
肃北地区稀有植物裸果木种群的空间分布格局
2010-06-08 07:52柴永青曹致中蔡卓山万丽霞李学玲
草业学报 2010年5期
柴永青,曹致中,蔡卓山,万丽霞,李学玲
(1.甘肃农业大学草业学院,甘肃 兰州730070;2.甘肃省肃北县草原站,甘肃 肃北736300;3.西北师范大学生命科学院,甘肃 兰州730070;4.兰州大学草地农业科技学院,甘肃 兰州730020)
植物种群分布格局是关于种群在水平空间上的配置状况或分布情况,也是在水平空间上种群个体之间彼此的相互关系,它是由种群自身特性,种间关系和环境条件等因素综合影响所决定,是种群生物学特性对环境条件长期适应和选择的结果[1-7]。植物种群分布格局是植物种群在群落中所处的空间结构,可定量化描述的基本特征,研究种群分布格局的目的不仅在与了解群落空间结构特征和定量描述种群水平结构,更重要的是揭示格局的成因,种群的某些生物、生态学特性,揭示阐述种群及群落的动态变化等,也可为保护措施的制定提供一定的科学依据[8-14]。
裸果木(Gymnocarposprzewalskii)属于双子叶植物纲石竹科裸果木属植物[15],是亚洲中部荒漠区内较为稀少的第三纪孑遗植物,同时又是亚洲中部荒漠区的特有植物。它的生存与演化可以反映第三纪以来古地中海气候的沧桑巨变及植物的旱化演化过程。裸果木主要分布在中国新疆南部山区、甘肃西部、青海西部、内蒙古西部、宁夏东南部(沙坡头)等局部地区[16-27],在国外分布仅限于蒙古国南部[19-22],主要生长在海拔800~2 800 m 的荒漠地带[25]。
早在1987年,裸果木被中国植物红皮书收录[28],是国家首批公布的二级重点保护植物[17-19],到1997年国家林业局组织国家重点保护野生植物资源调查时又被确定为国家一级重点保护植物[29]。裸果木是构成石质荒漠植被的重要建群种之一。裸果木具有抗干旱、耐盐碱、耐贫瘠、耐风蚀沙埋、耐水蚀、寿命长、根系深等特点,对防止荒漠化,保护和维持荒漠生态平衡起到积极的作用[30]。
由于裸果木分布区自然条件极端严酷,自然更新和繁殖困难,加之近年来受人类各种经济活动的干扰和破坏,裸果木的种群数量急剧减少,分布面积日趋缩小和片段化[22-26]。目前国内外对裸果木的研究,在形态解剖学[31]、引种学[32-34]、水分生理[35-37]、蒸腾强度[38,39]、化学成分研究[40]、花粉形态学[41,42]、组织培养和再生[43]以及细胞学[44]等方面有所报道,而对裸果木种群生态学方面的研究还十分缺乏。
植物种群空间分布格局的研究,可以揭示种群的动态变化及其成因,是植物种群研究的重要内容,是种群的重要结构特征之一[8-10]。开展裸果木的种群空间分布格局研究,可以为这一稀有物种的保护提供科学理论依据。为此,对肃北地区裸果木的分布情况、生境和群落特征等进行了详细深入的野外调查。
1 材料与方法
1.1 研究区自然概况
肃北地区即指甘肃省肃北蒙古自治县,地处河西走廊西端,辖地由南部祁连山地区和北部马鬃山地区2部分组成。南北地区地理位置相距较远,气候差异较大。北部马鬃山地区位于东经95°27′~97°58′,北纬40°42′~42°08′,平均海拔2 000 m,年辐射量为630.53 kJ/cm2,年日照时数为3 316.5 h,年平均气温3.9℃,年平均最高气温18.5℃,最底气温-11.8℃,无霜期为128 d,年平均降水量为85.2 mm,年蒸发量为3 072.9 mm,为典型大陆性气候。南部祁连山地区位于东经94°33′~90°59′和北纬38°13′~39°56′,平均海拔3 000 m,年平均气温6.3℃,年降水量157.2 mm,年蒸发量2 493.3 mm,平均无霜期156 d,平均日照时数2 841.40 h,水源补给除积雪外,冰川占很大比重(表1)。本区位于青藏高原北缘,依据地形和气候特点可分为高寒山区、戈壁平原和山间盆(谷)地3种地貌特征,其面积所占比例分别为49%,27.4%和23.6%[45]。
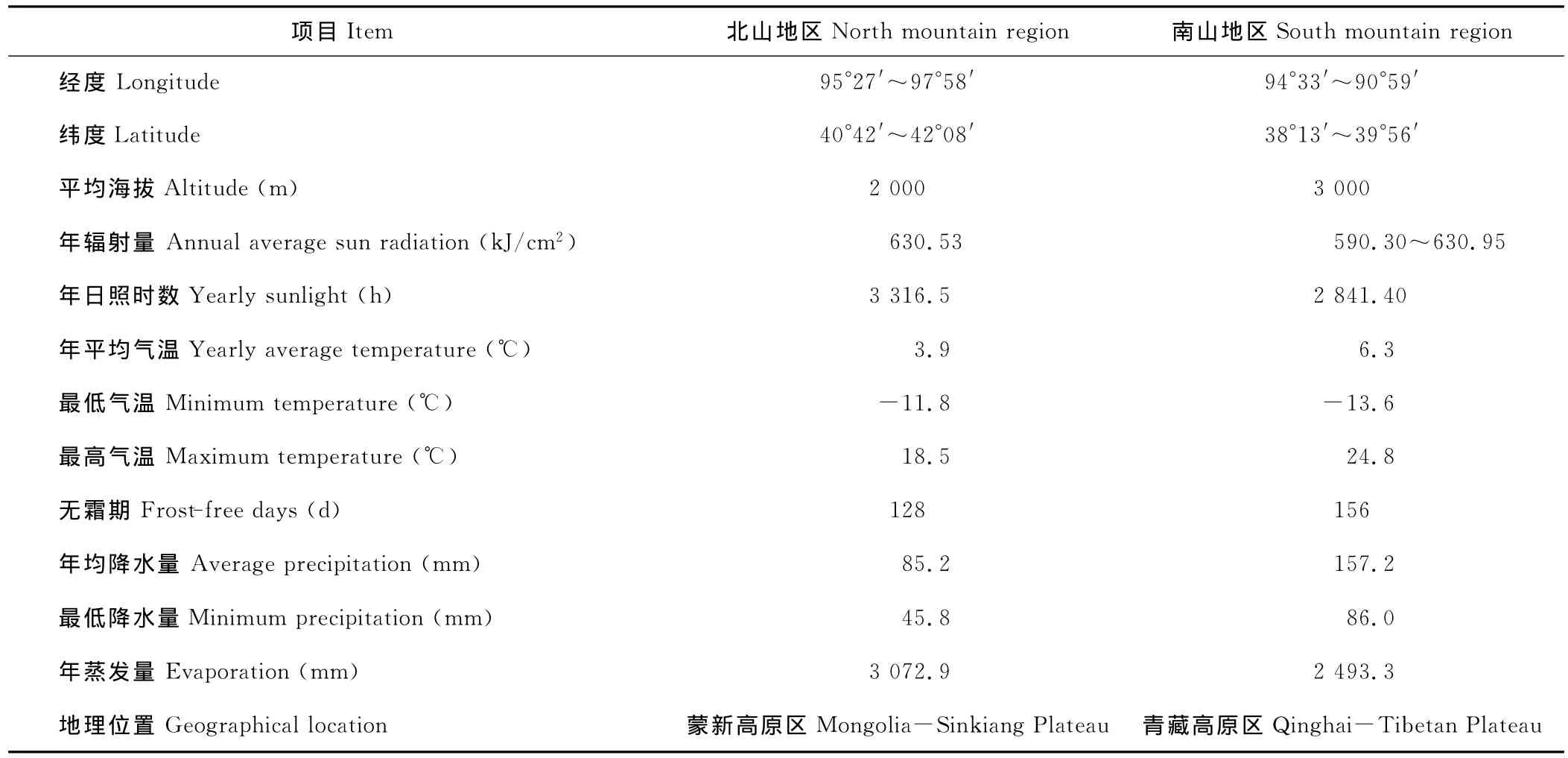
表1 肃北南北山地区自然概况Table 1 The general situation of south mountain and north mountain region of Subei
该地区植物群落主要有合头草(Sympegmaregelii)、中亚紫菀木(Asterothamnuscentrali-asiaticus)、红砂(Reaumuriasoongorica)、灌木亚菊(Ajaniafruticulosa)、裸果木、珍珠猪毛菜(Salsolapasserina)、蒿叶猪毛菜(S.abrotanoides)、戈壁针茅(Stipagobica)、西伯利亚白刺(Nitrariasibirica)、木本猪毛菜(S.arbuscula)、细枝盐爪爪(Kalidiumgracile)、多根葱(Alliumpolyrhizum)、驼绒藜(Ceratoideslatens)、细叶亚菊(Abaniaparvifora)、沙蒿(Artemisiaarenaria)、木本铁线莲(Clematisfruticosa)、霸王(Zygophyllumxanthoxylum)等。
1.2 取样、样地选择和研究方法
野外调查和取样选在甘肃省肃北蒙古族自治县南山地区和北山地区的荒漠草地。这2个地区地理特征和气候差异较大。南山地区地处青藏高原的北缘地带,气候高寒干旱(在南山地区有成片的裸果木荒漠群落)。而北山地区(即马鬃山地区)地处蒙新高原地带,气候极端干旱,这一地带裸果木分布区接近于蒙古国。裸果木在肃北南北两地区相距600 km。该区域既有成片的裸果木荒漠群落,又有裸果木为伴生种的群落。根据肃北地区裸果木集中分布区域、海拔、降水量、裸果木分布范围和分布情况等,设置6个大的调查样地,其中裸果木集中分布区设置4个样地,分别在肃北县红柳峡干沟一带设置1个3 000 m2样地;红柳峡干沟口子一带设置1个3 000 m2样地,红柳峡一棵树一带设置1个3 000 m2样地;红柳峡芦草沟一带设置1个3 000 m2样地(以上4个地带为裸果木集中分布区),另外2个样地分别在肃北县马鬃山地区金庙井、音凹峡一带(海拔较低的地区),设置1个3 000 m2样地;在野马河一带设置1个3 000 m2样地(海拔较高的地区);样地周围有围栏保护(表2)。调查时间为2006年4月-2009年10月。
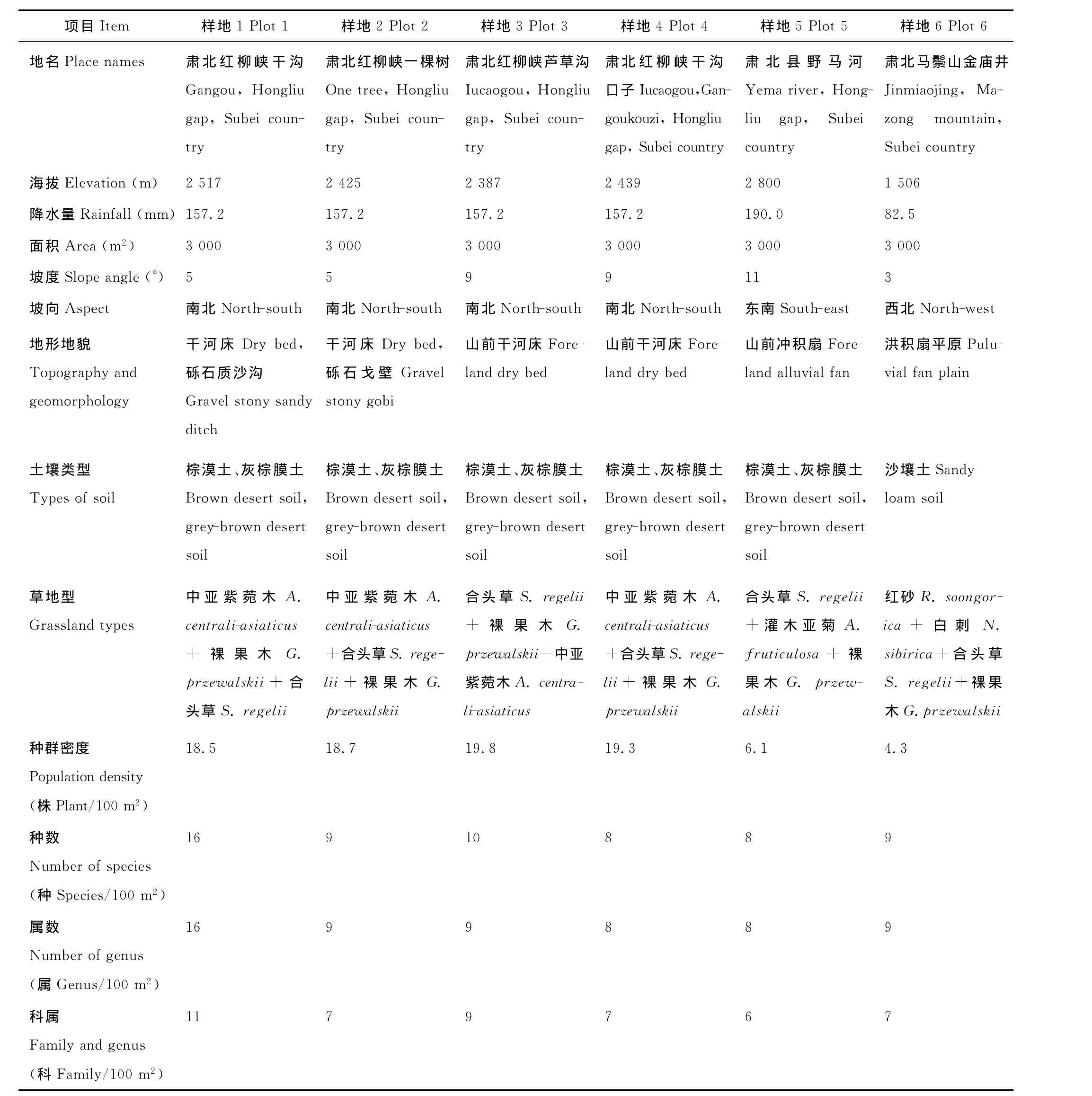
表2 裸果木群落样地概况Table 2 The basic situation of G.przewalskii communities plots
在以上6个样地中,每个样地内随机设定100 m2(10 m×10 m)的样方10个,共计60个样方。进行植物群落特征、裸果木种群特征调查和生境描述。统计每一样方的种属组成、建群种、优势种和伴生种等,在各样地分别测定并计算样方内种群的盖度、密度、频度、高度和多度等,优势度(重要值)、伴生植物种类等。具体调查方法如下[45-47]。
1)盖度:采取样线法测定,样线用50~100 m的皮圈尺。从样线的一端开始测定,以线段为单位测定并登记垂直投影在样线上的植物种名,种的个体所占的线段长度。登记完后整理资料,得样线长度L,总线段数N,样线上出现的植物种数,出现的种的个体数M,种在样线上的总长度B,所有种在样线上的总长度∑B,种以线段为单位出现的次数F。此外另行用样线法按种测定10~20株株丛平均直径r,计算所有种的加权平均直径R。分盖度(%)=B/L×100,总盖度(%)=∑(B/L)×100。
2)密度:单位面积中某种植物的平均株数,用公式表示为:D=N/S(株/m2)。式中,D为密度;N为样地中某种植物的株数;S为样地面积。
3)频度:频度的测定是用0.1 m2的样圆,即用粗铁丝制成的直径为35.6 cm的圆圈,在样地中沿随机的方向随机抛出50~100次,登记每一取样中的植物名录。计算公式为:R(%)=n/N×100。式中,R为频度;n为某一个种在全部取样中出现的次数;N为全部取样数。
4)优势度(重要值):种在群落中所起的作用和所占地位的重要程度叫优势度(SDR)或重要值(IV)。重要值[46-48]=(相对频度+相对高度+相对盖度)/3。
5)裸果木荒漠群落的种属组成调查:记录100 m2样方内的种属组成,统计所有灌丛和草本植物的名称、种类和个体数量、每一物种分盖度,调查统计裸果木以及其他灌木植物的冠幅、高度等。
1.3 数据处理
种群空间分布格局采用偏离指数(C)、丛生指数(I)、负二项参数(K)、LIoyd的平均拥挤度(m*)和聚块指数(m*/m)等进行测定[48,59],分析不同样地和不同海拔下的分布格局。
偏离指数:偏离指数(C)是检验种群是否偏离随机分布的一个系数。其统计基础是Poisson分布中方差等于总体平均数。C=1时,种群属于随机分布;C>1时,为集聚分布(或核心分布,contagious distribution);C<1时,为均匀分布。
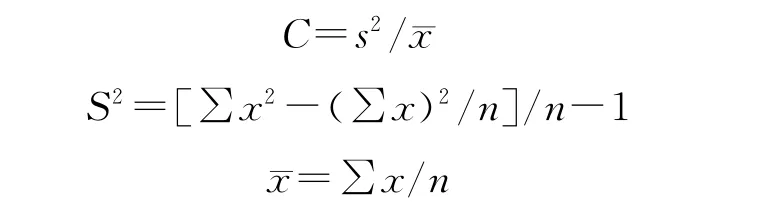
式中,C表示偏离指数,S2表示样本方差,珚x表示样本平均数,n表示样本总数,x表示样方中出现的裸果木个体数。
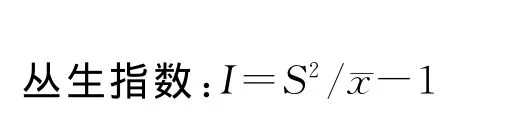
当I=0时,随机分布;I>0时,集聚分布;I<0时,均匀分布。

式中,珚x表示样本平均数,n表示样本总数,xj表示每一样方中出现的裸果木个体数,K表示参数值,K值用于度量集聚程度,K值愈小,聚集程度越高。如果K值趋于无穷大,(一般为8以上),则接近随机分布。显著性检验采用 Kolmogogorov-Smirnov检验[60]。

m*表示一个样方内每个个体的平均拥挤程度。即每个个体在同一单位中其个体的平均数,其数值越大表示该个体受其他个体的拥挤效应越大。由于针对每个个体,其值依赖于现有的总个体数。式中x表示出现的株数,珚x表示样本平均数,S2表示样本方差。
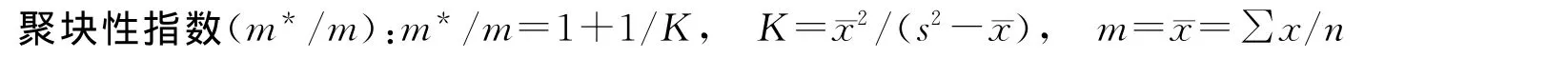
m*/m表示样方内每个个体平均有多少个其他个体对其产生拥挤的程度。
聚块性指数考虑了空间格局本身的性质,并不涉及密度,其值越大,聚集性越强。
式中,m*为平均拥挤度,m为总体平均数,可由样本平均数估计,珚x为样本平均数,x为出现的株数,n为抽样单位总数,K表示参数值,S2表示样本方差。
2 结果与分析
2.1 不同样地裸果木的种群空间分布格局
不同样地裸果木种群空间分布格局判定参数见表3。红柳峡干沟种群和红柳峡一棵树种群的偏离指数分别为2.45和2.57,丛生指数分别为2.61和2.72,负二项指数分布为6.49和6.54,这3个指数均表明这2个种群均为集群分布。红柳峡芦草沟种群和红柳峡干沟口子种群的偏离指数均小于1,丛生指数都大于0,而负二项参数分别为35.79和37.80,远大于8,经Kolmogogorov-Smirnov检验其差异显著性,P值均大于0.8,由此判断这2个种群均为随机分布。野马河种群的偏离指数为0.25,丛生指数为0.30,负二项参数为18.70,经Kolmogogorov-Smirnov检验其差异显著性,P值等于0.07,由此可以判定该种群为近似随机分布,是随机分布与集群分布的混合类型。马鬃山金庙井种群偏离指数为0.47,丛生指数0.61,负二项参数7.06,可判断为集群分布,但集聚程度并不高。
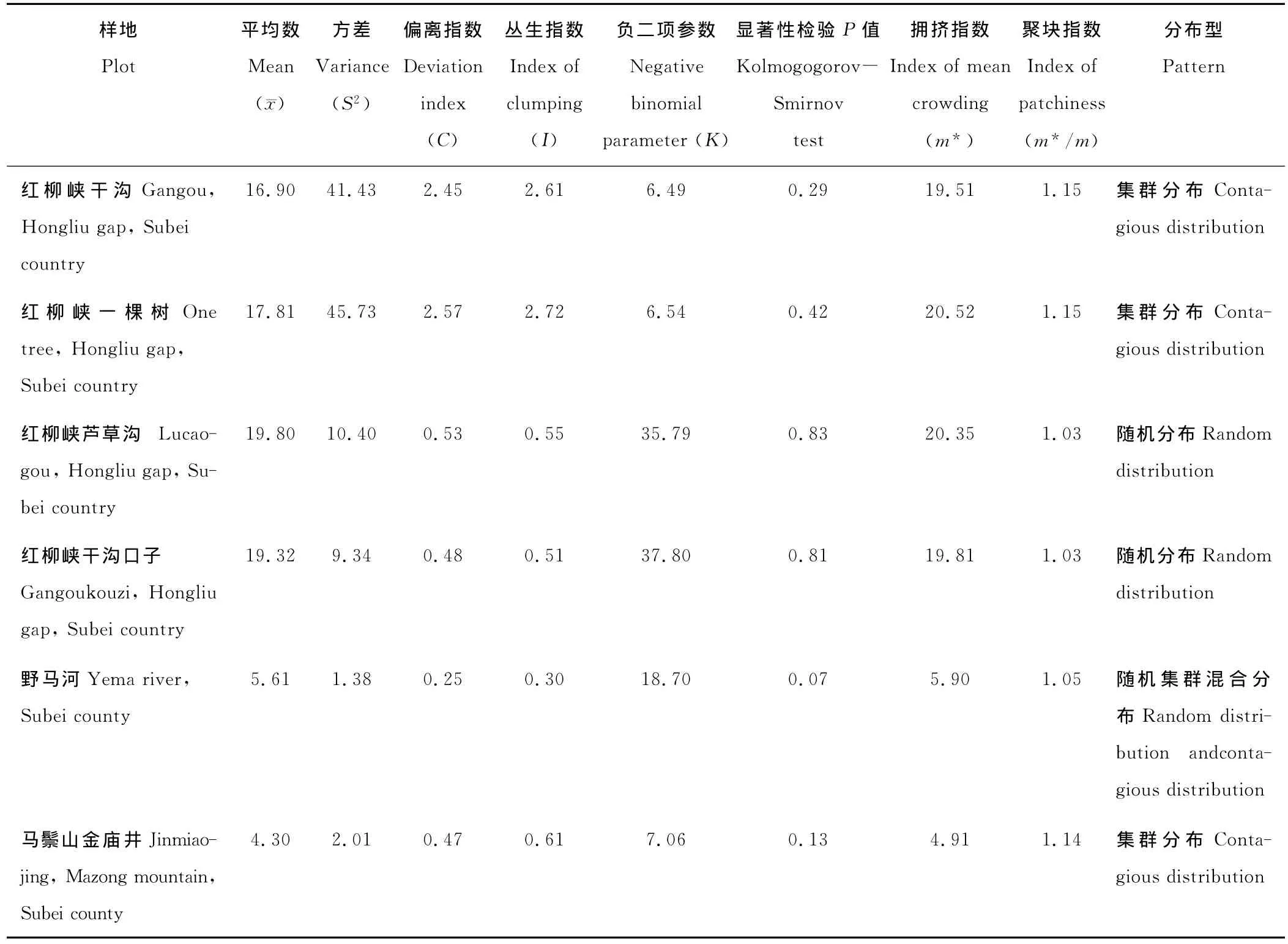
表3 不同样地裸果木种群空间分布格局的判定参数值Table 3 The determined parameter values on spatial distribution patterns of G.przewalskii communities in different plots
2.2 不同海拔下裸果木种群的分布格局
根据研究区域的海拔梯度,分为4个区段,分别为1 500~1 550,2 400~2 450,2 450~2 500和2 700~2 800 m(表4)。海拔在1 500~1 550 m区段,只有马鬃山金庙井种群为集群分布,但集聚程度并不高。海拔在2 400~2 450和2 450~2 500 m区段,偏离指数均大于1,负二项参数均大于12.00,经显著性检验P值均大于0.50,这2个海拔区段均为随机分布。海拔在2 700~2 750 m区段,偏离指数为0.64,丛生指数为0.76,由这2个参数可判断为集群分布。再经负二项参数检验,结果为7.97,即等于8,显著性检验差异不显著(P=0.16),故该区段种群属于集群与随机混合型分布格局。
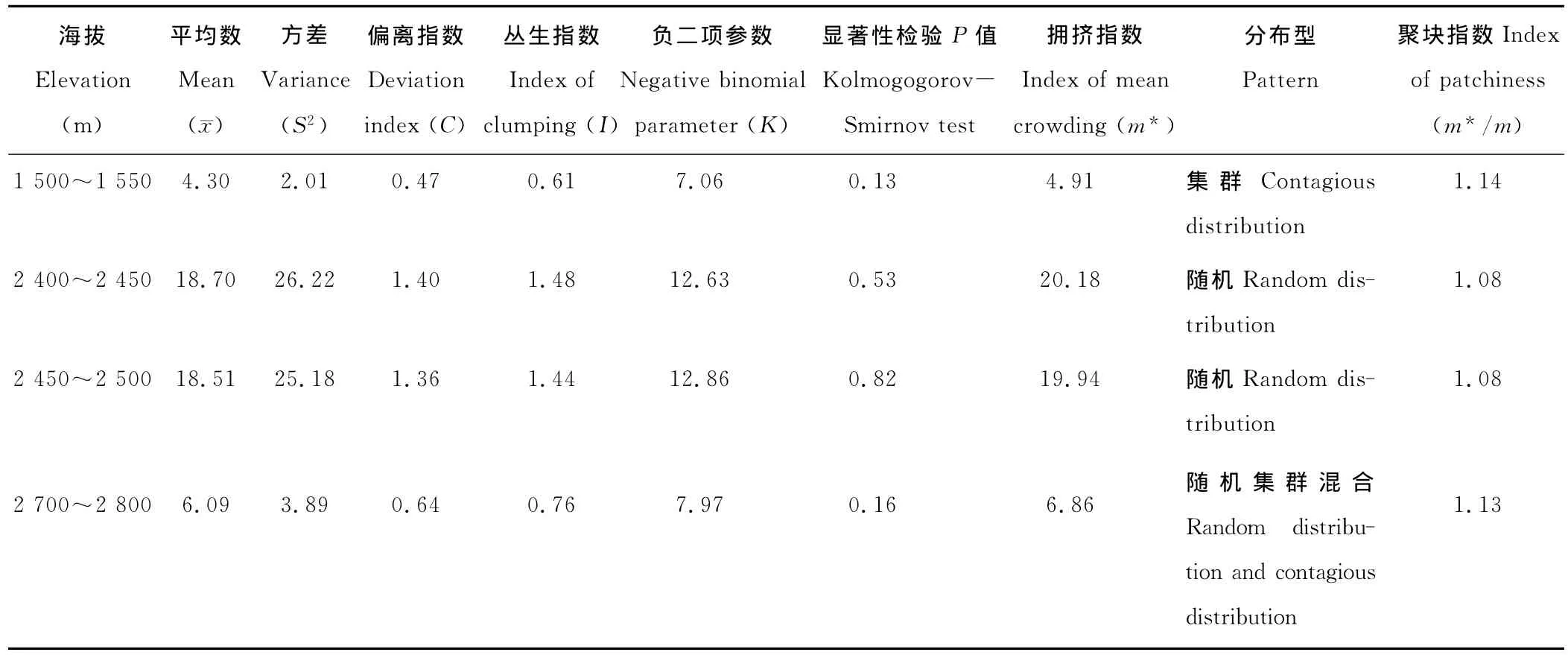
表4 不同海拔下裸果木的种群分布格局判定参数值Table 4 The determined parameter values on spatial distribution patterns of G.przewalskii communities at different altitudes
3 结论与讨论
种群空间分布格局是生物群落中各种内外因素相互作用的结果,不仅与物种的生物学特性和种群间的竞争排斥有关,而且与物种的生境有密切的联系[61]。植物种群的空间分布格局不但因种而异,而且同一种在不同发育阶段、不同的生境条件下也有明显差别。在某种意义上,种群的空间分布格局是种群对环境长期适应的结果,因而种群格局通常反映着一定环境因子对个体行为、生存和生长的影响[62]。种群在空间上的分布状态决定于植物种的生物学特性和生境条件及其相互作用[63]。
3.1 不同样地裸果木种群的空间分布格局
红柳峡干沟和一棵树的干河床、砾石质戈壁,地势南高北低,南北坡度较小,裸果木种群分布为集群分布。这与裸果木的克隆生长特性及其对恶劣环境的适应性密切相关。对裸果木有性繁殖的大量调查和统计发现,开花量很大,但结实率很低,结实率仅为0.7%。在长期干旱的情况下,种子不易萌发,由种子繁殖的幼苗补充量很少,种群数量的有性增殖十分有限。裸果木有性繁殖能力的衰退,使其无性繁殖成为一种有效的补充。在野外调查时发现在干河床和砾石质沙沟裸果木通过根茎和根蘖的方式进行无性繁殖的植株较多,并且在距离母株1~4 m处无性繁殖比较旺盛,有许多侧根经过暴雨和洪水的冲刷后被暴露在土壤外部,而在这些根上通过根茎和根蘖的方式,分蘖可产生3~14个幼苗。最多的有分蘖出21个幼苗。由于无性繁殖在其母株周围分蘖产生许多幼龄的植株,使其分布格局呈现集聚分布。从生物学特性上讲,裸果木具有有性繁殖和无性繁殖的双重能力,当其有性繁殖受到局限时,无性繁殖就作为一种有效的补充方式。从植物适应环境的对策来看,在较大的环境压下,营养繁殖在选择上占优势。这种分布格局特征是对环境异质性的响应和长期处在逆境中的生态适应性[63-67]。
在红柳峡芦草沟和红柳峡干沟口子山前干河床地带,裸果木种群分布均为随机分布。分布区地势南高北低,南北坡度较大。地面有较多的岩石,土壤砾石含量高,生境恶劣。其种群分布格局主要受裸果木自身的繁殖特性、地形地貌、生境和自然灾害等影响的结果。在这一地带裸果木幼龄的植株和幼苗很少,而中老龄的植株较多,中龄以后的植株,由于种间的竞争等因素也是造成这一地带裸果木随机分布的原因[68,69]。用种子繁殖的植物在初入侵时也常呈随机分布。有时候同一种群在同一群落中,可以形成2种分布格局,例如,当某一物种初入侵时,依靠种子自然撒播而形成随机分布,随后则由于无性繁殖而形成集聚分布,最后又由于竞争或其他原因又成随机分布,还有一种情况是,在有些地方为随机分布,而在另外一些地方为集聚分布,这说明分布并不完全是由植物本身的特性所决定的,也与生境条件有关[68,69]。由于这一带地形地貌为山前干河床,地势南北坡度大,裸果木遭受暴雨、洪水和泥石流的破坏非常严重。在每一次暴雨、洪水和泥石流冲刷之后,都有大量的裸果木植株被冲走,并且有许多被洪水冲出土壤而干死的裸果木植株。这也是裸果木种群数量急剧减少重要原因,也是影响裸果木种群分布格局的外部因素。环境的空间异质性,是影响种群空间分布格局形成和发展的主要因素之一,随着时间的变化,植物的分布格局也发生改变[70]。
在肃北野马河山前冲积扇地带,地势北高南低,地势坡度较大,土壤砾石含量高,裸果木种群分布格局呈集聚偏向随机分布型。在这一地区调查时发现,裸果木幼龄的植株和老龄的植株都较少,而中龄植株相对较多。成体以后的裸果木植株,由于种间的竞争等因素也造成这一地带裸果木的随机分布[68,72]。此外由于这一地带地处山前冲积扇地带,地势坡度大,受暴雨、洪水和泥石流的冲刷破坏和水蚀很严重,在野外调查时发现这一带也有许多被洪水冲出土壤而干死的裸果木植株和被洪水冲刷淹没的植株,造成裸果木种群数量的急剧减少。因此在这一带洪水和泥石流的破坏也是影响裸果木种群分布格局的重要外部因素。
在肃北马鬃山金庙井洪积扇平原地带,裸果木种群分布格局呈集群分布,但集聚程度并不高。在这一地区气候极端干旱,降水量稀少,蒸发量很大,干旱是限制裸果木自然繁殖、种群发展和分布格局的重要环境因素。但由于这一带属于洪积扇平原,地势平坦,洪水携带来大量的泥沙沉积物沉积在这一带,使土壤成为沙壤土,土壤疏松,透气性好。在这种疏松的土壤中裸果木的根入土较深,这有利于其吸收深层的水分,以适应极端干旱的环境。同时也有利于其无性繁殖,在泥沙沉积物中,也有从其他地带被洪水携带来的裸果木种子和其他植物的种子,随着泥沙沉积物沉积到这一地带,因此也使这一带物种多样性较丰富,植物种类有11种[27]。由于地势平坦,洪水的缓缓冲刷以及泥沙的淤积和沉积等也是造成这一地区裸果木呈集群分布的原因。
3.2 不同海拔裸果木种群的空间分布格局
3.2.1 肃北裸果木分布的低海拔地区(马鬃山地区,海拔在1 500~1 550 m) 在马鬃山地区海拔1 500~1 550 m的洪积扇平原地带,裸果木种群分布格局呈集群分布,但集聚程度并不高,种群的拥挤度指数也相应降低。这一地区气候极端干旱,降雨量稀少,(年平均降水量为85.2 mm,年蒸发量为3 072.9 mm,最低降水量45 mm),干旱是限制裸果木在这一地区自然繁殖、发展及其种群分布格局的重要因素。在马鬃山地区野外调查时发现,在持续干旱的环境下,裸果木的叶片脱落,成假死状态。在持续干旱的环境下,裸果木开花量很少,甚至不开花。在严酷的环境压力下,裸果木的有性繁殖受到极大限制,种子繁殖的幼苗补充量很少,裸果木种群数量的有性增值十分有限。但由于这一地区为洪积扇平原,地势平坦,洪水携带来的泥沙逐渐淤积在这一地区,使这一带土壤形成沙壤土。土壤中也有从上游其他地带被洪水携带来的裸果木种子,随着泥沙沉积物沉积到这一地带,由于地势平坦,洪水的缓缓冲刷和泥沙的沉积等也是造成这一地区裸果木集群分布的原因。此外也受人为因素的影响。由于这一地区是牧民的春季草场,裸果木是山羊和骆驼喜食的饲料。因此不可避免的受到放牧因素的干扰,同时由于这一地区煤矿较多,煤矿的开采也使裸果木遭到人为的破坏。也影响到裸果木种群的分布格局。
3.2.2 肃北红柳峡地区海拔2 400~2 500 m地带裸果木种群的分布格局 在红柳峡干沟、一棵树山前干河床和砾石质沙沟地带,地势南高北低,坡度较小,裸果木种群分布格局为随机分布。其分布格局是自身的繁殖特性、环境因子、自然灾害等因素共同影响的结果。同时这一带是肃北裸果木的集中分布区,也是裸果木荒漠群落集中的地带,在这一地区裸果木植株的冠幅较大,最大的裸果木植株高为80 cm,冠幅为长263 cm×宽230 cm。通过野外大量的调查,发现裸果木开花量很大,而结实率很低,结实率仅为0.7%,裸果木中老龄的植株较多,而幼龄的植株和幼苗较少,这是由于裸果木在有性繁殖过程中,结实率很低,加之生境恶劣、长期干旱,种子不易萌发,或萌发以后由于持续干旱的原因影响到幼苗的生存。在调查裸果木种子撒落的去向时发现,种子开始撒落时是在母株植株下面和周围1~5 m处,后来被风吹落到其他地带或被洪水冲刷到河床下游地带。由于裸果木生境大多是干河床,土壤是砾石沙土,土壤质地差,砾石含量多,遇到持续的干旱时,种子不易萌发。经过第2年观察种子实生苗情况,在裸果木种子撒落地带,在母株附近或周围观察,实生苗很少,仅有零星分布的几株。而另外一些种子被风吹到其他地带或被洪水冲刷到河床下游。只有在下游方向的一些流水沙沟和细沙沉积地带,才有种子萌发的幼苗。裸果木进行种子繁殖比较困难。裸果木幼龄植株少而中老龄植株多,裸果木在长成成体植株后,由于种间的竞争等因素也造成这一地带裸果木的随机分布。无性繁殖作为一种有效的补充。在野外调查时还发现:在河床地带和砾石质沙沟,裸果木通过根茎和根蘖的方式进行无性繁殖的植株较多,但由于这一地区处在山前干河床、自然环境严酷,生境恶劣,通过无性繁殖分蘖产生的幼苗,能活下来长成成体的植株很少。加之这一地区经常受暴雨、山洪和泥石流的破坏非常严重,在每一次大的暴雨和泥石流的冲刷之后,都有大量的裸果木植株被冲走或淹没。这也是造成裸果木集中分布区,种群数量集聚减少的一个重要原因。裸果木种群的分布格局受暴雨和泥石流等自然灾害的影响很大。这也说明裸果木在恶劣的自然环境中,种群的分布格局并不稳定。
3.2.3 肃北裸果木分布的高海拔地区(野马河地区,海拔2 700~2 800 m) 在肃北野马河一带,地形地貌为山前冲积扇,地势北高南低,坡度较大,裸果木种群分布格局呈集聚偏向随机分布型,种群集聚强度降低,这种分布格局是裸果木自身内在因素——有性繁殖能力衰退、高海拔地区环境因子和自然灾害等共同作用的结果。在野马河地区,海拔2 700~2 800 m,气候高寒干旱、生境恶劣,土壤砾石含量高。由于裸果木开花量很大,而结实率很低,在这一高海拔地区,气温低,种子不易萌发,或萌发后幼苗生长速度缓慢,在野外调查时发现,在这一地区没有冠幅较大的裸果木植株。无性繁殖能力也因气温低而减弱。其分布格局呈集聚偏向随机分布。加之这一地区受暴雨、泥石流等的冲刷和水蚀严重。这也是影响裸果木种群分布格局的重要原因。
3.3 肃北地区裸果木种群空间分布格局的影响因素
裸果木由于其分布区域的局限性,主要分布在在海拔800~2 800 m的山前冲积扇、山前洪积扇、干河床、砾石质沙沟)、由于自身有性繁殖能力的衰退、加之自然环境的严酷和生境的恶劣、干旱少雨、土壤砾石含量高,自然灾害(暴雨、泥石流)和人为因素(放牧、开矿和砍挖)的影响等使裸果木这一稀有物种分布面积日趋缩小,种群数量急剧减少。在红柳峡干沟、红柳峡一棵树、芦草沟一带,裸果木中龄植株和老龄植株较多,幼龄植株较少。从年龄结构上说明种群数量趋于下降,属于一个衰退性种群[33]。总之,在肃北地区裸果木种群分布格局主要受其自身内在因素(繁殖特性),自然环境因素(气温、降水量和海拔等)以及自然灾害和人为因素等多方面的影响。
笔者认为裸果木自身内在因子和外部因素共同影响造成了裸果木种群数量的日趋减少。因此应该进一步对稀有植物裸果木深入研究,比如裸果木对人类干扰,出苗对水分条件的响应等[70],可以与其他植物比对研究[71,72],揭示这一古老孑遗植物的适应机制、生存的关键因子等,从而使这一稀有物种能够得到有效的保护提供更多科学依据。
[1]Greig-Smith P.Quantitative Plant Ecology(3rd edn)[M].Oxford:Blackwell Scientific Publications,1983.
[2]Kershaw K A,Looney J H.Quantitative and Dynamic Plant Ecology(Third edition)[M].London:Edward Amold,1985:50-127.
[3]Gittins R.Canonical Analysis a Review with Applications in Ecology[M].Berlin:Sprinter Verlag,1985.
[4]周纪纶,郑师章,杨持.植物种群生态学[M].北京:高等教育出版社,1992.
[5]谢宗强,陈伟烈,刘正宇,等.银杉种群的空间分布格局[J].植物学报,1999,41(1):95-101.
[6]李毅,胡自治,王志泰,等.东祁连山高寒地区山生柳种群分布格局研究[J].草业学报,2002,11(3):48-54.
[7]韩路,王海珍,周正立,等.塔里木胡杨林主要种群空间分布格局与动态研究[J].西南林学院学报,2007,27(2):1-10.
[8]Pieliu E C.Mathematical Ecology[M].New York:Wiley-Interserence,1985:84-193.
[9]Arista M.The structure and dynamics ofAbiespinsapoforest in Southern Spain[J].Forest Ecology and Management,1995,74:81-89.
[10]魏新增,黄汉东,江明喜,等.神农架地区河岸带领春木种群数量特征与空间分布格局[J].植物生态学报,2008,32(4):825-837.
[11]兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-21.
[12]康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构分布格局[J].生态学报,2007,27(1):389-396.
[13]鲁为华,朱进忠,王东江,等.天山北坡两种退化梯度下伊犁绢蒿种群空间分布的点格局分析[J].草业学报,2009,18(5):142-149.
[14]鲁为华,朱进忠,王东江,等.天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化规律研究[J].草业学报,2009,18(4):17-26.
[15]刘媖心.中国沙漠植物志(第一卷)[M].北京:科学出版社,1985:250-447.
[16]中国科学院植物研究所.中国高等植物图鉴(第一册)[M].北京:科学出版社,1972:619.
[17]吴征镒.中国植被[M].北京:科学出版社,1983:593.
[18]中国科学院中国植物志编辑委员会.中国植物志(第26卷)[M].北京:科学出版社,1996:50-52.
[19]内蒙古植物编辑委员会.内蒙古植物志(第二卷)[M].呼和浩特:内蒙古人民出版社,1978:159.
[20]内蒙古植物志编辑委员会.内蒙古植物志(第三卷)[M].呼和浩特:内蒙古人民出版,1989:158-160.
[21]中国科学院内蒙古宁夏综合考察队.内蒙古植被(综合考察专集)[M].北京:科学出版社,1985:690.
[22]中国科学院兰州沙漠研究所.中国沙漠植物志(第一卷)[M].北京:科学出版社,1985:9-11.
[23]中国科学院新疆综合考察队,中国科学院植物研究所.新疆植被及其利用[M].北京:科学出版社,1978:100.
[24]巴哈尔古丽,汪志军,郭中军,等.珍稀濒危植物裸果木地理分布与资源现状[J].中国野生植物资源,2005,24(5):39-40.
[26]黄大桑.甘肃植被[M].兰州:甘肃科学技术出版社,1997:46-214.
[27]吴玉虎.青海植物名录[M].西宁:青海人民出版社,1997.
[28]柴永青,曹致中,蔡卓山.肃北地区裸果木荒漠群落构成特征及物种多样性研究[J].草业学报,2010,19(1):21-27.
[29]傅立国.中国植物红皮书-稀有濒危植物(第一册)[M].北京:科学出版社,1992:202.
[30]汪之波,马金林.珍稀濒危植物裸果木群落物种多样性及濒危原因初探[J].天水师范学院学报,2007,27(2):55-57.
[31]宋朝枢,徐容章,张清华.中国珍稀濒危植物保护[M].北京:中国林业出版社,1989:135.
[32]汪之波.国家一级重点保护植物裸果木生物学特性的初步研究[D].兰州:兰州大学,2004:1-42.
[33]刘生龙,郭志中,王理德,等.裸果木引种栽培试验研究[J].中国沙漠,1995,15(1):88-91.
[34]刘生龙,王理德,高志海.八种珍稀植物引种试验[J].甘肃林业科技,1995,(3):10-13.
[35]严子柱,李爱德,李得禄,等.裸果木扦插繁殖试验研究[J].甘肃农业大学学报,2005,40(4):550-554.
[36]刘家琼.我国荒漠不同生态类型植物的旱生结构[J].植物生态学与地植物学丛刊,1982,6(4):314-319.
[37]刘家琼,蒲锦春.我国沙漠中部地区主要不同类型的植物的水分和旱生结构的比较研究[J].植物学报,1987,29(6):662-673.
[38]王理德,刘生龙.沙区五种珍稀濒危植物水分生理指标测定及分析[J].甘肃林业科技,1995,23(3):6-9.
[39]蒋志荣.沙冬青抗旱机理的探讨[J].中国沙漠,2000,20(1):71-74.
[40]周宜君,刘春兰,冯金朝,等.沙冬青抗旱、抗寒机理的研究进展[J].中国沙漠,2001,21(3):312-316.
[41]黄玉林,潘伯荣.15种荒漠珍稀濒危植物化学成分分析初步研究[J].干旱区研究,1991,3(3):63-67.
[42]党荣理,姜彦成.新疆裸果木亚科植物花粉形态学研究[J].西北植物学报,1996,1(1):61-64.
[43]莫日根.残遗种裸果木花粉的扫描电镜观察[J].内蒙古大学学报,1993,24(1):94.
[44]汪之波,高清祥,孙继周,等.稀有植物裸果木的组织培养及植株再生[J].西北植物学报,2004,24(7):1319-1321.
[45]肃北蒙古族自治县人民政府.肃北蒙古族自治县志[M].酒泉:酒泉出版社,1989:1-85.
[46]任继周.草业科学研究方法[M].北京:中国农业出版社,1998.
[47]李博.生态学[M].北京:高等教育出版社,2000.
[48]李博.普通生态学[M].呼和浩特:内蒙古大学出版社,1993.
[49]周萍,刘国彬,侯喜禄,等.黄土丘陵区铁杆蒿群落植被特性及土壤养分特征研究[J].草业学报,2008,17(2):9-18.
[50]刘世荣,蒋有绪,史作民,等.中国暖温带森林生物多样性研究[M].北京:中国科学技术出版社,1998.
[51]孙儒泳,李博.普通生态学[M].北京:高等教育出版社,1993:136-138.
[52]孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001:383-436.
[53]张德罡.东祁连山杜鹃灌丛生态系统研究[M].兰州:甘肃教育出版社,2002.
[54]祖元刚,张文辉,阎秀峰,等.濒危植物裂叶沙参保护生物学[M].北京:科学出版社,1999.
[55]查普曼S B[英].植物生态学的方法[M].阳含熙,译.北京:科学出版社,1980.
[56]云南大学生物系.植物生态学[M].北京:人民教育出版社,1980.
[57]林鹏.植物群落学[M].上海:上海科学技术出版社,1982.
[58] 米勒-唐布依斯D,埃仑伯格 H.植被生态学的目的和方法[M].鲍显诚,张绅,杨邦顺,等译.北京:科学出版社,1986.
[59]Walter Larcher[奥地利].植物生态生理学[M].翟志席,郭玉海,马永泽,等译.北京:中国农业大学出版社,1997.
[60]肖枝洪,朱强.统计模拟及其R实现[M].武汉:武汉大学出版社,2010.
[61]程煜,闰淑君,洪伟.檫树群落主要树种分布格局及其动态分析[J].植物资源与环境学报,2003,12(1):32-37.
[62]谢宗强,陈伟烈,刘正宇.银杉种群的空间分布格局[J].植物学报,1999,41(1):95-101.
[63]Jang H .Population Ecology of Spruce(Piceaasperata)[M].Beijing:Chinese Forestry Press,1992.
[64]马克明,祖元刚.兴安落叶松种群格局的分形特征记盒维数[J].植物研究,2000,20(1):104-111.
[65]江洪.云杉种群生态学[M].北京:中国林业出版社,1992.
[66]Grime J P.Plant strategies and vetetation processes[M].Chichester:John Wiley and Sons,1979.
[67]王昱生.羊草种群无性系生长格局[J].植物生态学与地植物学学报,1992,16(3):234-241.
[68]苏智先.缙云山慈竹种群生物结构研究[J].植物生态学与地植物学学报,1991,15(3):240-251.
[69]刘庆,钟章成.斑苦竹无性系种群克隆生长格局动态的研究[J].应用生态学报,1996,7(3):240-244.
[70]何恒斌,张惠娟,贾桂霞.磴口县沙冬青种群结构和空间分布格局的研究[J].林业科学,2006,42(10):14-18.
[71]罗亚勇,赵学勇,黄迎新,等.三种一年生藜科沙生植物出苗对沙埋深度和水分条件的响应[J].草业学报,2009,18(2):122-129.
[72]郑伟,朱进忠,潘存德.草地植物多样性对人类干扰的多尺度响应[J].草业科学,2009,26(8):72-80.
猜你喜欢
今日农业(2022年15期)2022-09-20
特区文学(2020年2期)2020-09-10
今日农业(2019年13期)2019-08-12
药学研究(2019年4期)2019-05-15
人民调解(2019年5期)2019-03-17
红土地(2018年7期)2018-09-26
广西林业科学(2016年2期)2016-03-20
小说月刊(2015年12期)2015-04-23
当代畜禽养殖业(2014年10期)2014-02-27
中国火炬(2009年9期)2009-07-24
