
不同季节放牧对矮嵩草草甸植物叶面积指数的影响
2010-05-13 08:10苏爱玲张振华汪诗平段吉闯常小峰杨晓霞罗彩云徐广平胡宜刚
草原与草坪 2010年1期
苏爱玲 ,张振华 ,汪诗平 ,段吉闯 ,常小峰 ,杨晓霞 ,罗彩云 ,徐广平 ,胡宜刚
(1.中国科学院西北高原生物研究所,高原生物适应与进化重点实验室,青海西宁 810008;2.中国科学院研究生院,北京 100049)
植物对放牧响应的研究是放牧生态学中深入探讨家畜-植物相互作用机制、阐明放牧草地退化机理的重要部分[1]。Gibson和Brown[2]等认为,放牧可以改变植被变化的基本模式,但与植被固有的变化速度和演替方向上的区域变化相比是次要的。许多研究表明,放牧率对不同草地的群落结构和生产力及物种多样性有显著影响[3]。根据植物对采食反应的研究,在生长季期间,采食的时间不同,植物的生长反应不同;且同一时间,不同植物的反应亦不同[4]。由此,将会改变植物个体之间(种内与种间)的竞争关系,进一步将可能影响群落的物种多样性。在高寒草甸地区,由于过度放牧已经导致了天然草地生态系统发生了不同程度的退化[5],但退化草地主要分布在冬春草地,而夏秋草地退化程度较轻。这可能是因放牧季节不同,草地植物从个体水平上和群落水平上对牧食的反应不同。夏季牧场每年放牧5个月,冬季牧场放牧约7个月,所以即使是在同等载畜量条件下,冬季牧场的放牧压力较大,表现为过度放牧。过度放牧是盐漠灌木生态系统退化的重要原因,但放牧季节对植物种类成分的变化影响比放牧率更大[6]。
草地植被结构的主要参数,例如生物量、草群高度、叶面积指数、分蘖密度与茎叶比等能反映草地草群的“质”与“量”的变化,而家畜最适放牧率的确定,不仅依赖家畜畜群本身(群体采食行为),更决定于以上这些草地植被结构特征[7-10]。Dudley和Schmitt[11]研究发现,在放牧或刈割条件下,植物冠层叶面积指数大大减小,使植物群落的光合能力快速下降;但 Wilsey[12]研究发现,植物为了减轻因家畜采食而引起的光合作用下降的影响,往往将新产生的光合产物优先分配给地上部分,尤其是光合器官叶,因而采食后整株植物光合能力的下降未必与所移去的叶面积或生物量成正比,其下降的速率主要取决于采食后保留在植株上嫩叶的相对比例。植物的相对生长速率可以表征植物在单位时间的净积累速率,这一参数可以反映草地植被结构的功能特征变化[13]。
在我国放牧生态学研究中,多数研究利用地上、地下生物量以及物种多样性等指标来表征不同放牧率对草原群落结构和功能的影响[14],有关叶面积指数变化的研究往往是结合光合和蒸腾速率等生理生态学研究进行[15],而对于群落叶面积指数及其与以上指标间关系的变化对不同放牧季节的响应模式的研究较少。通过研究矮嵩草草甸植物群落对不同放牧季节的响应,进行不同放牧季节的可控放牧试验,旨在探讨不同放牧季节对高寒矮嵩草草甸群落植物叶面积指数、群落地上植物量及其相对生长速率、以及叶茎比的影响,为合理利用高寒地区草地及提高生产力提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于青藏高原东北隅的青海海北藏族自治州门源县境内中国科学院海北高寒草甸生态系统定位站,N 37°29′~ 37°45′,E 101°12′~ 101°23′。该区位于亚洲大陆腹地,具有明显的高原大陆性气候,受高海拔条件的制约,气温极低,海北站年平均气温-1.6℃ ,降水量约560 mm。年内无绝对无霜期,相对无霜期20 d,最热的7月仍可出现霜冻、结冰、降雪(雨夹雪)等冬季的天气现象,表现出冷季寒冷、干燥、漫长,暖季凉爽、湿润、短暂[16]。样地内优势植物为矮嵩草(Kobresia humilis)、羊茅(Festuca ovina)、垂穗披碱草(Elymusnutans)、小嵩草(Kobresia pygmaea)、早熟禾(Poa spp)、冾草(Koeleria cristata)、藏异燕麦(Helictotrichon tibeticum)、苔草(Carex spp.)、二柱头藨草(Scirpus distigmaticus)、麻花艽(Gentiana straminea)、线叶龙胆(Gentiana f arre)、美丽凤毛菊(Saccs-surea superba)、雪白委陵菜(Potentilla nivea)等,土壤为草毡寒冻雏形土。
1.2 试验设计
试验设在海北站综合观测场,海拔3 220 m,按随机区组设计,设3个处理,为冬春季放牧,夏秋季放牧和不放牧(对照),重复3次,共设 9个小区,各小区面积为5 m×5 m。为了避免干扰和便于放牧,2006年8月19日在各小区周围设置了围栏。从2007年开始放牧,每个小区每次放进4只大小一致的绵羊自由采食,直到目测植被冠层高度为放牧前的1/2为止,每次放牧时间为一个白天。冬春季放牧和夏秋季放牧小区每年各放牧2次,2007年冬春季放牧时间分别为5月28日和6月13日,夏秋季放牧时间分别为7月13日和8月3日。
1.3 取样方法
分别于2007年7月9日,7月24日和8月9日在各小区随机选取25 cm×25 cm样方,将其地上绿色部分齐地面刈割放于塑料自封袋中带回实验室,分种并将每种植物的茎和叶分开,用美国 Licor公司的Li-3000叶面积仪迅速测定所有叶片的总叶面积,然后在65℃烘箱中烘干至恒重称重。
1.4 指标测定
地上植物量相对生长速率(RGR)=(W2-W1)/(t2-t1)。
式中W1和W2分别表示 t1和t2时刻的地上植物量,单位为g/m2。
1.5 数据分析
所有数据均采用SPSS软件进行放牧季节和取样日期的两因子多重比较分析,以检验在0.05水平上是否差异显著。
2 结果与分析
2.1 不同放牧季节放牧前后地上现存植物量变化
由表1可看出,不管是冬春放牧还是夏秋放牧,每次放牧绵羊对牧草的利用率均在30%~50%,属于轻度—适度放牧。放牧后的8月,夏秋季放牧的地上现存植物量明显低于冬春季放牧和不放牧地上现存植物量,不放牧的最高。因为放牧减小了植物光合作用的面积,导致营养物质的生产和积累下降,从而影响地上植物量的形成。图1表明,7~8月,矮嵩草草甸地上活植物量都是先增加后降低,在7月24日达到峰值,其中夏季放牧矮嵩草草甸地上活植物量值最大,为322.7 g/m2。
2.2 不同季节放牧对叶面积指数的影响
从图2可以看出,7~8月3种处理对矮嵩草草甸植物叶面积指数的影响不同。7月9日夏季放牧显著降低了群落叶面积指数,但到7月24日群落叶面积指数快速恢复到不放牧的水平,并且在7月24日放牧后,到8月8日群落叶面积指数并没有显著性变化;相反,7月24日以冬季放牧群落叶面积指数最低;而8月8日3种处理群落叶面积指数均达到最大值(3.6~3.9)。
从植物地上活植物量与叶面积指数的回归分析可看出(图3),两者之间呈正相关,因此,控制放牧强度,增加茎数和叶面积是提高矮嵩草草甸地上活植物量的重要途径之一。
2.3 不同季节放牧对相对生长速率的影响
植物的相对生长速率(RGR)可以表征单位质量的植物在单位时间内的净积累速率,是衡量植被再生的综合性参数之一。图4的结果表明,3种处理下矮嵩草草甸植物1个月内相对生长速率变化不同,7月9日~7月24日,RGR>0,表明群落地上植物量的净积累呈正增长过程,而7月24日~8月 9日,RGR<0,表明净积累为负增长,植物量下降,这与其地上活植物量的月动态变化规律相一致。
2.4 不同季节放牧对叶茎比的影响
图5表明,7月9日~8月8日,冬春季放牧下矮嵩草草甸各类植物的叶茎比不断增加,8月8日达最大值6;而夏秋季放牧和不放牧对叶茎比的影响则为夏秋季放牧先增加后降低,7月24日达最大值6.9;不放牧先降低后增加,7月24日降到最小值4.3,8月8日达到最大值6.6。
3 讨论
植物叶面积指数为植物群落多样性的一种度量,可以定量表示某一群落地段内植物叶片的密度,是决定太阳辐射强度在群落内削减程度的一个主要因素[17]。李英年[18]等对矮嵩草草甸、金露梅灌丛草甸和藏嵩草沼泽化草甸的叶面积测定表明,3种植物群落的叶面积均表现出相同的变化规律,具有S形曲线变化形式。研究结果与上述研究基本一致,5月中下旬开始随着降水的增多以及温度的升高,植物加速生长后叶面积指数迅速增加,到7月中下旬达最高,以后逐渐降低。叶面积的这种变化与高寒草甸植物的生长过程相联系。
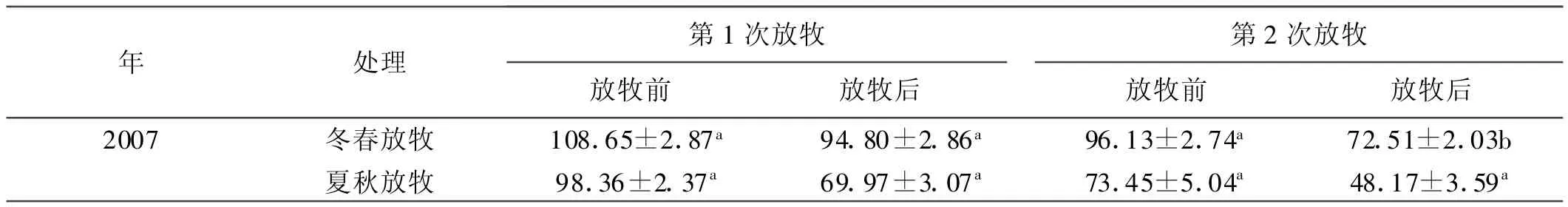
表1 放牧前后地上植物量的变化Table 1 Aboveground biomass before grazing and after grazing
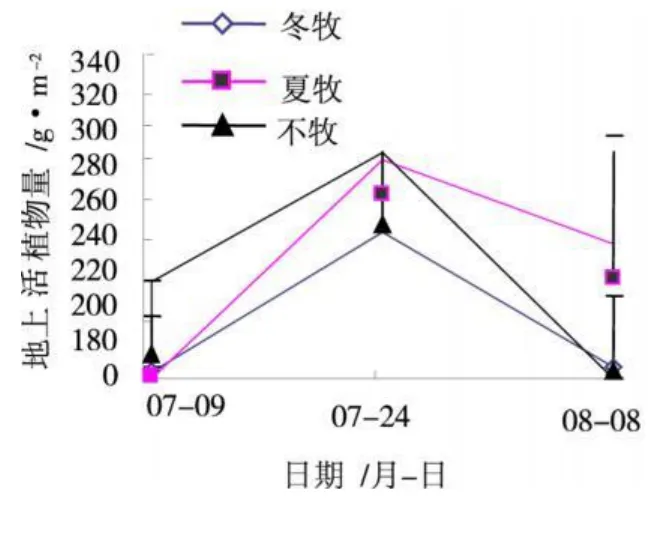
图1 不同季节放牧对地上活生物量的影响Fig.1 The effect of different grazing season on above-ground biomass

图2 不同季节放牧对叶面积指数的影响Fig.2 The effect of different grazing season on LAI

图3 叶面积指数与地上活植物量之间的关系Fig.3 The relationship between LAI and above-ground biomass
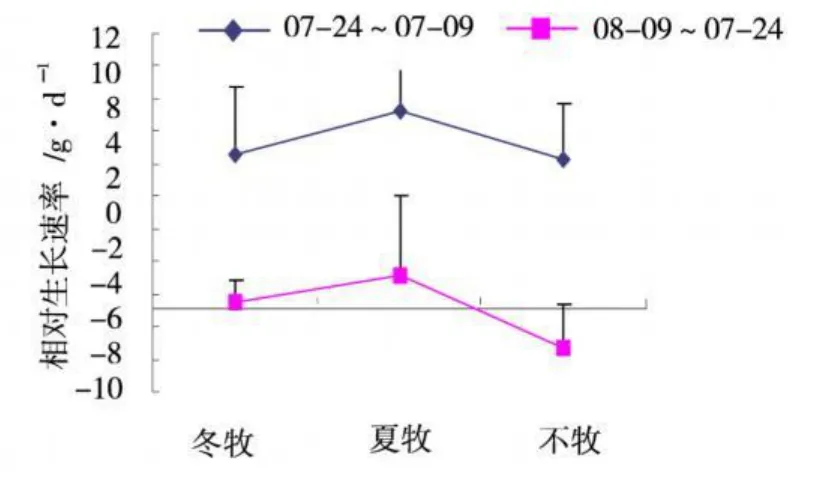
图4 不同季节放牧对相对生长速率的影响Fig.4 The effect of different grazing season on relative growth rate

图5 不同季节放牧对叶茎生物量比的影响Fig.5 The effect of different grazing season on ratio of leaf biomass and stem biomass
地上现存植物量是反应草原生态环境的指标[19],是生态系统研究中最重要的生物物理参数之一,与植物的叶面积之间有着密切的关系。植被地上植物量是群落功能的重要指标,放牧是影响草地地上植物量积累和季节性变化的主要原因之一。而李勤奋等[20]的研究表明,在固定放牧时间长度的情况下,早放牧小区的草群结构比晚放牧的小区受到的影响大,家畜对早放牧小区的牧草利用率高于晚放牧的小区,早放牧小区的草地生产力明显低于晚放牧小区。研究发现,各处理的地上净初级生产力差异不显著。造成以上差异的原因可能,一方面前者放牧小区实行轮流放牧制度,而本试验中,冬春放牧基本上集中在植物的生长期(冬季没有进行放牧),并且冬春放牧和夏秋放牧分别集中在2个时间段,没有形成持续的放牧效应。试验各处理没有发现植物的超补偿生长现象,而汪诗平等[21]对不同放牧率下糙隐子草种群的研究发现,糙隐子草种群在放牧条件下都存在超补偿性生长现象,所以存在一种可能,即放牧草场中一些植物存在超补偿生长,但另外一些植物却表现出欠补偿生长的情况,相互抵消之后个别植物的超补偿生长的效应无法在群落水平上表现出来。本研究结果表明,在轻中度放牧率条件下不同放牧季节对高寒草甸地上现存植物量的影响较大,但对群落的物种组成、多样性的影响不显著。各处理的地上净初级生产力的差异不显著,说明在冬春季放牧处理和夏秋季放牧处理上不存在植物的超补偿生长现象。
高寒矮嵩草草甸植物在不放牧、春季放牧和夏季放牧3种处理下地上活植物量从7月初逐渐增加,到7月下旬达到高峰。此后,随生育期的进程和气温的下降植物进入衰老枯黄阶段,植物量也逐渐减少。整个植物量的月动态与其生长速率的变化相似,呈单峰曲线。李英年等[22]研究发现,在各时期不同植被类型植物量变化的幅度即相对增长率 、植物量达最高的时间均有所不同。矮蒿草草甸植物群落生长最快在6月中旬~8月中旬,在9月15日前后达最高;金露梅灌丛草甸生长最快约出现在7月中旬到8月中旬,并于8月30日前后达最高。不论何种植被类型,其生长过程均遵循逻辑斯蒂曲线增长过程,表现出缓慢积累-快速增加-相对稳定-折损减少等4个不同阶段。只是不同植被类型所处的位置不同,光照、土壤等环境条件也随之不同,所产生的萌动发芽、返青、强度生长、成熟、进入枯黄等出现的时问略有差异,从而导致生长过程曲线不同,主要表现在各主要生长阶段出现的时间发生提前和滞后。研究发现,不放牧条件下其地上活植物量的降低与其地上残留了大量的枯落物有关,枯落物抑制了植物对光和空间资源的利用能力以及植物再生和幼苗的形成,不利于植物的繁殖和更新,从而影响了植物的生长,使得地上活植物量降低。而放牧条件下,不论冬春季节还是夏秋季节,不但改变了草地的土壤环境,更重要的是减少了植物的光合作用面积,导致营养物质生产和积累下降,进而影响了地上植物量的形成。在7~8月份,地上部分植物量一直处于先增加后减少的状态,7月上旬到下旬,随着嫩叶的展开和叶片数量的不断增加,光合速率和效率都上升,植物量不断积累,叶面积指数达到最佳状态,植物量处于快速积累阶段;7月下旬至8月上旬,由于植物叶面积指数继续增加,下层叶片处于遮荫状态,加上叶片不断老化,对光的利用效率降低,光合能力下降,呼吸消耗增强,因此,植物量积累速率变缓。
植物的相对生长速率可以表征单位质量的植物在单位时间内的净积累速率,是衡量植被再生的综合性参数之一,这一参数可以反映草地植被结构的功能特征变化[13]。植物的相对生长速率不仅与生长速率密切相关,而且还取决于现存量。牧草的再生与放牧后草地的地上现存量密切相关,夏季放牧,牧草现存量小,再生所需的物质和能量不能及时得到供应,从而影响到牧草的再生速率;而春季放牧和不放牧条件,由于地上现存量大,地上部分固定的营养物质有很大一部分用于维持其较强的呼吸作用,因此,牧草的再生速率也不会大;只有通过适度放牧,使地上植物量维持在一定的水平范围,再生速率较大,牧草才能得到及时更新。草地种群生产能力和质量特性也与其茎叶构成特点有关,在一定放牧强度和牧草利用率范围内,不仅可以降低牧草枯萎、凋落损失,还有利于草地牧草的更新、再生生长和草地高生产力的保持。Smith[23]的研究表明,随着放牧强度的增加,牧草的再生能力降低,叶生物量及叶数下降。这可能与植物对放牧的适应对策有关,植物为了逃避被采食,减少叶数及叶变小,防止由于大量叶被采食而导致的资源损耗。本文研究是在固定放牧强度的放牧模式下,夏季放牧小区的牧草利用率高,采食强度大,放牧后地上现存量低,牧草的再生速率也低,因此,叶茎比降低。
[1] 马银山,张世挺.植物从个体到群落水平对放牧的响应[J].生态学杂志,2009,28(1):113-121.
[2] GibsonI C W D,Brown V K,Losito L,et al.The response of invertebrate assemblies to grazing[J].Ecography,1992 ,15(2):166-176.
[3] 汪诗平,王艳芬,陈佐忠.放牧生态系统管理[M].北京:科学出版社,2003.
[4] 张东杰,都耀挺.禁牧封育对退化草地的改良效果[J].草原与草坪,2006(4):52-54.
[5] 王小利,张力,张德罡,等.青海湖地区中度退化线叶嵩草型草地地下植物量的研究[J].草原与草坪,2006(1):24-27.
[6] Whisenan S G,Wagstaff F J.Successional trajectories of a grazed salt desert shrubland[J].Plant Ecology,1991,94(2):133-140.
[7] 王德利.草地植被结构对奶牛放牧强度的反应特征[J].东北师大学报自然科学版,2001,33(3):73-79.
[8] 王希群,马履一,贾忠奎,等.叶面积指数的研究和应用进展[J].生态学杂志,2005,24(5):537-541.
[9] 张祝平.鼎湖山森林群落的光能利用效率[J].植物生态学与地植物学学报,1990,14(2):139-150.
[10] 韩发,贲桂英,师生波.不同放牧强度下高寒灌丛植物的生长特点[J].植物生态学与地植物学学报,1993,17(4):331-338.
[11] Dudley S A,Schmitt J.Testing the adaptive plasticity hypothesis:Density-dependent selection on manipulated stem length in Impatiens capensis[J].American Naturalist 1996,147(3):445-465.
[12] Wilsey B J.Variation in use of green flushes following burns among African ungulate species:The importance of body size[J].African Journal of Ecology ,1996 ,34(1):32-38.
[13] 王军强,牛俊义,刘建华,等.模拟放牧和自然放牧条件下线叶嵩草地下植物量的比较[J].草原与草坪,2008(6):5-9.
[14] 徐震,于应文,常生华.放牧强度对黑麦草/白三叶混播草地种群牧草量构成与年生产力的影响[J].草业学报,2003,12(5):31-37.
[15] 鲍雅静,李政海,韩兴国,等.刈割对羊草叶面积指数的影响[J].草地学报,2004,12(4):313-317.
[16] 晁增国,汪诗平,徐广平,等.围封对退化矮嵩草草甸群落结构和主要种群空间分布格局的影响[J].西北植物学报,2008,28(11):2320-2326.
[17] 赵新全.高寒草甸生态系统与全球变化[M].北京:科学出版社,2009.173-174.
[18] 李英年,王勤学,古松,等.高寒植被类型及其植物生产力的监测[J].地理学报,2004,59(1):40-48.
[19] 开花,敖特根,布仁吉雅,等.暖季限时放牧对草地植被的影响[J].中国草地学报,2008(3):28-31.
[20] 李勤奋,韩国栋,敖特根白英,等.划区轮牧中不同放牧利用时间对草地植被的影响[J].生态学杂志,2004,23(2):7-10.
[21] 汪诗平,王艳芬.不同放牧率下糙隐子草种群补偿性生长的研究[J].植物学报:(英文版),2001,43(4):413-418.
[22] 李英年,王勤学,古松,等.高寒植被类型及其植物生产力的监测[J].地理学报,2004,59(1):40-48.
[23] Smith S E.Variation in response to defoliation between populations of Bouteloua curtipendula var.caespitosa(Poaceae)with different livestock grazing histories[J].American Journal of Botany,1998,85(9):1266-1272.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
今日农业(2021年10期)2021-07-28
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
活力(2019年21期)2019-04-01
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20
河北林业科技(2016年5期)2016-11-08
中国麻业科学(2015年5期)2015-12-28
西藏科技(2015年11期)2015-09-26
