
长爪沙鼠线粒体DNA控制区全序列测定及分析
2010-09-08 08:14李长龙卢领群郭红刚柯贤福戴方伟萨晓婴
中国比较医学杂志 2010年4期
李长龙,卢领群,郭红刚,柯贤福,戴方伟,萨晓婴
(浙江省医学科学院 浙江省实验动物中心,杭州 310013)
研究报告
长爪沙鼠线粒体DNA控制区全序列测定及分析
李长龙,卢领群,郭红刚,柯贤福,戴方伟,萨晓婴
(浙江省医学科学院 浙江省实验动物中心,杭州 310013)
目的对长爪沙鼠线粒体DNA控制区全序列进行测定,并对其进行鉴定及进化分析。方法根据长爪沙鼠已知基因序列设计引物,采用PCR产物测序法,对所得的片段进行测序鉴定。结合已公布啮齿类动物D-loop区序列,分析其碱基组成、遗传距离、并基于最小进化法和UPGMA法构建系统进化树。结果获得长爪沙鼠D-loop区序列,其与家鼠、小家鼠和仓鼠平均同源性为58%;碱基组成分析显示,长爪沙鼠与啮齿类动物有相似的碱基组成和碱基偏离,其A-skew和G-skew分别为0.0047和-0.28。进化分析结果显示,长爪沙鼠与家鼠(0.35)、黑家鼠(0.38)和仓鼠(0.39)具有较近的遗传距离,其分化顺序为跳鼠、蔗鼠、长爪沙鼠、仓鼠、家鼠和小家鼠。结论
本研究获得长爪沙鼠D-loop区全序列,确定了长爪沙鼠与仓鼠、家鼠、小家鼠及其它啮齿动物的进化关系,为长爪沙鼠进化研究、线粒体的结构和功能研究奠定基础。
长爪沙鼠;D-loop区;序列测定;进化分析
线粒体DNA(mitochondrial DNA,mtDNA)是高等动物唯一的核外遗传物质,呈共价闭合的环状双链结构。与核基因组相比,线粒体基因组结构简单、进化速度快(是单拷贝核基因的5~10倍),呈严格的母性遗传,遗传行为相对独立,变异发生的几率相对稳定、无组织特异性、提取方便[1]。由于mtDNA的这些独特遗传特性,已被广泛地用于物种起源与进化、生物分类、及群体遗传结构等方面的研究[2-4]。在进化分析时,研究者往往选择不同的区域进行不同时间尺度的进化分析。线粒体控制区(D-loop region)是mtDNA中进化最快、变化最复杂的区域,常用于种内或种间遗传分化研究[5]。对线粒体D-loop区结构和功能的研究不仅将有助于了解DNA复制、转录的机制和进化规律研究,而且对线粒体结构和功能研究也具有很大意义[6]。
长 爪 沙 鼠 (Mongolian gerbil, Meriones unguiculatus)俗称蒙古沙鼠,属于啮齿目,仓鼠科,沙鼠亚科,沙鼠属,又称长爪沙土鼠、蒙古沙鼠和黄耗子等,野生长爪沙鼠主要分布于我国内蒙古及其毗邻的干旱和半干旱地区[7]。1935年大连卫生所的春日送给日本北里研究所20对长爪沙鼠并开始驯化,后引种到美、英、法等国。日本国家实验动物中心(NIBIO)现已培育成3个长爪沙鼠近交系[8]。我国有两个主要的长爪沙鼠群体,分别保存于浙江省实验动物中心和首都医科大学[7]。长爪沙鼠具有独特的解剖学、生理学和行为学性状,对于一些疾病(如:脑缺血、癫痫、高血脂、寄生虫、细菌、病毒和老年性疾病等)的研究具有极为重要的价值[9-11]。长爪沙鼠被认为是研究脂质代谢良好的模型动物[7,8,12,13],在越来越多的研究所使用。但是,国内外对长爪沙鼠线粒体D-loop区的研究还未见报道。本研究对长爪沙鼠线粒体 D-loop区全序列进行测定和鉴定,并结合已公布啮齿类动物D-loop区序列进行系统进化分析,旨在为长爪沙鼠系统进化关系提供遗传学资料,为全长线粒体测定分析奠定基础。
1 材料和方法
1.1 材料
长爪沙鼠来自浙江省实验动物中心(生产许可证号:SYXK(浙)2008-033;使用许可证号:SYXK (浙)2008-0014),长爪沙鼠解剖采集肝脏样品,-20℃保存。
1.2 总DNA提取
采用酚氯仿抽提法提取基因组DNA,0.7%琼脂糖凝胶电泳检测,-20℃保存。
1.3 引物设计和PCR扩增
分别参照长爪沙鼠 mtDNA序列 AB381901和AJ851249设计上下游引物[14,15],引物由上海生工合成。上游(cyt B 3′端)5’-ATCGGACAAGTCGCT TCAAT-3’,下游(12S 5′端)5’-AGCGATGGCTCG TAGTTCTC-3’。
扩增反应体系总体积为25 μL:模板DNA 100 ng、10×buffer 2.5 μL、MgC12(2.5 mol/L)2 μL、dNTP 2 μL、上下游引物(10 pmol/L)各1 μL、1 U pfuDNA聚合酶,加灭菌纯水补足。扩增条件:94℃预变性5 min,94℃变性30 s,56.5℃复性30 s,72℃延伸1 min,30个循环,72℃延伸7 min。
1.4 序列纯化、测序、拼接
用北京天根凝胶回收试剂盒对 PCR扩增产物进行纯化,纯化后的产物由上海美季公司完成测序工作,Chromas 2.22校对测序图,DNAMAN5.5拼接序列。
1.5 序列的鉴定和分析

图1 PCR扩增长爪沙鼠线粒体D-loop区Fig.1 The results of PCR amplified D-loop region in Mongolian gerbil.
从GenBank下载其他啮齿类动物类mtDNA控制区序列,GenBank登录号分别为:NC_005089家鼠(Mus musculus)、NC_012387小家鼠(Mus musculus castaneus)、NC_006914西欧家鼠(Mus musculus domesticus)、NC_006915日本小鼠(Mus musculus molossinus)、NC_010339东欧家鼠(Mus musculusmusculus)、NC_012389缅鼠(Rattus exulans)、NC_ 012461帅家鼠(Rattus praetor)、NC_012374黑家鼠(Rattus rattus)、NC_011638达氏家鼠 (Rattus tanezumi)、AC_000022褐家鼠(Rattus norvegicus)、NC_013068大仓鼠(Tscherskia triton)、NC_007936中国仓鼠(Cricetulus griseus)、NC_003041台湾田鼠(Microtus kikuchii)、NC_001892胖睡鼠(Glis)、NC_ 005314非洲跳鼠(Jaculus)、NC_009056鳞尾松树(Anomalurus)、NC_000884豚鼠(Cavia porcellus)和NC_002658蔗鼠(Thryonomys swinderianus)等18条全长 mtDNA序列中 D-loop序列进行分析。用DNAMAN5.5进行同源性分析;用 EMBOSS中compseq程序统计碱基含量和计算A和G偏离[16];用MEGA 4.1统计物种间遗传距离,并基于Kimura-Parameter双参数模型,用UPGMA和最小进化法构建分子系统进化树[17]。
2 结果与分析
2.1 D-loop区序列组成
PCR产物电泳结果显示,特异性扩增约1.5 kb的片段(图1)。除去两端不准确的部分,共测得长爪沙鼠mtDNA序列1 677 bp,其中包括D-Loop区全序列、tRNA-Phe、tRNA-Val、tRNA-Thr、tRNA-Pro基因部分序列。序列分析显示,长爪沙鼠线粒体 D-loop区序列全长为997 bp(图2),碱基A、C、G、T的含量分别为32.1%、23.2%、12.9%、31.8%;A +T(63.9%)含量大于G+C(46.1%)含量。A-skew的范围为0.15(松鼠)至-0.034(台湾田鼠); G-skew的范围为-0.17(豚鼠)至 -0.41(睡鼠)。长爪沙鼠的 A-skew和 G-skew分别为 0.0047和-0.28,A和G偏离明显(图3)。
2.2 长爪沙鼠D-loop区序列鉴定
长爪沙鼠与其他啮齿动物的 D-loop序列进行同源性分析,结果显示,家鼠(97%)、小家鼠(83%)和仓鼠(68%)动物均具有较高同源性,长爪沙鼠与上述三种动物的同源性为 58%。鉴于 D-loop是DNA中进化最快的部分,说明该序列为长爪沙鼠线粒体D-loop区(图4)。
2.3 系统进化树的建立及遗传距离计算

图2 长爪沙鼠D-loop区序列Fig.2 The complete sequence of D-loop region in Mongolian gerbil
基于D-loop区全序列,用邻接法和最小进化法构建相关动物系统进化树得到基本一致的拓扑结构(图5,图6)。结果显示,进化树主要有了两个分支,A分支由家鼠、小家鼠、仓鼠、长爪沙鼠、蔗鼠和睡鼠构成;在A分支内,家鼠、小家鼠、仓鼠分别聚合后依次与长爪沙鼠、蔗鼠和跳鼠聚合。家鼠、小家鼠、西欧家鼠、日本小鼠和东欧家鼠集中在小家鼠分支上。缅鼠、帅家鼠、黑家鼠、达氏家鼠和褐家鼠集中在家鼠分支上。大仓鼠、中国仓鼠和台湾田鼠集中在仓鼠分支上。B分支由松鼠、豚鼠和跳鼠聚合而成。结果说明啮齿动物的线粒体控制区序列包含了重要的系统发育信息。枝长表示分歧度,枝上的数值为 1 000次重复抽样检验得到的支持率。选择家鼠、黑家鼠和中国仓鼠分别代表家鼠、小家鼠和仓鼠与长爪沙鼠计算遗传距离。结果显示(表1),长爪沙鼠与黑家鼠遗传距离最近为0.35;与家鼠遗传距离为0.37;与中国仓鼠遗传距离为0.39。结果表明长爪沙鼠与黑家鼠最近的亲缘关系,与家鼠较近的亲缘关系,与豚鼠亲缘关系最远为0.49。
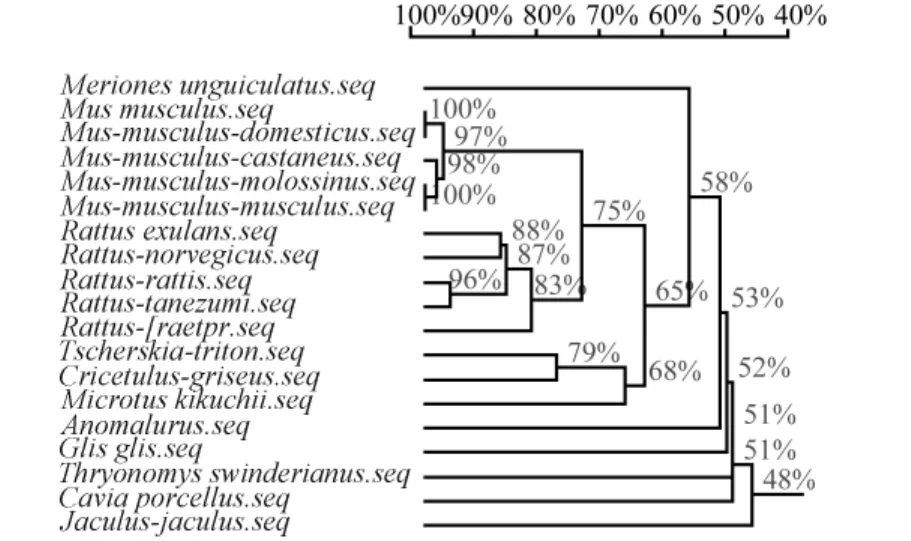
图4 长爪沙鼠与其他啮齿类动物的同源性分析Fig.4 The homology analysis between Mongolian gerbil and other rodents

图5 M-E法构建的D-loop基因系统发生树Fig.5 Phylogenetic tree based on D-loop region sequences generated by minimum-evolution methods.

图6 UPGMA法构建的D-loop基因系统发生树Fig.6 Phylogenetic tree based on D-loop region sequences generated by UPGMA methods.
3 讨论
3.1 长爪沙鼠D-loop区序列的测定和鉴定
mtDNA是闭合的双链环状DNA,D-loop区位于mtDNA复制起点(终点)前,在cyt B和12S基因之间。本研究根据已有的长爪沙鼠线粒体cyt B和12S基因序列分别设计上下游引物,并在线粒体基因组DNA中扩增出约1.5 kb的DNA片段(图1)。经过重复双向测序,结果拼接后获得长1 677 bp的DNA片段。序列中去除转运RNA后获得997 bp序列。为了鉴定该序列,本研究选择18种不同的啮齿类动物线粒体D-loop区序列进行同源性分析,结果显示,长爪沙鼠与家鼠、小家鼠和仓鼠的平均同源性为58%。D-loop区是mtDNA中高度变异的非编码区,进化速率是mtDNA其它区域的3~5倍[18],据此,该序列为长爪沙鼠全长D-loop区序列。
3.2 长爪沙鼠D-loop区序列及进化分析
动物的起源进化长期以来一直困扰着遗传学家。分子遗传学方法为研究群体的起源和进化开辟了新的途径[19]。mtDNA具有很多的特性,利用mtDNA序列构建的系统发生数能很好的反映动物母系的迁徙和进化。mtDNA比核DNA的突变率高5~10倍,其中的 D-loop区是线粒体基因组进化速率最快的区域,进化速率是mtDNA其他区域的3~5倍[18]。许多科学家利用 mtDNA序列差异来研究动物的遗传多样性与起源演化。Giufra对欧洲野猪、欧洲家猪、亚洲野猪和中国梅山猪 mtDNA D-1oop区进行分析,发现家猪有欧洲野猪和亚洲野猪两个母系起源并证实欧洲大白猪是亚洲猪和欧洲猪的杂交种,该结果与18~19世纪初亚洲猪被引入欧洲的历史记载相符[20]。在近年来,动物 mtDNA研究取得了一系列令人瞩目的成果。
不同物种的mtDNA碱基组成和特性存在明显的差异[21]。DNA的鸟嘌呤和胞嘧啶的碱基含量(G +C含量)在不同物种的基因组中波动范围广阔[22,23],甚至在同一物种的基因组不同区域也不相同。这种基因组局部碱基不均衡的现象表现为不同区域的核苷酸碱基组成存在差异[24]。长爪沙鼠 D-loop区序列A、C、G、T的含量分别为32.1%、23.2 %、12.9%、31.8%,其中G的含量显著低于其他碱基的含量,这是mtDNA的一个显著特征[25];A+ T(63.9%)含量大于G+C(46.1%)含量,与其他哺乳动物 A、T含量高,G、C含量低的特点相似[26]。长爪沙鼠的 A-skew和 G-skew分别为0.0047和 -0.28(图3)。与长爪沙鼠遗传距离较近的动物在碱基组成和A和G偏离上较为接近,具有明显的种属特异性。

表1 长爪沙鼠与其他鼠类遗传距离Tab.1 The genetic distance between Mongolian gerbil and other rodents
Chevret等曾利用DNA在溶解温度的方法研究长爪沙鼠与家属和小家鼠的进化关系,结果显示长爪沙鼠与家属和小家鼠存在较相近的进化关系[27]。Colangelo等利用 cyt B和16S rRNA序列研究非洲沙鼠的分子进化关系[28];Chevret等利用 cyt B和12S rRNA基因序列研究沙鼠亚科内分子进化关系[29]。本研究应用MEGA 4.1软件,基于D-loop区全序列,用邻接法和最小进化法构建系统进化树得到基本一致的拓扑结构(图5,6),结果显示,家鼠、小家鼠和仓鼠等动物均分别各自聚合到同一个分支上来,之后依次与长爪沙鼠、蔗鼠和睡鼠聚合在一起。结果表明,长爪沙鼠与家鼠、小家鼠和仓鼠有具有相近的亲缘关系。遗传距离计算结果验证了进化分析结果,长爪沙鼠与褐家鼠最近的亲缘关系,说明家鼠较近的亲缘关系,与豚鼠亲缘关系最远。其分化顺序为跳鼠、蔗鼠、长爪沙鼠、仓鼠、家鼠和小家鼠。以上数据根据动物单一个体的D-loop区序列计算得出,进一步研究应增加每种动物数量,并综合线粒体基因组信息和考古学等其他数据综合分析。
3.3 长爪沙鼠线粒体研究展望
线粒体是细胞的“能量工厂”,多数细胞内90%的氧消耗在线粒体内进行[30]。线粒体序列的测定将为分子进化、能量代谢及相关疾病研究奠定基础。长爪沙鼠是一种正在广泛应用的多功能实验动物。特别是长爪沙鼠的脂质代谢与其它实验动物存在明显差异,但是与人类又有很大的相似之处,是研究人类脂质代谢良好的动物模型[7,8,12,13]。长爪沙鼠线粒体 D-loop区序列的测定和进化分析不但为长爪沙鼠进化研究提供基础数据,而且为获得全长mtDNA序列奠定基础,为长爪沙鼠线粒体的结构和功能研究及广泛应用提供保障。
[1] 徐其放,陈嘉昌,朱世杰,等.动物线粒体DNA的特异结构及应用分子系统学分析的方法[J].中国比较医学杂志,2005,15(5):315-319.
[2] 廖顺尧,鲁成.动物线粒体基因组研究进展[J].生物化学与生物物理进展,2000,27(5):508-512.
[3] 赵兴波,李宁,吴常信.动物线粒体核质基因互作的研究进展[J].遗传,2001,23(1):81-85.
[4] Nishibori M,Shimogiri T,Hayashi T,et al.Molecular evidence for hybridization of species in the genus Gallus except for Gallus varius[J].Animal Genetics,2005,36(5):367-375.
[5] Donne-Goussé C,Laudet V,Hanni C.A molecular phylogeny of anseriformes based on mitochondrial DNA analysis[J].Mol Phylogenet Evol,2002,23(3):339-356.
[6] 孙玉华,王伟,刘思阳,等.中国胭脂鱼mtDNA控制区遗传多样性分析[J].遗传学报,2002,29(9):787-790.
[7] 方喜业,邢瑞昌,贺争鸣.实验动物质量控制[M].北京:中国标准出版社,2008:pp 913-916.
[8] 丁贤明,钱宝珍,Junichiro M,等.长爪沙鼠的遗传多样性分析[J].遗传,2008,30(7):877-884.
[9] Hegsted DM,Gallagher A.Dietary fat and cholesterol and serum cholesterol in the gerbil.J Lipid Res,1967,8(3):210-214.
[10] Tovar-palacio C,Potter SM,Hafermann JC,et al.Intake of soy protein and soy protein extracts influences lipid metabolism and hepatic gene expression in gerbils[J].J Nutr,1998,128(5): 839-842.
[11] Forsythe WA 3rd.Comparison of dietary casein or soy protein effects on plasma lipids and hormone concentrations in the gerbil (Meriones unguiculatus)[J].J Nutr,1986,116(7):1165-1171
[12] Ohkusa T,Okayasu I,Miwa H,et al.Helicobacter pylori infection induces duodenitis and superficial duodenal ulcer in Mongolian gerbils[J].Gut,2003,52(6):797-803.
[13] 钟民涛,黄敏,卢静,等.长爪沙鼠速发型高脂血症模型的初步建立[J].中国实验动物学报,2006,14(3):217-221.
[14] Martin Y,Gerlach G,Schlotterer C,et al.Molecular phylogeny of European muroid rodents based on complete cytochrome b sequences[J].Mol Phylogenet Evol,2000,16(1):37-47.
[15] Chevret P, Dobigny G. Systematicsand evolution ofthe subfamily Gerbillinae(Mammalia,Rodentia,Muridae)[J]. Mol Phylogenet Evol,2005,35(3):674-688.
[16] Salvato P,SimonatoM,BattistiA,etal. Thecomplete mitochondrial genome of the bag-shelter moth Ochrogaster lunifer (Lepidoptera,Notodontidae)[J].BMC Genomics,2008,15 (9):331-346.
[17] Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software,version 4.0[J].Mol Biol Evol.2007,24(8):1596-1599.
[18] Wenink PW,Baker AJ,Tilanus MG.Mitochondrial control region sequences in two shorebird species:the Tumstone and the dunlin,and their utility in population genetic studies[J].Mol Biol Evol,1994,11(1):22-31.
[19] Garrigan D,Hammer MF.Reconstructing human origins in the genomic era[J].Nat Rev Genet,2006,7(9):669-680.
[20] Giuffra E,Kijas J MH,Amarger V,et al.The origin of the domestic pig:independent domestication and subsequent introgression[J].Genetics,2000,154(4):1785-1791.
[21] 高英凯,苗永旺,苏小茜,等.74种鸟类线粒体基因组碱基组成及特征分析[J].云南农业大学学报:自然科学版,2009,24(1):51-58.
[22] Belozersky AN, Soirin AS. A correlation between the compositions of deoxyribonucleic and ribonucleic acids[J]. Nature,1958,182(4628):111-112.
[23] Daubin V,Perrière G.G+C3 structuring along the genome:a common feature in prokaryotes[J].Mol Biol Evol,2003,20 (4):471-483.
[24] Sueoka N.Two aspects of DNA base composition:G+C content and translation-coupled deviation from intra-strand rule of A=T and G=C[J].Molecular Evolution,1999,49(1):49-62.
[25] 樊乃昌,施银柱.中国鼢鼠(Eospalax)亚属分类研究[J].兽类学报,1982,2(2):183-197.
[26] 杨路存,陈桂琛,刘荣堂,等.3种鼢鼠mtDNA控制区结构及其系统发育分析[J].草业科学,2009,26(2):100-106.
[27] Chevret P,Denys C,Jaeger JJ,et al.Molecular evidence that the spiny mouse(Acomys)is more closely related to gerbils (Gerbillinae)than to true mice(Murinae).Proc Natl Acad Sci U S A,1993,90:3433-3436
[28] Colangelo P,Granjon L, Taylor PJ, etal. Evolutionary systematics in African gerbilline rodents of the genus Gerbilliscus:Inference from mitochondrial genes.Mol Phylogenet Evol,2007,42(3):797-806.
[29] Chevret P,Dobigny G. Systematics and evolution ofthe subfamily Gerbillinae(Mammalia,Rodentia,Muridae).Mol Phylogenet Evol,2005,35(3):674-688.
[30] Chance B,Sies H,Boveris A.Hydroperoxide metabolism in mammalian organs[J].Physiol Rev,1979,59(3):527-605.
Determination and Analysis of Mitochondrial DNA D-Loop Region Complete Sequence of Mongolian Gerbil(Meriones unguiculatus)
LI Chang-long,LU Ling-qun,GUO Hong-gang,SA Xiao-ying
(Zhejiang Center of Laboratory Animals,Zhejiang Academy of Medical Sciences,Hangzhou 310013,China)
ObjectiveTo determine the complete sequence of mitochondrial D-loop region of Mongolian gerbil based on the PCR products.The sequences were analyzed for phylogenetic tree,nucleotide composition and genetic distance。MethodsThe primer was designed according to the published partial sequences.The PCR products were sequenced and determined.Combined with the known D-loop region sequence of other rodents,nucleotide composition and genetic distance were analysed,and the phylogenetic tree was constructed by minimum-evolution(ME)methods and UPGMA methods。ResultsThere was a high homology(58%)between the sequences of Mongolian Gerbil and that of rat,mouse and hamster.The nucleotide composition of Mongolian Gerbil was similar with that of other rodents.The A-skew and G-skew were 0.0047 and -0.28,respectively.The results of phylogenetic tree analysis showed a higher genetic relationship with rat(0.35),mouse(0.38)and hamster(0.39).The order of differentiation was cane rat,desert jerboa,Mongolian gerbil,hamster,mouse and rat。Conclusions The mitochondrial D-loop region sequence of Mongolian gerbil has been determined.To our knowledge,it has not been reported in the literature before.In this study,the genetic relationship between Mongolian gerbil and other rodents has been analyzed.The results of this study may be of importance for studies on evolution,structure and function of the mitochondria in Mongolian gerbil.
Mongolian gerbil;D-loop region;Sequence determination;Phylogenetic analysis
R-33
A
1671-7856(2010)04-0040-06
2009-12-17
“十一五”科技支撑重点项目(2009BAI83B02);浙江省实验动物与安全性研究重点实验室(2008F3021)。
李长龙(1976-),男,助理研究员,博士,研究方向:实验动物遗传学,E-mail:li-changlong@126.com。
萨晓婴(1952-),男,研究员,研究方向:实验动物学,E-mail:saxiaoyin@163.com。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
意林·少年版(2021年13期)2021-08-30
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
童话世界(2020年32期)2020-12-25
作文评点报·低幼版(2020年3期)2020-02-12
小学科学(学生版)(2020年1期)2020-01-19
浙江医学(2018年16期)2018-09-08
小学生必读(中年级版)(2018年5期)2018-08-15
第二课堂(小学版)(2017年11期)2017-11-27
