
不同建植密度对松嫩平原肥披碱草无性系种群构件结构的影响
2010-08-08 06:37:06丁雪梅李玉梅付婷婷张晓君孙宏刚沈景林杨允菲
东北农业大学学报 2010年9期
丁雪梅,李玉梅,周 婵,杨 举,付婷婷,张晓君,孙宏刚,沈景林,杨允菲
(1.吉林大学畜牧兽医学院,长春 130062;2.辽宁大学生命科学学院,沈阳 110036;3.吉林大学农学部网络与教育技术中心,长春 130062;4.吉林大学农学部党务办公室,长春 130062;5.吉林省林业勘察设计研究院,长春 130022;6.东北师范大学草地研究所植被生态科学教育部重点实验室,长春 130012)
肥披碱草(Elymus excelsus Turcz.),种别名是高滨麦,是禾本科多年生丛生型草本植物。肥披碱草饲用价值较高,为品质优良牧草,产草量及种子产量高,抗寒、耐旱,抗风沙及一定的耐盐碱能力,对土壤的适应性广,对水肥反应敏感,适宜干旱及半干旱、轻度及中度盐渍化土壤上栽培的优良牧草[1]。
以往对肥披碱草的研究主要集中在肥披碱草抗寒性、抗旱性、耐热性、营养繁殖体的冬眠特性、生殖器官的数量性状以及盐胁迫对肥披碱草种子萌发特性影响等[1-9],对栽培条件下不同密度肥披碱草种群构件结构的研究尚未见报道。本文对不同密度条件下松嫩平原人工建植第2年的肥披碱草无性系分蘖株和分蘖苗构件结构进行统计分析从分蘖株和分蘖苗构件的结构中了解无性系的动态,为构件理论的深入研究提供科学积累。
1 材料与方法
1.1 样地设置方法
研究地点位于松嫩平原南部吉林省长岭县种马场即东北师范大学草地生态定位研究站,地理位置为东经 123°45′~123°47′,北纬 44°40′~44°44′。该地区为松嫩平原南部冲积平原,地势平坦,海拔137.8~144.8 m 之间[1]。
于2005年5月初,在实验田进行2种建植方式,即实生苗和分蘖苗建植肥披碱草种群。实验田地势平坦、土壤为风沙土,经机械和人工精细整地后,规划成面积为2 m×6 m的实验小区,3次重复,定时除杂草和进行防虫害处理。
1.1.1 实生苗建植
实生苗建植5个密度:16、25、45、100、400株·m-2,即实生苗与实生苗之间的距离分别为25、20、15、10、5 cm,采取穴播,每穴播5粒,覆盖草帘以防晒、保温、保湿,隔1天灌溉1次,出苗率为100%,在分蘖之前间苗为单株。
1.1.2 分蘖苗建植
分蘖苗建植2个密度:9和45株·m-2,即分蘖苗与分蘖苗之间的距离分别为30和15 cm,从天然种群中选取肥披碱草分蘖丛,将其分成单一的分蘖苗栽植至实验小区中,每天灌溉1次,直至成活。
1.2 取样方法
2006年9 月下旬,于肥披碱草生长季末期,对实生苗建植的5个密度和分蘖苗建植的2个密度肥披碱草,随机挖取25个分蘖丛,按营养株、生殖株和冬性苗从分蘖节处掰下分置,分别记数3种构件数量,并于80℃下烘至恒重后称重。
1.3 数据分析方法
用 SPSS统计软件(Version 13.0,SPSS Inc.,Chicago,Illinois),计算不同密度条件下种群营养分蘖株、生殖分蘖株和分蘖株的数量和生物量的平均值(M)和标准误(SE),其中,平均值(M)代表种群平均水平数量指标量度,标准误(SE)和变异系数(CV)用以反映其变异度。对绝对数量指标,即不同密度条件下种群营养株、生殖株和冬性苗的数量和生物量、生殖分配等数量指标,以及对相对数量指标,即对同一密度营养株、生殖株和冬性苗的数量和生物3种构件所占的比率进行单因素方差分析,用Duncan新复极差进行多重比较。生殖分配指标(RAⅠ、RAⅡ、RAⅢ)的计算公式如下:
RAⅠ=(生殖分蘖株生物量/地上部分总生物量)×100%
RAⅡ=(花序生物量/地上部分总生物量)×100%
RAⅢ=(成熟饱满籽粒生物量/地上部分总生物量)×100%
2 结果与分析
2.1 肥披碱草无性系种群构件数量结构
肥披碱草种群地上部分构件可被分为生殖分蘖株、营养分蘖株和冬性苗。建植后,经过2个生长季的生长和营养繁殖,各初始密度无性系种群构件数量结构见表1。

表1 肥披碱草无性系种群构件数量结构Table 1 Number structure of modules of Elymus excelsus clone population
两种建植方式的种群生殖分蘖株、营养分蘖株、冬性苗以及地上部分构件总数量均随密度升高而呈下降的趋势(P<0.05)。分蘖苗建植的 9株·m-2种群,生殖分蘖株数量皆高于实生苗建植的5个密度种群(P<0.05);营养分蘖株和冬性苗数量与实生苗建植的 16,25 株·m-2种群无显著差异(P>0.05),皆高于实生苗建植的其他3个密度的种群(P<0.05);地上部分构件总数量与实生苗建植的16株·m-2种群无显著差异(P>0.05),皆高于实生苗建植的其他4个密度的种群(P<0.05)。分蘖苗建植的45株·m-2种群,生殖分蘖株数量高于相同密度的实生苗建植的种群(P<0.05),而营养分蘖株、冬性苗以及地上部分构件总数量皆低于相同密度的实生苗建植的种群(P<0.05)。
3种构件中,两种建植方式不同密度的无性系种群皆以冬性苗数量所占比率最大,在61.5%与92.8%之间。从生殖株与营养株数量二者所占比率来看,分蘖苗建植的2个密度无性系种群的生殖分蘖株高于营养分蘖株数量所占比率,而实生苗建植的3个低密度种群营养分蘖株高于生殖分蘖株所占比率,而2个高密度种群二者所占比率无显著差异。
2.2 肥披碱草无性系种群构件生物量结构无性系种群构件生物量结构见表2。
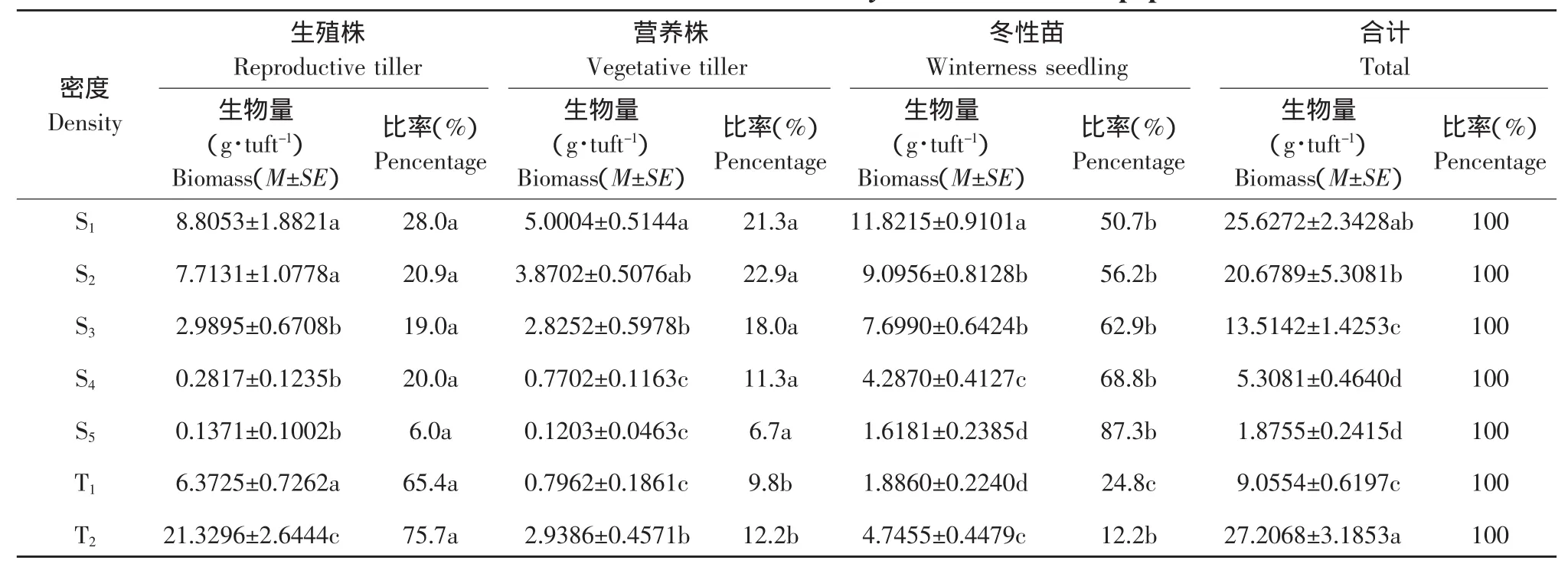
表2 肥披碱草无性系种群构件生物量结构Table 2 Biomass structure of modules of Elymus excelsus clone population
两种建植方式的肥披碱草种群生殖分蘖株、营养分蘖株、冬性苗以及地上部分构件总生物量均随密度升高而呈下降的趋势(P<0.05)。分蘖苗建植的9株·m-2种群,生殖分蘖株生物量皆高于实生苗建植的5个密度种群(P<0.05);营养株生物量低于实生苗建植的16株·m-2(P<0.05),高于2个高密度种群(P<0.05),与 25,45 株·m-2无显著差异(P>0.05);冬性苗生物量与 100 株·m-2无显著差异(P>0.05),高于 400 株·m-2种群(P<0.05),低于实生苗建植的3个低密度种群(P<0.05);地上部分构件总生物量与实生苗建植的16株·m-2无显著差异(P>0.05),高于实生苗建植的其他4个密度种群(P<0.05)。而分蘖苗建植的45株·m-2种群,生殖分蘖株数量高于相同密度的实生苗建植的种群(P<0.05),而营养分蘖株、冬性苗皆低于相同密度的实生苗建植的种群(P<0.05),但二者地上部分构件总生物量无显著差异(P>0.05)。
分蘖苗建植的2个密度无性系种群皆以生殖株生物量所占比率最大,皆高于60%,而实生苗建植的5个密度种群皆以冬性苗生物量所占比率最大,在50.7%与87.3%之间。从生殖株与营养株数量所占比率来看,分蘖苗建植的2个密度种群的生殖分蘖株高于营养分蘖株所占比率,而实生苗建植的5个密度种群二者所占比率无显著差异。
2.3 肥披碱草无性系生殖分配
结果见图1。
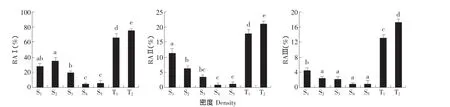
图1 肥披碱草的生殖分配Fig.1 Reproductive allocation of Elymus excelsus clones
生殖分蘖株由花序和茎组成。生殖分蘖株生物量与地上部分构件总生物量的比值(RAⅠ)、花序生物量与地上部分构件总生物量的比值(RAⅡ)以及成熟饱满子粒生物量与地上部分构件总生物量的比值(RAⅢ)是衡量生殖分配的重要指标。不同密度肥披碱草RAⅠ、RAⅡ、RAⅢ见图1。两种建植方式的肥披碱草RAⅠ、RAⅡ和RAⅢ均随密度升高而呈下降的趋势(P<0.05),分蘖苗建植的2个密度皆高于实生苗建植的5个密度的RAⅠ、RAⅡ和 RAⅢ(P<0.05)。
3 讨论与结论
栽培条件下的肥披碱草种群有两种建植方式:实生苗和分蘖苗建植。本试验中两种建植方式的肥披碱草种群在第二年的生长季节末期,生殖分蘖株、营养分蘖株、冬性苗数量和生物量,RAⅠ(生殖分蘖株生物量与地上部分总生物量的比值)、RAⅡ(花序生物量与地上部分总生物量的比值)、RAⅢ(成熟饱满子粒生物量与地上部分总生物量的比值),以及地上部分构件总数量和生物量皆呈现共同的规律,即随密度升高而呈下降的趋势,密度对肥披碱草种群生长和有性生殖分配产生影响。当环境资源一定时,在低密度条件下,种群个体数量相对比较少,对环境资源的利用比较充分,种群内不存在竞争。但当密度达到一定高度时,种群内个体为维持个体正常生长,必然导致对资源的竞争。因此密度对种群生长和有性生殖均产生显著影响。
生活史理论认为生殖与营养生长或存活之间存在负耦联关系。关于植物的生殖分配模式是否随密度而发生变化的问题,以往的研究结果有三种情况[10-11]:第一种是高密度下将更多的资源分配给繁殖体;第二是高密度下将更多的资源分配给营养体;第三种是分配到繁殖体和营养体的资源比例不受密度的影响,即生殖分配为定值。垂穗披碱草属于第三种情况,生殖分配比例为一定值,竞争对垂穗披碱草繁殖分配的影响不大,其繁殖体大小随密度的可塑性变化是由于个体总生物量随密度的可塑性变化所致[10]。而肥披碱草种群属于第二种情况,与高密度相比,低密度肥披碱草种群将更多的资源分配给生殖分蘖株、花序和成熟饱满籽粒。可见,即使同为丛生型禾草,不同种类种群的生殖分配规律也不尽相同。在低密度的条件下,肥披碱草种群空间充足,个体之间对资源利用不存在竞争,个体可以在正常生存的条件下正常进行生殖;但在高密度条件下,种群间个体对空间和资源均产生竞争,个体必然把更多能量投入到维持自身正常生存上,相当于生殖自然投入的比例减少。
3个生殖分配指标RAⅠ、RAⅡ、RAⅢ代表了不同水平上生殖分配,其中,RAⅠ为种群水平,即种群中投入到生殖分配的生物量比例,在这个生殖投入的基础上,个体在投入一定比例生物量到有性生殖的器官(RAⅡ),而投入到有性生殖器官的能量不可能完全被用与生殖,其中只有只有一少部分被真正用于籽粒的成熟,即RAⅢ,这在一定程度上体现了个体的繁殖成效。
大多数多年生植物既可通过种子进行有性繁殖,又可通过根、茎、根茎、芽等进行营养繁殖。当生态或遗传条件使其中一种繁殖方式受到限制时,两种繁殖方式所产生的后代数量以及两种类型后代的更新在同一物种内通常会有很大变化[12]。不同建植方式对肥披碱草种群生长和繁殖也产生影响。在第二年的生长季节末期,分蘖苗建植的肥披碱草种群皆倾向于有性繁殖,而实生苗建植的肥披碱草种群则倾向于营养繁殖。分蘖苗建植的2个密度无性系种群皆以生殖株生物量所占比率最大,皆高于60%,而实生苗建植的5个密度种群生殖株生物量所占比率皆低于30%。分蘖苗建植的9株·m-2种群,生殖分蘖株数量和生物量皆高于实生苗建植的5个密度种群(P<0.05)。同为密度 45株·m-2肥披碱草种群,分蘖苗建植的肥披碱草种群生殖分蘖株数量和生物量、RAⅠ、RAⅡ、RAⅢ皆高于实生苗建植的肥披碱草种群(P<0.05);而营养分蘖株、冬性苗数量和生物量,地上部分构件总数量皆小于实生苗建植的肥披碱草种群(P<0.05)。但两种建植方式的种群地上部分构件总生物量之间没有显著差异(P>0.05)。分蘖苗建植的2个密度生殖株数量和生物量所占比率皆高于实生苗建植的5个密度肥披碱草种群(P<0.05)。对于两种建植方式造成的繁殖方式侧重的不同以及其生长规律的可变性及所蕴涵的无性系调节机理与生物生态学意义有待于进一步研究。
[1] 郑慧莹,李建东.松嫩平原的草地植被及其利用保护[M].北京:科学出版社,1993:59-67.
[2] 周德宝.不同披碱草抗寒性的研究[J].阴山学报:自然科学版,1996,13(3):36-39.
[3] 董瑞英,张冬梅,张丽娟,等.利用根冠淀粉水解法鉴定五种牧草的抗旱性及耐热性[J].哲里木畜牧学院学报,1996,6(4):24-27.
[4] 苏慧,刘玉平,张继星,等.Na2CO3胁迫对牧草苗期生长的影响[J].中国草地,2005,27(1):22-25.
[5] 苏慧,沈秀瑛,程伟燕,等.Na2CO3胁迫对牧草苗期生理特性的影响[J].草业科学,2005,22(6):46-48.
[6] 苏慧,尉红梅,马岩,等.Na2CO3胁迫对牧草种子萌发特性影响的研究[J].内蒙古民族大学学报:自然科学版,2005,20(2):168-171.
[7] 杨允菲,祝玲.松嫩平原十五种多年生禾草种群营养繁殖体冬眠特性的介绍[J].草业学报,1994,3(2):26-31.
[8] 杨允菲.东北四种野生披碱草结实器官和种子产量性状的比较[J].中国草地,1990(2):66-68.
[9] 郑红梅,杨允菲.松嫩平原天然肥披碱草种群生殖器官的数量性状分析[J].草原与草坪,2006(4):21-24.
[10] 杜国祯,王刚.垂穗披碱草个体大小与种群密度的关系[J].植物生态学报,1998,22(6):552-558.
[11] Weiner J.The influence of competition on plant reproduction[M]//:Doust L L.Plant reproductive ecology:patterns and strategies.Oxford:Oxford University Press,1988:228-245.
[12] 张玉芬,张大勇.克隆植物的无性与有性繁殖对策[J].植物生态学报,2006,30(1):174-183.
猜你喜欢
甘肃畜牧兽医(2023年6期)2023-05-11 20:51:26
美文(2022年18期)2022-10-20 23:07:14
农业科技通讯(2022年1期)2022-01-17 06:06:40
现代畜牧科技(2021年7期)2021-07-28 06:40:30
山东林业科技(2019年2期)2019-06-03 10:10:54
——以新源县为例
新疆农业科技(2018年5期)2018-05-07 02:25:08
农家科技中旬版(2018年2期)2018-05-05 10:09:54
现代农业(2018年7期)2018-02-18 10:58:09
中国糖料(2016年1期)2016-12-01 06:48:57
新农村(2015年10期)2015-09-28 00:37:29
