上属于 T型双曲线关系(呈反变关系)[1,2]。不过,由于本实验采用了上限性数据,即有一固定的最长时间 60 s,应归于 P型双曲线关系(可以看作T型的特例)[1,2]。对于靶象限活动时间的百分比,看起来是一种构成比数据,但其具有活动时间性质,也可看作为类似的时反应指标,其与刺激量的关系为正变关系,但其规律与报道的 H型双曲线关系[1,2]又有所不同,可称为反 H型双曲线关系,其具体定量规律有待于进一步研究。
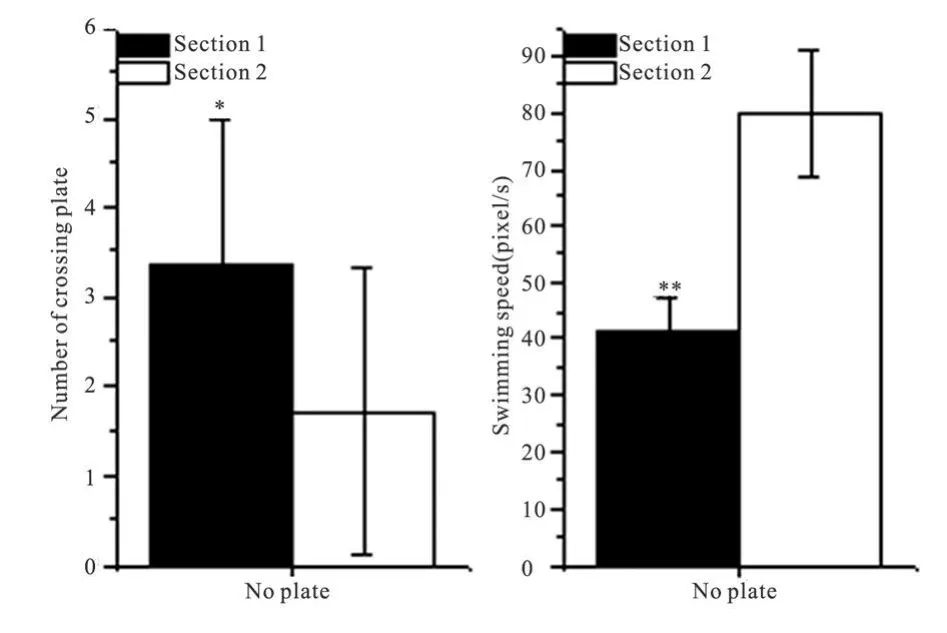
图 4 空间探索实验前后两时段大鼠穿台次数和游泳速度的比较
实验中发现,空间探索实验中的首次登台时间其实等同于定位航行实验中的逃避潜伏期,因此可与之前的逃避潜伏期一起比较,与第 6、7天的逃避潜伏期相近,且与第 1天的潜伏期差异有显著性意义,提示在进行空间探索实验之初,大鼠的空间记忆水平维持在定位航行训练后的水平。然而,观察空间探索实验的靶象限活动时间百分比,却得出与逃避潜伏期不一致的结果,即与定位航行实验中第1天相比有显著性差异(高于第 1天的靶象限活动时间百分比),说明空间探索实验有空间记忆的体现,但同时又显著低于第 7天的靶象限活动时间百分比,说明其探索实验并未能反映定位航行实验学习的结果,鉴于本实验中探索实验与定位航行实验间隔仅 1 h,是否由遗忘因素所致尚不能确定。为此,我们进一步将空间探索实验分为前后两段时间(各 30 s)分别分析穿台次数和游泳速度,结果表明前段的穿台次数显著高于后段,但游泳速度显著低于后段。结合上述空间探索实验的游泳速度明显高于第 7天的结果,可以看出在空间探索实验的初期,大鼠显然是与前期定位航行实验的学习相关的记忆状态进行平台搜寻的,是记忆的体现,而在后期则是不同的状态。可能的解释是,撤去平台后大鼠对原平台的空间记忆在空间探索实验的初期得到表达,但随着不断寻找的失败(平台已撤),进而呈现记忆的消退,并开始尝试重新寻找求生平台,这不仅是靶象限活动时间百分比相比于第 7天定位航行实验时降低的原因,也为上述后一时间段的游泳速度提高所证明。由此可见,空间探索实验作为大鼠空间记忆的检测手段,其本身对记忆能力存在着干扰或消退作用,与报道的重复 Morris水迷宫训练并不改变空间探索实验的穿台次数和靶象限活动时间百分比的结果相吻合[10]。因此,在 Morris水迷宫实验中如何进一步合理地设计空间探索实验的次数、时间,如何进一步有效观察和分析其动态过程,甚至同时进行记忆与记忆消退过程的分析,都值得进一步的研究。
[1] 汪萌芽,孙瑞元.时反应量-效关系及其研究现状[J].中国药理学通报,1987,3(2):111-114.
[2] 汪萌芽,孙瑞元.双曲线型时反应量-效关系的数学模型[J].中国药理学报,1987,8(6):481-486.
[3] 汪萌芽,郑建全,王邦安.药物升降压作用的时反应量-效关系分析[J].中国药理学报,1992,13(6):501-504.
[4] WANG MY,ZHANG FT.Timed dose-response relationship of depressive action of ouabain on toad heart contraction in vitro[J].Acta Pharmacol Sin,1993,14(6):523-526.
[5] 胡凤玉,蒋娟,汪萌芽.迷宫学习模型实验的时反应量-效关系[J].皖南医学院学报,2001,20(3):159-161.
[6] 梅镇彤.学习和记忆的神经生物学[M].第 1版.上海科技教育出版社,1997:61.
[7] 陈罗西,郭玲玲,李亮.Morris圆形水迷宫的应用及其相关检测指标分析[J].辽宁中医药大学学报,2008,10(8):55-56.
[8] 胡镜清,温泽淮,赖世隆.Morris水迷宫检测的记忆属性与方法学初探[J].广州中医药大学学报,2000,17(2):117-119.
[9] VORHEES CV,WILLIAMS MT.Morris water maze:procedures for assessingspatial and related forms of learning and memory[J].Nature Protocols,2006,1(2):848-858.
[10]郑红,王文,刘渝,等.重复 Morris水迷宫训练提高大鼠空间学习能力却不影响空间记忆能力[J].神经解剖学杂志,2007,23(6):599-603.
猜你喜欢
中学生数理化·高一版(2022年3期)2022-04-05 08:17:14
电子制作(2019年24期)2019-02-23 13:22:26
西南交通大学学报(2018年5期)2018-11-08 10:58:04
电子测试(2018年11期)2018-06-26 05:56:04
中学生数理化·七年级数学人教版(2017年4期)2017-07-08 11:48:34
苏州科技大学学报(自然科学版)(2017年1期)2017-03-20 15:25:20
文理导航·科普童话(2016年7期)2017-02-04 20:30:44
知识产权(2016年8期)2016-12-01 07:01:32
文理导航·科普童话(2016年4期)2016-05-31 23:32:38
儿童故事画报·智力大王(2015年12期)2016-01-23 01:14:35
