
当前细菌耐药性的药理分析
2010-09-13 06:03白晓黎
中国医药导报 2010年29期
白晓黎
(河南科技大学第一附属医院,河南洛阳 471003)
细菌对抗生素产生耐药性是医学领域最大的挑战之一。细菌耐药性是细菌产生对抗生素不敏感的现象,产生原因是细菌在自身生存过程中的一种特殊表现形式。现代社会因抗生素滥用,细菌的耐药性越来越强,已引起广大医务工作者的高度重视[1]。现对我院分离得到的256株细菌的耐药情况进行分析,以期为临床提供参考。
1 材料与方法
1.1 材料
选取我院 2009年 10月~2010年 1月分离得到的256株细菌,其中,从患者痰液标本中分离得到45株,尿液中分离得到59株,血液中分离得到82株,大便中分离得到17株,胸腹水和分泌物等其他感染性标本中分离得到53株。
1.2 方法
采用WHO推荐的K-B纸片扩散法测定细菌对药物的敏感性,将含有定量抗菌药物的纸片贴在已接种测试菌的琼脂平板上。纸片中所含的药物吸取琼脂中的水分溶解后不断地向纸片周围区域扩散形成递减的梯度浓度。在纸片周围抑菌浓度范围内测试菌的生长被抑制,从而形成透明的抑菌圈。抑菌圈的大小反映测试菌对测定药物的敏感程度,抑菌圈愈大,最低抑菌浓度(MIC)愈小。
2 结果
本组菌株中,金黄色葡萄球菌35株,链球菌属22株,革兰阳性杆菌8株,大肠埃希菌89株,肺炎克雷伯杆菌23株,阴沟肠杆菌17株,铜绿假单胞菌15株,不动杆菌7株,产气肠杆菌6株,其他菌种34株。可见细菌以革兰阴性杆菌最多,其次是革兰阳性球菌。菌株均有一定程度的耐药,有的已形成多重耐药状况。见表1、2。
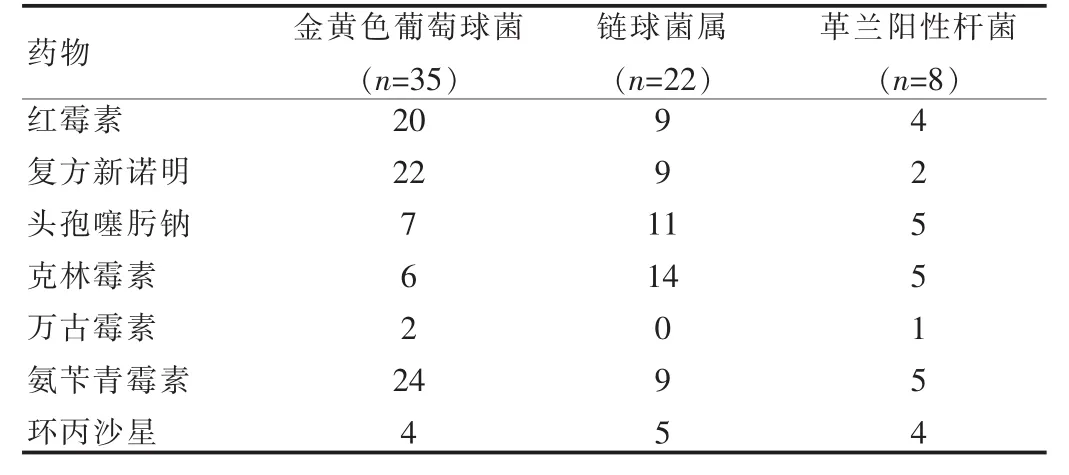
表1 本组革兰阳性菌的耐药菌株数(株)

表2 本组革兰阴性菌的耐药菌株(株)
3 讨论
细菌产生灭活的抗菌药物酶使抗菌药物失活是耐药性产生的最重要机制之一,使抗菌药物作用于细菌之前即被酶破坏而失去抗菌作用[2]。这些灭活酶可由质粒和染色体基因表达。抗菌药物作用靶位改变也是细菌耐药的重要机制,由于改变了细胞内膜上与抗生素结合部位的靶蛋白,降低与抗生素的亲和力,使抗生素不能与其结合,导致抗菌失败。如肺炎链球菌对青霉素的高度耐药就是通过此机制产生的;细菌与抗生素接触之后产生一种新的原来敏感菌没有的靶蛋白,使抗生素不能与新的靶蛋白结合,产生高度耐药[3-4]。靶蛋白数量的增加,即使药物存在时仍有足够量的靶蛋白可以维持细菌的正常功能和形态,导致细菌继续生长、繁殖,从而对抗抗菌药物产生耐药。如肠球菌对β-内酰胺类的耐药性是既产生β-内酰胺酶又增加青霉素结合蛋白的量,同时降低青霉素结合与抗生素的亲和力,形成多重耐药机制。改变细菌外膜通透性也是细菌耐药的机制之一,很多光谱抗菌药都对铜绿假单胞菌无效或作用很弱,主要是抗菌药物不能进入铜绿假单胞菌菌体内,故产生天然耐药[5]。细菌接触抗生素后,可以通过改变通道蛋白性质和数量来降低细菌的膜通透性而产生获得性耐药性。正常情况下细菌外膜的通道蛋白以OmpF和OmpC组成非特异性跨膜通道,允许抗生素等药物分子进入菌体,当细菌多次接触抗生素后,菌株发生突变,产生OmpF蛋白的结构基因失活而发生障碍,引起OmpF通道蛋白丢失,导致β-内酰胺类、喹诺酮类等药物进入菌体内减少。在铜绿假单胞菌还存在特异的OprD蛋白通道,该通道允许亚胺培南通过进入菌体,而当该蛋白通道丢失时,同样产生特异性耐药[6]。另外,某些细菌能将金土菌体的药物泵出体外,这种泵因需能量,故称主动流出系统,由于这种主动流出系统的存在及它对抗菌药物选择性的特点,使大肠埃希菌、金黄色葡萄球菌、铜绿假单胞菌等对四环素、氟喹诺酮类、大环内酯类、氯霉素、β-内酰胺类产生多重耐药。本研究中256株均有一定程度的耐药,有的已形成多重耐药状况。故笔者认为临床工作者应掌握细菌耐药性的相关知识,以指导临床正确用药。
[1]朱佑明,李文桂.细菌耐药机制研究现状[J].重庆医学,2006,35(13):1224-1226.
[2]杨家义,余发春,沈金娥,等.临床标本128株细菌分布分析[J].中国医药导报,2009,6(10):180,183.
[3]郝邯生,郝素云,刘敏,等.2006-2007两年临床分离菌的细菌耐药监测研究[J].武警医学院学报,2009,18(7):625-627.
[4]胡发明.从细菌耐药情况看合理选用抗菌药[J].中国药物应用与监测,2008,5(1):58-60.
[5]何旭瑛.细菌耐药性产生的机制与最新研究进展[J].临床和实验医学杂志,2009,8(11):77.
[6]王玉巧,张跃斌,吴艳梅.细菌多药耐药的分子机制及研究进展[J].中国医师进修杂志,2008,31(15):31.
猜你喜欢
当代水产(2021年8期)2021-11-04
纺织科技进展(2021年5期)2021-07-22
云南医药(2019年3期)2019-07-25
中国生殖健康(2019年12期)2019-01-07
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
中国卫生标准管理(2015年24期)2016-01-14
现代检验医学杂志(2015年1期)2015-02-06
食品工业科技(2014年23期)2014-03-11
世界科学(2013年11期)2013-03-11
