
小兴安岭天然白桦林植物物种多样性的多尺度分析1)
2010-08-08 07:22:52赵丽娜孙广玉尹鹏达周志强
东北林业大学学报 2010年6期
赵丽娜 孙广玉 尹鹏达 刘 彤 周志强 王 恒
(东北林业大学,哈尔滨,150040) (白城农业广播电视学校)
黑龙江小兴安岭的地带性植被是阔叶红松(Pinus koraiensis)混交林[1],但由于长时间过度采伐利用等原因,原始林资源消耗殆尽,各种类型次生林成为小兴安岭的主要林型,其中次生白桦(Betula platyphylla)林是主要天然次生林类型之一[2]。小兴安岭实施森林分类经营以来,大部分林型被划入生态公益林,作为商品林的白桦林成为小兴安岭地区经济输出的主要来源之一[3]。因此,维持白桦林及整个生态系统的平衡、稳定,提高白桦林内生物多样性,对整个区域经济发展、生态建设至关重要。目前,对不同林型和立地条件下植物物种多样性的研究很多[4-6],但对于天然白桦林下植物多样性的研究较少。本文应用多样性加法分配法则[7],对小兴安岭带岭林业实验局20年生天然白桦林下的植物物种多样性进行了多尺度分析。探讨天然次生白桦林物种多样性在多尺度内和多尺度间的分配关系,以期为今后合理经营天然白桦林,充分发挥该类型林分的生态功能和经济效益提供一定的科学依据。
1 研究区概况
研究地点位于小兴安岭带岭林业实验局东方红林场(E129°04′42″~129°07′15″,N46°49′21″~46°53′51″),属北温带大陆季风气候区。四季分明,冬季严寒、干燥而漫长,夏季温热而短暂,年平均气温1.4℃左右。地貌以低山为主,多为缓坡,海拔在400~800 m。年降水量550~670 mm,降雨集中在6—8月份;全年无霜期90~120 d。土壤为暗棕壤(色)森林土,植被(植物区系)以长白植物区系的物种为主。研究地点的地带性植被为阔叶红松林,由于长期的采伐利用,目前该区域森林植被以天然次生林和针阔叶混交林为主,其中天然白桦林是研究地点的主要森林类型。在白桦林中常伴生有少量水曲柳(Fraxinus mandschurica)、黄檗(Phellodendron amurense)、胡桃楸(Juglans mandshurica)、大青杨(Populus ussuriensis)等树种;林下灌木层(包括层间藤本植物和乔木幼苗)约有20科37属55种,草本层约有29科62属83种。
2 研究方法
2.1 样地设计和群落调查
2009年6月,在小兴安岭带岭林业实验局东方红林场选取了样方、林分郁闭度、林分坡向3个尺度进行物种多样性的研究。设置等级取样样地(见图1),即有阴坡(北)、半阴坡(西北)、阳坡(南)3个坡向,每个坡向含5个郁闭度(0.4、0.5、0.6、0.7、0.8)样地,每个郁闭度连续调查3 个25 m×20 m的植被样地,这就形成了3个坡向、5个郁闭度、45个植被样地组成的3尺度等级取样系统。对于每个25 m×20 m的植被样地,采取分层取样的方式进行群落调查,在每个25 m×20 m的植被样地的4角和中间设置5个5 m×5 m的灌木样方,灌木样方记录样方内每个物种的高度、盖度、多度(个体数);在每个灌木小样方内随机设置3个1 m×1 m的草本样方,草本样方记录每个物种的高度、相对盖度和多度。
2.2 数据分析
根据取样设计(图1),按照物种多样性加法法则计算框架[7-9],群落内多样性(α)、群落间多样性(β)、区域多样性(γ)在样方、郁闭度、坡向各尺度之内和之间的分配关系为γ=α1+β1+β2+β3(图2)。在 EXCEL中计算灌木和草本的重要值Pi(灌木Pi=(相对优势度+相对盖度+相对多度)/3,草本Pi=(相对高度+相对盖度+相对多度)/3),利用PARTITION软件通过物种加法分配法可以计算出 α(α1,α2,α3)、β(β1,β2,β3)和γ多样性的物种丰富度指数(R)、Simpson(D)指数和Shannon-Wiener(H)指数。
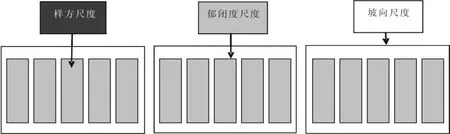
图1 天然白桦林下植物物种多样性的等级尺度取样设计
多样性指数的计算公式详见文献[10]、[11];
物种丰富度Patrick指数:R=S;
Simpson 指数:D=1-∑Pi×Pi;
Shannon-Wiener指数:H=-∑Pi×lnPi。
式中:S为第i物种所在样方的物种总数,Pi为样本中i种的重要值。
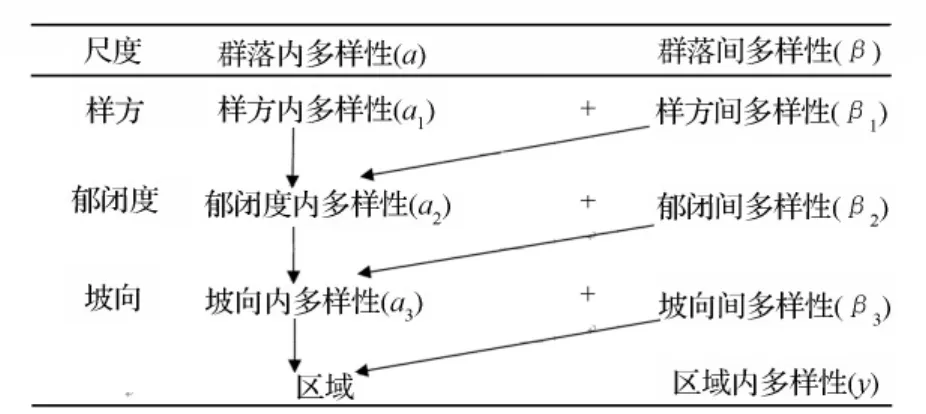
图2 按照物种多样性加法分配法则划分的不同尺度物种多样性组分的关系[6]
3 结果与分析
3.1 天然白桦林下植物多样性在各尺度内的分配关系
3.1.1 不同郁闭度白桦林下灌木层、草本层的植物多样性差异
根据45个植被样地不同郁闭度林下植物物种多样性分析可知,不同郁闭度对林下灌木层和草本层植物物种多样性影响较大(表1)。以物种丰富度为度量指标时,灌木层在0.7郁闭度林分下物种丰富度最高,0.7郁闭度林分下约有34个物种,比其它4个郁闭度多5~11个物种;草本层的物种丰富度也在0.7郁闭度林分下达到最高,0.7郁闭度林分下约有53个物种,比其它4个郁闭度多4~17个物种。这可能与0.7郁闭度林分下土壤中的含水量、水解N和速效K含量相对较高(表2)有关。随郁闭度的增加,灌木层和草本层物种丰富度表现为先增大后减小,这种变化与林下的光照强度、土壤中的含水量、土壤中的养分含量、植物本身的生理特征等综合因素有关。
物种丰富度仅考虑物种的存在与否,而Simpson指数和Shannon指数综合考虑了物种数目和各物种的多度。灌木层的Simpson指数和Shannon指数与物种丰富度呈相同的变化(表1),0.5郁闭度林分下的Simpson指数最高,这是由于0.5郁闭下的常见种多物种分布均匀所致。0.7郁闭度林分下Shannon指数最高,因为大量偶见种的增加使得0.7郁闭度林分的Shannon指数偏高。
草本层的Simpson指数和Shannon指数除0.7郁闭度外,随郁闭度的增大表现为降低的趋势,0.4郁闭度Simpson指数和Shannon指数最高。草本植物的物种数目虽然逐渐增多,露珠草(Circaea cordata)、北重楼(Paris verticillata)、林生茜草(Rubia sylvatica)、烟管蓟(Cirsium pendulum)等一些偶见种数目少,使得群落内物种分布不均匀,所以 Simpson指数和Shannon指数表现为随草本层偶见种的增多而逐渐降低。由于0.7郁闭度物种数目多且分布均匀,所以Simpson指数和Shannon指数相对其它郁闭度较高。
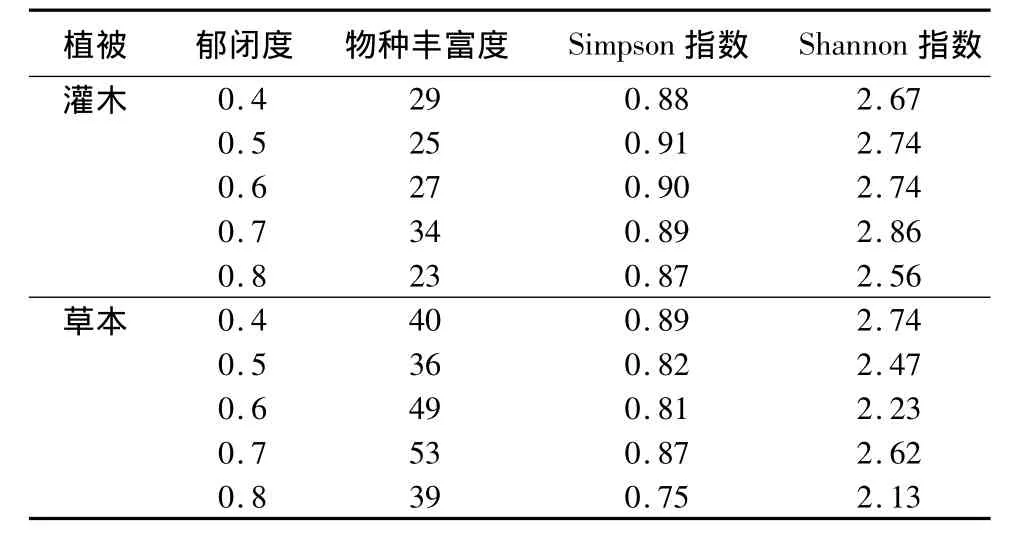
表1 不同郁闭度林下植被多样性

表2 不同郁闭度林下土壤营养元素质量分数
3.1.2 不同坡向白桦林下灌木层、草本层的植物多样性差异
从表3可以看出,不同坡向林下植物物种丰富度差异较大。灌木层在半阴坡物种丰富度为最高,阴坡物种丰富度最低,阴坡比半阴坡少约15个物种。草本层在阳坡物种丰富度为最高,阴坡物种丰富度最低,阴坡比阳坡少约27个物种。由于阴坡林下光照强度较弱,限制了灌木层和草本层植物的生长发育,所以物种丰富度低;而半阴坡和阳坡林下光照相对充足,适宜灌木层和草本层植物的生长发育,所以物种丰富度高。通过(表4)可知,半阴坡有效P质量分数最高,阳坡有机质质量分数最高,这可能是半阴坡和阳坡灌木层和草本层物种丰富度高的另一原因。而水解N在阴坡较之其它2个坡向质量分数高,可能是抑制灌木和草本植物发育的因素。
灌木层和草本层的Simpson指数和Shannon指数在半阴坡最大。这是由于半阴坡物种丰富度高且物种数目分配均匀所致。
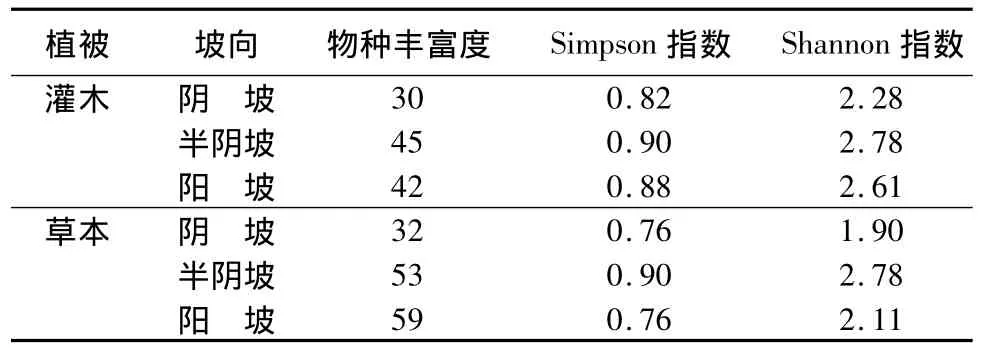
表3 不同坡向林下植被多样性

表4 不同坡向林下土壤营养元素质量分数
3.2 天然白桦林下植物多样性在各尺度间的分配关系
从表5可以看出,当以物种丰富度为植物多样性度量指标时,灌木层和草本层物种丰富度在区域内的最大贡献来自坡向间多样性(β3)。从植物组成的角度分析,灌木层和草本层常见种,如暴马丁香(Syringa reticulata var.amurensis)、东北山梅花(Philadelphus schrenkii)、珍珠梅(Sorbaria sorbifolia)、白花碎米荠(Cardamine leucantha)、毛缘苔草(Carex campylorhina)、狭叶荨麻(Urtica angustifolia)等,种类少但个体数量多;而灌木层和草本层偶见种,如山刺梅(Rosa davurica)、暖木条荚蒾(Viburnum burejaeticum)、草芍药(Paeonia obovata)、落新妇(Astilbe chinensis)等,种类多但在样方内的个体数量少。导致样方内部多样性的均值(α)小于样方内缺失物种的均值(β)。因而,物种丰富度的加法分配法则产生的最大贡献会出自β多样性组分。在坡向间、郁闭度间(β2)、样方间(β1)对区域内多样性(γ)的贡献大小的顺序为:β3>β2>β1。从环境异质性的角度分析,在样方尺度会有微地形和局域小气候的差异;而郁闭度尺度则会出现光照强度、土壤水分和土壤养分含量的差异;坡向尺度又会出现光照时间等大尺度环境因子的差异。因而尺度越大环境异质性越高,β多样性也越大。
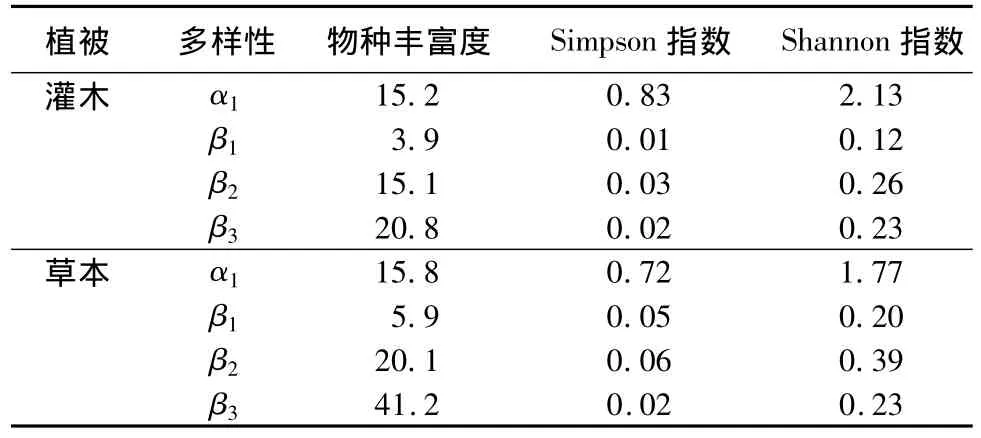
表5 灌木物种,草本物种在样方,郁闭度,和坡向3个尺度上α和β多样性组分构成的物种丰富度,Simpson指数和Shannon指数
以Simpson和Shannon指数为度量指标时,灌木层和草本层对区域多样性(γ)的最大贡献则来自样方内多样性(α1),这是由于灌木层和草本层的一些种类,如暴马丁香、珍珠梅、白花碎米荠、毛缘苔草等,其局域的优势度和均匀度给样方尺度的指数值提供了更多的信息。而由于扩散作用,使群落组成趋于同质,导致β多样性减小。灌木层和草本层的Simpson和Shannon指数在β多样性的水平上,对白桦林区域多样性(γ)最大贡献来自β2,其原因是不同郁闭度间,白桦林下灌木层和草本层的物种种类和盖度的综合差异比坡向间物种种类和盖度的综合差异大所致。
4 结论与讨论
在郁闭度尺度内,0.7郁闭度林分下灌木层和草本层的物种多样性均相对最高,是天然白桦林保护植物多样性适宜密度。
在坡向尺度内,半阴坡林分下灌木层和草本层的物种多样性相对最高,小兴安岭天然白桦林下灌草层的物种多样性与林分所处的坡向有关。
在样方-郁闭度-坡向3个尺度间,当以物种丰富度为植物多样性度量指标时,灌木层和草本层物种丰富度在区域内的最大贡献来自坡向间多样性(β3),说明坡向尺度对物种丰富度的格局起决定作用,是维持小兴安岭天然白桦林物种多样性的有效尺度。各尺度间β多样性组分大小的顺序为:坡向间(β3)>郁闭度间(β2)>样方间(β1)。以 Simpson和 Shannon指数为植物多样性度量指标时,对天然白桦林区域多样性(γ)的最大贡献来自灌木层和草本层样方内多样性(α1),郁闭度间的多样性(β2)在β多样性组分中为最大。这种差异主要是由于后两种指数不仅考虑了物种的存在与否,也考虑了其在样方内的多度。
本文根据野外样地调查结果,开展了天然白桦林生物多样性多尺度分析的尝试,但对尺度内和尺度间生物多样性差异的生物学解释及其环境因子诱因,还有待于在后续研究中补充和加强。
[1] 闫海冰,郝占庆,杨秀清,等.黑龙江省红松林资源动态变化研究[J].生态学杂志,2005,24(09):985-988.
[2] 李俊清,李景文.中国东北小兴安岭阔叶红松林更新及其恢复研究(英文)[J].生态学报,2003,23(07):1268-1277.
[3] 郝占庆,陶大立,赵士洞.长白山北坡阔叶红松林及其次生白桦林高等植物物种多样性比较[J].应用生态学报,1994,5(1):16-23.
[4] 周志强,魏晓雪,刘彤.新疆奇台荒漠植物群落的数量分类及土壤环境解释[J].生物多样性,2007,15(3):264-270.
[5] 刘松春,牟长城,屈红军.不同抚育强度对"栽针保阔"红松林植物多样性的影响[J].东北林业大学学报,2008,36(11):32-35.
[6] 张育新,马克明,祁建,等.北京东灵山辽东栎林植物物种多样性的多尺度分析[J].生态学报,2009,29(05):2179-2185.
[7] Chiarucci A,Bacaro G,Rocchini D.Quantifying plant species diversity in a Natura 2000 network:Old ideas and new proposals[J].Biological Conservation,2008,141:2608-2618.
[8] Gering J C,Crist T O,Veech J A.Additive partitioning of species diversity across multiple spatial scales:implications for regional conservation of biodiversity[J].Conservation Biology,2003,17:488-499.
[9] Chandy S,Gibson D J,Robertson P A.Additive partitioning of diversity across hierarchical spatial scales in a forested landscape[J].Journal of Applied Ecology,2006,43:792-801.
[10] 牛丽丽,杨晓晖.四合木群丛分布区的植物物种多样性研究[J].水土保持研究,2007,14(05):62-66.
[11] 夏鑫,范海兰,洪伟,等.南方红豆杉群落物种的多样性[J].东北林业大学学报,2007,35(11):23-26.
猜你喜欢
南方农业(2022年23期)2022-02-14 09:45:26
思维与智慧·下半月(2021年7期)2021-08-13 16:55:24
草原歌声(2021年4期)2021-06-10 02:25:48
福建林业(2020年5期)2020-03-18 08:23:02
文苑(2019年22期)2019-12-07 05:29:12
防护林科技(2019年7期)2019-08-19 00:50:58
热带林业(2019年4期)2019-03-05 09:53:58
北极光(2018年5期)2018-11-19 02:23:58
现代农村科技(2018年11期)2018-11-15 11:27:58
森林工程(2018年3期)2018-06-26 03:40:46
