
猪及猪肉中沙门氏菌快速检测的研究进展
2010-05-09 07:12耿士忠潘志明刘志成焦新安
猪业科学 2010年6期
耿士忠,潘志明,刘 杰,刘志成,方 强,焦新安
(扬州大学江苏省人兽共患病学重点实验室,江苏 扬州 225009)
沙门氏菌是常见的人和动物的重要病原菌之一,呈全球性分布。多种沙门氏菌均可感染猪,主要包括:海德堡沙门氏菌(Salmonlla heidelberg)、华盛顿沙门菌(Salmonlla worthington)、鼠伤寒沙门氏菌(Salmonlla typhimurium)、鼠伤寒沙门氏菌哥本哈根变种(S.typhimurium-var.copenhagen)、新生儿德尔卑沙门氏菌(Salmonlla derby)、 婴 儿 沙 门 氏 菌(Salmonlla infantis),以及乙型副伤寒沙门氏菌(Salmonlla paratyphi B)、斯坦利沙门氏菌(Salmonlla stanley)、雷根特沙门氏菌(Salmonella regent)、猪霍乱沙门氏 菌(Salmonlla choleraesuis)、 肠 炎沙门氏菌(Salmonella enteritidis)等血清型。其中猪霍乱沙门氏菌和德尔卑沙门氏菌对猪具有宿主适应性,感染母猪可长时间携带病原。世界上最大的一起由沙门氏菌引起的食物中毒,1953年发生在瑞典,是由食用猪肉而引起的鼠伤寒沙门氏菌中毒,导致7 717人中毒,90人死亡。由于沙门氏菌菌型、菌株不同,致人发病的菌量也不同,一般使人发病的菌量平均为107cfu以上。鼠伤寒沙门氏菌是最常见的血清型,在国外占27.7%~80%,其次为肠炎沙门氏菌占10.3%。某省10年来的统计资料显示猪在屠宰前感染沙门氏菌是比较严重的,其检出率高达26.5%。由于带菌率高,在屠宰加工过程中,往往可造成猪肉污染,猪肉的沙门氏菌检出率为12.5%。肉类食品从宰杀到烹调加工的各个环节中,都可受到污染。烹调后的熟肉,如果再次受到污染,并且在较高的温度下保存,食用前又不再加热,则更为危险。有些细菌主要在生长猪以及一些母猪的肠道内繁殖。感染猪可连续数周甚至数月从粪便中排出病原,而不表现任何症状。在屠宰的时候,猪肠道中的沙门氏菌可能污染胴体,导致人类食物中毒,对公共健康构成潜在威胁。有些细菌如猪霍乱和猪伤寒沙门氏菌,可引起仔猪发病,是仔猪的一种较常见的肠道传染病,而在成年猪中很少发生。该病在各地猪场均有发生,特别是在卫生条件差的猪场常有发生,给养殖业造成很大损失。此病主要危害1~4月龄断奶仔猪,所以是规模化猪场提高仔猪成活率与经济效益的大敌[1]。由于这些沙门氏菌是多种猪肉产品的污染源,因而猪的沙门氏菌病备受关注,全世界所有国家都非常重视沙门氏菌控制。
沙门氏菌主要寄生于人和动物的肠道中,但由于其在环境中具有较强的生存能力,所以在环境中也广泛存在,常见于农场的污水、人类生产的污物以及被粪便污染的物体中。该细菌通常由亚临床感染动物的排泄物通过施肥或浇灌等方式造成环境源性污染,造成水果蔬菜等二次污染,引起人间接感染,或者引起食源性污染,如肉、蛋、奶等直接污染,引起人因食用污染食物而感染,并致人腹泻或系统性疾病。猪沙门氏菌病引起的严重的公共卫生问题,对猪肉的食品安全及国际贸易形成较大威胁。因此对猪沙门氏菌的检测及诊断就显得至关重要。
自19世纪后期,沙门氏菌首次被鉴定为人类的一种病原以来,检测方法学都是建立在采集感染动物或患病者的粪便或血液作为临床病料的基础上。此后的60年间,用于从食品中分离沙门氏菌的方法实质上与那些用于临床病料的方法是相同的。但至少有3个因素限制了用于临床病料的方法应用在食品分析上。第一,通常,沙门氏菌的含量水平在污染食品中比在感染病人的病料中要低很多;第二,食品本身的性质会干扰病原的检测,例如,某些食品中固有菌群可能处在一个很高的水平,从而影响特定细菌的选择性分离和鉴定;第三,与临床病料不同的是,经过加工的食品,由于加热、干燥、高含盐量、酸和冷冻等因素的作用,其中的沙门氏菌受到了尚不致命的损伤或称“致伤”,这就形成了一个具有不同生长特性的细菌群,这种现象对那些希望从食物样品中分离出沙门氏菌的食品分析家来说有很大影响,因为在选择培养基上直接培养“致伤”的沙门氏菌通常是以细菌死亡和试验失败而告终。为克服这些困难,人们建立了一种简单的微生物增殖步骤,专门针对以食品为传播载体的病原。虽然这些方法本身证明是可靠的,但却很费力、耗时,需要3~6 d才能完成。因此,在需要及时、快速评价食品中微生物的安全性时,通常不被采用。随着DNA和单克隆抗体技术的发展,近几十年间发展了许多改进的方法,其中一些方法可以在12~24 h内检出沙门氏菌,这些方法通称为快速检测。

图1 脉冲场凝胶电泳(PFGE)仪

图2 实时定量PCR仪

图3 磁分选仪
1 直接法——传统的培养鉴定方法
用于检测沙门氏菌的传统方法是将食物样品分步增菌,以提高病原菌的检出率。这种培养方法总体可分为3个不同阶段,预增菌、选择性增菌及分离步骤。传统沙门氏菌检测法全过程需时至少4~7 d,才能得出明确的诊断结果。GB 4789.4-94是目前我国规定的对畜产品中沙门氏菌的标准检测方法,主要是根据沙门氏菌的生化特性,进行预增菌、选择性增菌、分离培养、生化鉴定和血清分型5个步骤。在检测畜产品过程中,应注意根据不同的样品采取不同的检测步骤。第1步预增菌,不表现临床症状的动物粪便、环境样品、动物饲料和食物中的沙门氏菌数量通常很低,必须使用预增菌培养基(如缓冲蛋白胨水),它能使少量的沙门氏菌增殖,以利于细菌分离,或使用普通预增菌液以帮助细菌分离。将样品添加到一种高营养、无选择性的培养基中,37 ℃培养,有助于因被冷冻、加热、干燥或生物试剂等因素而致于濒死的沙门氏菌复苏。预增菌对较少的生长较活泼的沙门氏菌可能不是最好的方法,如来自粪便的热适应菌株,因为在非选择性预增菌过程中,竞争性细菌生长过快而难以分离。第2步是选择性增菌步骤,它能选择性地使沙门氏菌生长,而抑制其他细菌的生长,有时也对某些血清型的沙门氏菌有抑制和致死作用,如亚硒酸能抑制和致死猪霍乱沙门氏菌,亮绿对都柏林沙门氏菌有害。除了在培养基中添加特定成分外,还常常用提高温度的方法来增加增菌培养基的选择性,不少实验室采用的温度为43 ℃。目前应用的培养基主要有如下几种类型:含四磺酸钠的Muller-Kauffman肉汤、亚硒酸盐F-亚硒酸盐半胱氨酸-亮绿磺、Rappaport-Vassiliadis培养基或半固体Rappaport-Vassiliadis培养基、连四硫基盐肉汤(Tetrathionate broth)、硒酸盐胱氨酸肉汤(Selenitecystinebroth),现在Rappaport-Vassiliadis培养基的推荐温度为41.5 ℃。活力选择增菌培养基用来增加沙门氏菌的敏感性,半固体培养基,如MSRV或DIASALM(诊断用半固体培养基)可以提供更好的选择性。由于没有任何一种培养基可以全面地保持所有食品基质或各种沙门氏菌血清型,所以,较适当的做法就是使用2种培养基平行地进行试验。从某种程度上讲,使用多种选择性培养基或通过预增菌培养后,直接选择性增菌、直接涂板会更有利。第3步选择性平板培养,作为选择性固体培养基,能进行不同程度的鉴别培养,能抑制其他细菌生长,并能提供沙门氏菌的一些主要生化鉴别特征:无乳糖发酵和硫化氢产生,37 ℃培养24~48 h后判定结果。在这种培养基上,沙门氏菌形成特征性菌落,据此通常可与其他细菌菌落相区别。现在有一种较宽范围的显色培养基可以方便使用。有许多这样的培养基有助于分离可疑菌落,但是必须确认样品的基体、培养系统和血清型,因为在某些环境下,细菌的感受性是很差的。第4步是分离鉴定步骤,即选择性培养物在含一种或多种抑制非沙门氏菌生长制剂的琼脂平板上划线培养,然后对平板上肉眼可见的特征性可疑菌落进行确认,并将可疑菌落在选择和非选择性琼脂上作继代培养,对该菌落分离物进行一系列生化和血清学检测,以作出鉴定。鉴定O、H抗原和在特定的情况下鉴定Vi抗原,可用特异抗血清进行直接平板凝集试验。在具有双相菌的情况下,应通过含已知相抗血清的半固体培养基传代,用相转化法鉴定2个相,尽可能使用抗多种因子的抗血清加速筛选,因为单因子血清能更进一步纯化细菌。大多数实验室能鉴定较为普通的血清型,而进一步鉴定某菌株应参考实验室的设备。传统沙门氏菌检测法全过程需时至少4~7 d,才能得出明确的诊断结果。
噬菌体对细菌有特殊的裂解作用。Thal-Kalings对 1181 株 沙 门 氏 菌 和323株其他菌株进行沙门氏菌噬菌体O-I裂解试验,结果表明沙门氏菌裂解率为99.5%,而非沙门氏菌裂解率为0.3%。截至到1973年,Keils及Seeliger等人通过大量观察证实,沙门氏菌属内裂解率在95.0%~99.6%之间,属外误差为0%~2.33%之间。何晓青等发现一种肠杆菌科分属诊断噬菌体,其鉴定沙门氏菌只要6 min,加上培养菌落时间只需2 d。张碧波等应用噬菌体快速检测沙门氏菌,对100个肠杆菌菌株进行噬菌体O-I裂解试验,30株沙门氏菌全部为阳性,而其余非沙门氏菌株全部为阴性,与前人研究结果一致。由此可见,应用沙门氏菌噬菌体O-I对沙门氏菌进行检测方便可靠。
2 间接法
2.1 以抗体为基础的检测方法
利用抗原—抗体反应的显著特异性,来进行细菌的鉴别和血清学定型,已有半个多世纪的历史。细菌菌体或鞭毛抗原的特异性抗体的存在,使得人们可以建立一些快速方法来检测以食品为载体的病原。已经建立的沙门氏菌免疫学检测方法有许多种,大致可分为以酶标抗体(ELISA)、荧光抗体染色(免疫荧光法)、同位素标记抗体(放射免疫试验)为基础的方法及其他多种以抗体为基础,利用乳胶凝集、免疫传感器、免疫扩散及免疫色谱技术的方法。但最广泛采用的是以双位点ELISA技术即夹心ELISA为基础的方法。此法改进后用化学发光替代标记抗体,概括地说,是以固定在固体基质上的“捕捉”抗体来捕捉目标抗原,经洗涤除去未结合的成分,加入第2种酶标抗体,此者结合在捕捉到的抗原的不同位点上,第2次洗涤后加入酶作用基质,并令其与颜色成分反应,然后用分光光度法即很容易检测到目标抗原。采用微量滴定板作为固态基质使反应形式标准化,并促成其自动化。
HerdChek Swine Salmonella Antib ody Test Kit主要检测 B、C1、D血清型的沙门氏菌,在欧洲、亚洲和美洲等地区广泛使用。ELISA法检出沙门氏菌的极限范围在105~106cfu/mL,因此,要得出可靠的结果,食物样品首先必需进行预增菌、选择性增菌,通常还要在含有D-甘露糖的肉汤(M肉汤)中进行后增菌,以促进鞭毛发育。总的来说,标准的ELISA法样品的制备,约需要经过40~48 h的孵育才能完成。样品制备过程分3步:1)选用营养肉汤进行预增菌,使致伤、冷冻的沙门氏菌复苏;2)使用选择性培养基进行增菌,使沙门氏菌大量繁殖,同时抑制其他杂菌生长;3)使用营养肉汤(蛋白胨水)进行后增菌,使沙门氏菌的数量大大增加。比上述标准的ELISA法样品制备过程缩短了一半的时间。ELISA方法本身,则仅需要大约2 h而已(其中30 min是操作时间,90 min是孵育时间)。
最新的沙门氏菌免疫学检测法利用经特异性抗体敏化的免疫色谱卡片为基础。几滴样品加到卡片上,结果可以直接用肉眼读出。免疫色谱卡片极易操作,且由于无需要特殊设备,很适合小型实验室使用。尽管卡片检测法与ELISA法一样需要对样品进行增菌处理,但它(卡片法)本身通常需时不超过10 min。
文其乙[2]等建立了直接ELISA检测沙门氏菌方法,从已获得的4株沙门氏菌属特异杂交瘤单克隆抗体中,筛选出CB8和De7两株单抗相合成酶标检测试剂,并建立了检测沙门氏菌的直接ELISA方法,它能检出沙门氏菌属99%的菌株,而不与其他肠道杆菌反应(包括大肠杆菌、阴沟杆菌、产气杆菌、志贺氏菌、枸橼酸杆菌、克雷伯氏菌、变形杆菌和沙雷氏菌)。应用本方法检测粪样、鱼粉、奶样,其敏感性和特异性分别高达100%和97.6%,仅出现2.4%的假阳性率,该法不受各种样品成分的影响,而且在2~3 d即可完成样品筛选。直接ELISA具有快速简便、易于推广的优点。
殷月兰[3]等建立了单抗竞争ELISA检测猪沙门氏菌感染方法,以猪霍乱沙门氏菌为包被抗原,利用沙门氏菌O7单克隆抗体酶结合物建立了竞争ELISA方法检测猪霍乱沙门氏菌抗体。抗原包被浓度为 2×108cfu/mL,血清检测稀释度为1∶16,酶标抗体浓度为1∶700,包被液为碳酸盐缓冲液(0.05 mol/L,pH9.6)。应用单抗竞争ELISA和平板凝集试验(PAT)同时对506份血清进行猪霍乱抗体检测,PAT的检出率为3.60%,ELISA检出率为5.75%,两者符合率达96.4%。试验结果表明,ELISA方法敏感性高,特异性强,稳定性和重复性好,操作简便。
黄素珍[4]等建立了检测肠炎沙门氏菌的竞争ELISA方法,对210份肠炎沙门氏菌感染鸡泄殖腔棉拭子、羽毛和组织样品先筛选后鉴定的方法进行检测具有明显的竞争抑制作用。通过与国标法对大量的样品检测结果比较表明,竞争ELISA方法的检出率为18.09%,检出阳性样品36份,国标法检出率为17.14%,两者符合率为97.14%,竞争ELISA敏感性和特异性分别为94.4%和97.7%。
斑点酶联免疫吸附(Dot-ELISA)试验是一项免疫学检测技术,具有简单、快速、敏感、特异性强等特点,并具有较高的可重复性。目前,应用该法检测食品中沙门氏菌的报道很多。张红见[5]等应用Dot-ELISA法对85份猪胴体进行了沙门氏菌的检测,结果表明,在85份肉样中,Dot-ELISA检出沙门氏菌阳性65份,阳性率为76.47%,检测结果与常规方法一致。康明[6]等应用Dot-ELISA检测鱼粉85份,结果表明,64份为阳性,常规分离培养法53份为阳性,2种方法差异不显著。由此可见,该方法简便、特异、快速、结果直观,便于在基层推广使用。
由于食品检测样品常为固液多相混合体,采用常规方法难以将少量致病微生物分离出来。借助免疫磁性分离技术,可以很快地在含有大量杂菌的悬液中有选择性地分离出目的微生物,节省时间。目前,应用该法分离检验食品中的致病菌也有一些相关报道。Skjerve等报道了用免疫磁性分离技术从乳及乳制品、肉类和蔬菜中分离出沙门氏菌,其检测限为每克100个细菌。Mansfield等通过比较免疫磁性分离技术和常规方法从食品中分离沙门氏菌,他们用120种食物样品,其中一半样品中加入低浓度的沙门氏菌[7]。结果证实,就选择性而言,抗沙门氏菌磁珠和增菌方法中最有效的亚硒酸盐选择性培养基一样有效。值得强调的是,免疫磁性分离技术能捕获受损伤的靶细菌,而目前所用的几种常规方法则不具备这样的能力。
全自动荧光酶标免疫分析仪对沙门氏菌的检测鉴定结果比较成熟,并已先后被美国FDA、AOAC、USDA等部门认可,我国学者也有相关研究报道。寇运同等利用自动荧光酶标分析系统快速检测出口动物性食品中沙门氏菌,结果表明,其灵敏度高,操作简便,大大缩短检验周期,可用于检验出口动物性食品中沙门氏菌。陶军[8]等利用法国生物-梅里埃公司提供的“自动酶标免疫检测仪”与常规培养法对冻肉中沙门氏菌进行检测,指出该仪器最大特点是对被检细菌不需要纯培养,只需在增菌培养基中即可检出。黄玲[9]等利用自动酶标免疫测试仪和国标方法检测食品中沙门氏菌,结果表明,该法具有灵敏度高、无传染危险、检测速度快等特点。陈炜[10]等运用自动荧光酶标分析仪和国家标准方法分别对同一种脱水蔬菜样品中沙门氏菌进行检测,并对检测结果进行对比,表明运用自动荧光酶标分析仪检测脱水蔬菜中的沙门氏菌灵敏度高、速度快,完全满足检验检疫系统快速检测的需要。
Munron应用自动荧光抗体检测系统检测食品中的沙门氏菌,每小时能检测到120个玻片样品,与常规试验比较,准确率达96%。Kyrinr和heimarch用荧光抗体检测系统检测食品中的沙门氏菌,该系统可同时检测14个样品,并且结果可靠,操作简单。Cloak等利用表面吸附将检测样品吸附于玻璃表面的一种薄膜上,然后用免疫荧光显微镜进行结果判定。该方法检测限为103~105cfu/mL,且不呈现假阴性与假阳性反应。
英国Matrix公司是一家专业从事致病菌快速检测系统研究、开发和生产的专业公司,其采用的免疫磁分离(IMS)方法快速检测致病菌系统,是目前世界上速度最快的致病菌检测方法之一。该方法已经获得美国AOAC认证,具有高灵敏度、高特异性、高稳定性、无污染、无毒性、检测迅速等特点。它采用一种合成磁性材料—微磁珠,磁珠表面进行基团修饰并键合连接臂,将多克隆抗体或单克隆抗体连接后进行包被,制成带磁性的免疫抗体。磁性抗体可以选择性吸附流动样品中的目标抗原,借助外磁场的作用吸附在捕获区,实现对大容量样品中微生物的快速免疫浓缩纯化。
在抗原富集问题上,也有利用单克隆抗体标记磁珠方法,直接将样品与标记磁珠混合,再进行下一步ELISA检测。MaxSignalTM沙门氏菌酶联免疫检测试剂盒为国家管理部门、食品生产商、质检部门提供了方便有效的方法,检测水平为 105cfu/mL。
2.2 以核酸为基础的方法
2.2.1 核酸杂交
细胞核酸DNA和RNA是一类可以携带生物信息的大分子。由于所有的细胞都含有这类分子,可以利用它作为检测的标靶。标靶通常是一个特异性核酸序列,能够结合核酸分子作为探针。与免疫学方法相似,探针也需要附加适当的标记,如放射性同位素、酶或发光的标记物。Fitts等人1983年在食品沙门氏菌检测中引入了第1代DNA-RNA杂交技术,此法应用的探针含有用放射性同位素标记的伤寒沙门氏菌DNA片段,其敏感性高,经大约48 h的增菌步骤后,检测极限可达108cfu/mL,但由于要使用放射性同位素,只能在专门的实验室应用。为此,发展了以核酸杂交为基础的第2代技术——比色计。这种方法依赖于沙门氏菌核糖体RNA(rRNA)-核糖体发育过程中储存的核酸成分的检测[11]。核糖体是细胞蛋白质合成器的一部分,每个细菌细胞中存在 5 000 ~ 20 000 个复制体,而相比之下染色体DNA复制体仅为2~10个。这种天然富含rRNA标靶序列的情况使得用其他方法检测成为可能,同时又保持了与放射性同位素方法相当或更高的敏感性。其另一优点是由于rRNA为单链(而DNA为双链),杂交前无需经过变性步骤。要得到阳性结果,此法需要105cfu/mL,因此对沙门氏菌检测来说,需要进行预增菌和选择性增菌,总共约50 h。rRNA探针法比沙门氏菌ELISA法(酶联免疫检测法)更耗时,但二者成本相近。食品细菌检测法的最新进展是在化学扩增体系方面,即聚合酶链反应(PCR),用该体系可对制备好的样品进行细菌DNA扩增,以便更易于用诸如凝胶电泳法或比色型ELISA法检测。用于检测沙门氏菌和其他以食物为载体的病原的PCR方法也已建立,其中一些方法显示了极好的敏感性。但该法较难自动化,且必需经选择性增菌以稀释可能干扰检测反应的某些成分。一个完整的以PCR为基础的方法,需要2 d才能完成,很大程度上抵消了其高敏感性的优点,但这种方法对那些含沙门氏菌较少或沙门氏菌“致伤”严重难以复苏而仍保有沙门氏菌rRNA的样品尤其有效。
2.2.2 PCR 法
Rahn等首先设计出一对引物,用PCR检测沙门氏菌,检出率为97%,但许多肠道杆菌能同时扩增出来,特异性较差。此后,国内外许多学者进行了利用PCR技术检测沙门氏菌的研究。Nguyen等根据肠炎沙门氏菌C7克隆株具有的属特异性序列设计出一对引物,能快速地检出肉食品标本中的沙门氏菌,检测的敏感性和特异性均为100%,保证了检测的准确性。此外,我国卢强[12]等将自己建立的PCR扩增invA基因特异性检测沙门氏菌的方法用于食品样品中沙门氏菌的检测,并结合辣根过氧化物酶直接标记基因探针技术和增强化学发光反应(ECL)技术对扩增产物进行Slot-blot杂交,新建立了PCR-ECL沙门氏菌检测方法,检测限可达10 cfu/g,是一种有前途的快速检测食品中沙门氏菌的方法。章新生[13]等根据寡核苷酸序列合成一对引物用于沙门氏菌的检测,并对其特异性和扩增产物进行了验证。黄金林[14]等根据沙门氏菌组氨酸转运操纵子基因序列设计引物序列,成功扩增出495 bp沙门氏菌特异条带,而阴性对照则无此条带,从而建立了应用PCR进行沙门氏菌快速检测的方法。应用该方法对沙门氏菌属A-F各群中共15株沙门氏菌标准菌株、27株沙门氏菌现场分离菌株和其他10株非沙门氏菌菌株进行PCR扩增,结果沙门氏菌均显示出495 bp特异性DNA扩增条带,而所有对照均不出现特异条带,因此,此方法具有很强的沙门氏菌属特异性。PCR敏感性试验显示,该体系能检出14 pg以上的沙门氏菌DNA。汪琦等[15]利用PCR方法快速检测食品中的沙门氏菌。以全脂奶粉、生牛肉和加工过的鸡肉为实验对象,人为添加沙门氏菌,检测结果与传统培养方法系统检测结果一致。检出限为 100 cfu/25 g,准确性达 100%。
2.2.3 基因芯片技术
Chizhikov等采用寡核苷酸芯片研究沙门氏菌抗原和毒力因子与其致病性的关系,发现细菌毒力因子可用于肠道致病菌的分析检测。靳连群等利用基因芯片技术检测肠道中沙门氏菌等致病菌。结果显示制备的基因芯片能够检测出致病菌,对于未知菌落利用基因芯片能够在3 h完成对未知菌属的鉴定。
2.2.4 环介导等温扩增技术
环介导等温扩增(loop-mediated isothermal amplification,LAMP) 是一种在等温条件下高特异、高效、快速地扩增靶序列的DNA扩增新技术。以沙门氏菌为研究对象,根据其特异性的invA基因,设计了一套特异性引物对该基因进行了LAMP,同时优化了其反应条件,建立了沙门氏菌的LAMP快速检测技术。结果表明,LAMP的最佳反应条件为外引物浓度5 pmol/L、内引物浓 度 40 pmol/L,Mg2+浓 度 6 mmol/L,dNTP 浓度 0.8 mmol/L,甜菜碱浓度0.8 mmol/L,Bst DNA 聚合酶 8U,反应温度63 ℃,反应时间1 h。在此条件下,LAMP检测沙门氏菌DNA的敏感度达10 fg,且与其他常见的细菌无交叉反应。其对牛奶样品的检出量为102cfu/mL,适合于食品中污染沙门氏菌的快速检测[16-17]。
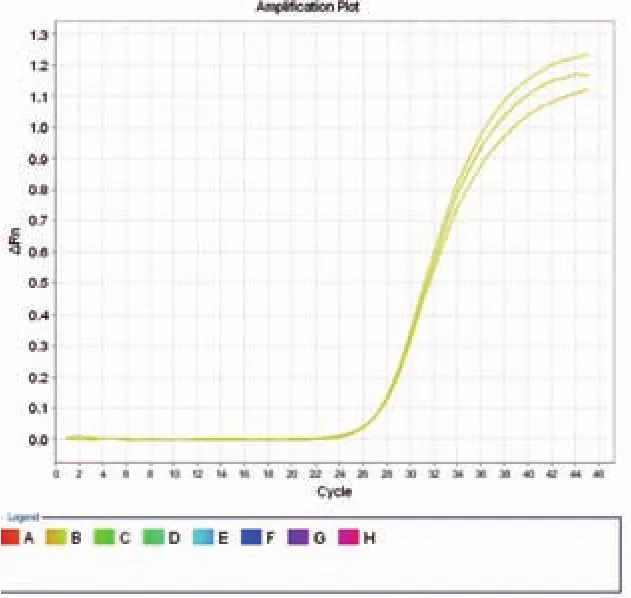
图1 Real-time PCR 有很好的重复性
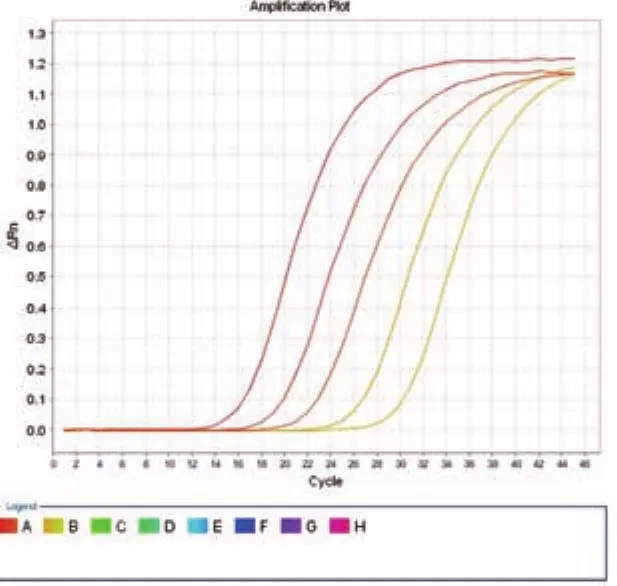
图2 Real-time PCR 特异性及浓度梯度
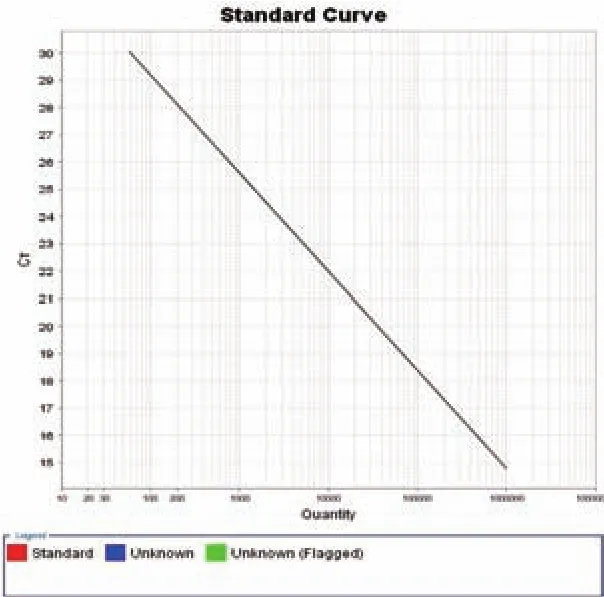
图3 Real-time PCR 所制的标准曲线
2.2.5 实时定量 PCR
实时荧光定量PCR(Realtime PCR) 技 术 于 1996 年 由 美 国Applied Biosystems公司推出,由于该技术不仅实现了PCR从定性到定量的飞跃,而且与常规PCR相比,它具有特异性更强、有效解决PCR污染问题、自动化程度高等特点,目前已得到广泛应用。TaqMan荧光探针:PCR扩增时在加入一对引物的同时加入一个特异性的荧光探针,该探针为一寡核苷酸,两端分别标记一个报告荧光基团和一个淬灭荧光基团。探针完整时,报告基团发射的荧光信号被淬灭基团吸收;PCR扩增时,Taq酶的5′-3′外切酶活性将探针酶切降解,使报告荧光基团和淬灭荧光基团分离,从而荧光监测系统可接收到荧光信号,即每扩增一条DNA链,就有一个荧光分子形成,实现了荧光信号的累积与PCR产物形成完全同步。而新型TaqMan-MGB探针使该技术既可进行基因定量分析,又可分析基因突变(SNP),有望成为基因诊断和个体化用药分析的首选技术平台。现在作为检测的基因有invA、stn、tyv、prt、viaB、flic-d/ flic-a、bipA 和 fimA。理论上只要有1个沙门氏菌就能检测出,但在实践中,一般能检测出5个沙门氏菌[18]。
磁珠富集与实时定量PCR,利用抗体标记的磁珠对沙门氏菌富集,进一步提高灵敏度。
3 其他检测方法
3.1 电阻抗法
阻抗法是几种方法中最少受到主观因素干扰的方法。我国陈广全等[19]用电阻抗法检测食品中的沙门氏菌。经与常规培养法进行比较,对加入食品中的21种已知沙门氏菌属检测,电阻抗法19个为阳性,检出率在90%以上,检测结果与常规培养法一致,对于阴性结果能在48 h内得出。实验结果表明电阻抗法能够快速、可靠地检测食品中沙门氏菌。
3.2 沙门氏菌毒素快速检验法
本法是根据沙门氏菌等肠科杆菌具有产生内毒素的特点,采用氧化呈色反应进行的快速检验。在被检肉浸液中,若含内毒素,加入的硝酸银使毒素氧化成氧化型毒素,氧化型毒素和加入肉浸液中的美蓝牢固结合,当再向肉浸液中加入高锰酸钾时,美蓝不与高锰酸钾作用,使肉浸液呈蓝绿色。
3.3 4-甲基伞形酮辛酯检测法
4-甲基伞形酮辛酯(MUCAP)检测法是利用沙门氏菌具有其他各属细菌都不具备的产生辛酯酶的特性,选用相应的底物和指示剂,将其配制在相关的培养基中,根据沙门氏菌反应后出现的明显颜色变化,而确定待检可疑菌株[20]。
3.4 葡萄球菌A蛋白协凝试验法
Staphylococcal Protein A 简称 SPA即葡萄球菌A蛋白。目前,国外已将SPA用于沙门氏菌的快速检验,所用的葡萄球菌菌株多为CowanⅠ株。国内应用该法检验食品中沙门氏菌报道很少,曹同雪等采用SPA协凝试验法检测100个肉样中的沙门氏菌,结果表明该法快速、准确、灵敏度高、检出率高并且节省费用[21]。
3.5 生物传感器
使用集成化手持式SpreetaTM SPR传感器快速检测沙门氏菌,利用亲和素-生物素系统保证了检测的准确性;同时利用沙门氏菌抗体的免疫吸附反应,保证了结果的特异性;并引入复合抗体作为第二抗体以扩大检测的响应信号,检测到鼠伤寒沙门氏菌的浓度为105cfu/mL,整个检测过程在1 h内完成,从一定程度上实现了食品中病原微生物的快速检测[22]。
沙门氏菌作为食品卫生检验一项重要目标菌,其检测方法的研究有着重要意义。提高沙门氏菌检测灵敏度,缩短检测时间、简化检测程序,使沙门氏菌检测方法向自动化、快速化、灵敏度高、特异性强、重复性好、简易、经济等方向不断发展。随着社会发展与科技进步,新的微生物检验方法和手段不断涌现,这些检测方法的检测结果与传统检测方法大体一致,能够快速、可靠的检测出食品中的沙门氏菌。
[1] 张玉臣,闫文菊.沙门氏菌食物中毒[J].社区医学杂志,2006(12):10-11.
[2] 文其乙,焦新安.直接ELISA检测沙门氏菌方法的建立及其应用研究[J].中国兽医学报,1995,15(2):105-111.
[3] 殷月兰,潘志明,焦新安,等.单抗竞争ELISA检测猪沙门氏菌感染方法的建立与应用[J].中国预防兽医学报,2003(1):65-68.
[4] 黄素珍,杜元钊,张艳红,等.检测肠炎沙门氏菌ELISA方法的建立与应用研究 [J].中国预防兽医学报,2006,28(2):197-200.
[5] 张红见,韩志辉,魏建华,等.Dot-ELISA检测猪胴体中沙门氏菌的研究[J].黑龙江畜牧兽医,2003(11):8-9.
[6] 康明,韩志辉,李刚,等.Dot-ELISA法检测鱼粉中沙门氏菌的研究[J].黑龙江畜牧兽医,2004(7):58-59.
[7] Mansfield L P,Forsythe S J.The detecti on of Salmonella serovars from animal fee d and raw chicken using a combined immunomagnetic separation and ELISA meth od[J].Food Microbiology,2001(18):361-366.
[8] 陶军,张树宏,吴仲梁.自动荧光酶标免疫测试仪与常规培养法对冻禽肉中沙门氏菌的检测效果的比较[J].现代科学仪器,2001(3):50-53.
[9] 黄玲,孟冬丽.利用mini-VIDAS和GB方法检测食品中沙门氏菌的比较试验[J].新疆师范大学学报自然科学版,2003,22(1):50-52.
[10] 陈炜,王德军,马巧兰,等.脱水蔬菜中沙门氏菌不同检测方法的对比研究[J].宁夏农林科技,2003(1):24-25.
[11] Torpdahl M , Sorensen G ,Lindstedt B-A,et al.Tandem repeat analysis for surveillance of human S almonella Typhimurium infections[J].Emerg Infect Dis,2007,13 :388-395.
[12] 卢强,陈贵连,林万明.应用PCRECL技术检测食品中的沙门氏菌[J].中国公共卫生,1995,11(4):189-193.
[13] 章新生,臧富妍.应用PCR技术检测沙门氏菌[J].中国兽医学报,1999,19(2):147-151.
[14] 黄金林,焦新安,潘志明,等.应用聚合酶链反应快速检测沙门氏菌[J].扬州大学学报:农业与生命科学版,2002,2(3):5-11.
[15] 汪琦,张昕,张惠媛,等.利用PCR方法快速检测食品中的沙门氏菌[J].检验检疫科学,2005,15(6):26-28.
[16] 朱胜梅,吴佳佳,徐驰,等.环介导等温扩增技术快速检测沙门菌[J].现代食品科技,2008,24(7):725-730.
[17] Cook N.The use of NASBA for the detection of microbial pathogens in food and environmental samples[J].J Appl Microbiol,2003,53 :165-174.
[18] Malorny B , Paccassoni E ,Fach P, et al. Diagnostic realtime PCR of Salmonella in food[J].Appl Environ Microbiol,2004,70 :7046-7052.
[19] 陈广全,张惠嫒,饶红,等.电阻抗法检测食品中沙门氏菌[J].食品科学,2001,22(9):66-70.
[20] 褚天新,苏丽霞,路文彬.MUCAP荧光试验快速检测沙门氏菌[J].中国公共卫生,2000(1):69-70.
[21] 曹同雪,唐中令,石如彦,等.SPA协凝试验法快速检验肉中沙门氏菌[J].肉品卫生,1997(6):1-5.
[22] 王凯,殷涌光.SPR生物传感器快速检测沙门氏菌研究初探[J].食品科技,2007,(9):191-95.
猜你喜欢
中国食品学报(2022年6期)2022-07-19
中国饲料(2021年17期)2021-11-02
食品安全导刊(2021年20期)2021-08-30
现代临床医学(2021年4期)2021-07-31
中国现代医药杂志(2020年10期)2020-12-14
食品安全导刊(2019年30期)2019-01-05
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
中国医药生物技术(2015年4期)2015-12-26
特产研究(2015年1期)2015-04-12
中国当代医药(2015年32期)2015-03-01
