
机械蝴蝶模型悬停飞行的流动显示实验
2010-04-15 10:55黄烁桥申功炘
实验流体力学 2010年2期
黄烁桥,申功炘,魏 来,郭 鹏
(1.北京航空航天大学流体所,北京 100191;2.北京航空精密机械研究所,北京 100076)
0 引 言
昆虫在低雷诺数环境下具备的卓越飞行本领,是同尺度人造飞行器所难以比拟的。因此,通过流体力学实验探索昆虫飞行的高升力机理,为微型飞行器(MAV)的气动布局设计及控制方式提供新概念,是当今的一个研究热点。
目前已经建立起来的非定常高升力理论[1-5],大都是在对蜂、蝇、蛾、蜻蜓等昆虫的研究中得出的。这些昆虫有很多共同点,如翅膀的展弦比较大,拍翅频率较高(最低的也有几十赫兹),且运动模式相似:翅膀拍动平面接近水平或者与水平方向成一定夹角,整个拍动过程可简化为“平动+翻转”两个部分 每次上拍或下拍的中间阶段翅膀保持迎角不变向前或向后运动,此过程称为“平动”,而上下拍转换时迎角变化的过程则称为“翻转”(“翻转”的过程中“平动”也没有停止)---由于拍动平面近似水平,所以“翻转”保证了产生的前缘涡永远在上翼面,即气动力方向永远向上见图1(a)。
蝴蝶也是一种常见的具备飞行能力的昆虫,它飞行速度不高,飞行轨迹飘忽不定,在稳定性和机动性上,比之蜂、蝇、蛾、蜻蜓等似乎都有不足。但蝴蝶中有许多具有迁徙习性的种类,如美洲帝王蝶等,长途飞行能力堪称昆虫之最。另外,蝴蝶在外形及运动模式方面都很特殊:它的翅膀非常宽大,展弦比很小(约为1.5),且无法实现"翻转"运动,只能在垂直于身体的面内做上下拍动,拍频很低,一般只有10Hz左右;另外,在其拍翅的过程中,伴随着明显的身体俯仰和上下振动[6]______也就是说,蝴蝶不能实现像上述昆虫一样,真正意义上的悬停飞行,只能在一段时间内近似保持平均位置不变见图1(b)。
目前针对蝴蝶飞行的研究还很少,且主要为形态、运动学参数的测量,以及一些初步的定性流动显示,因此,对蝴蝶飞行的气动机理还知之甚少,需要进行细致的流动观测。
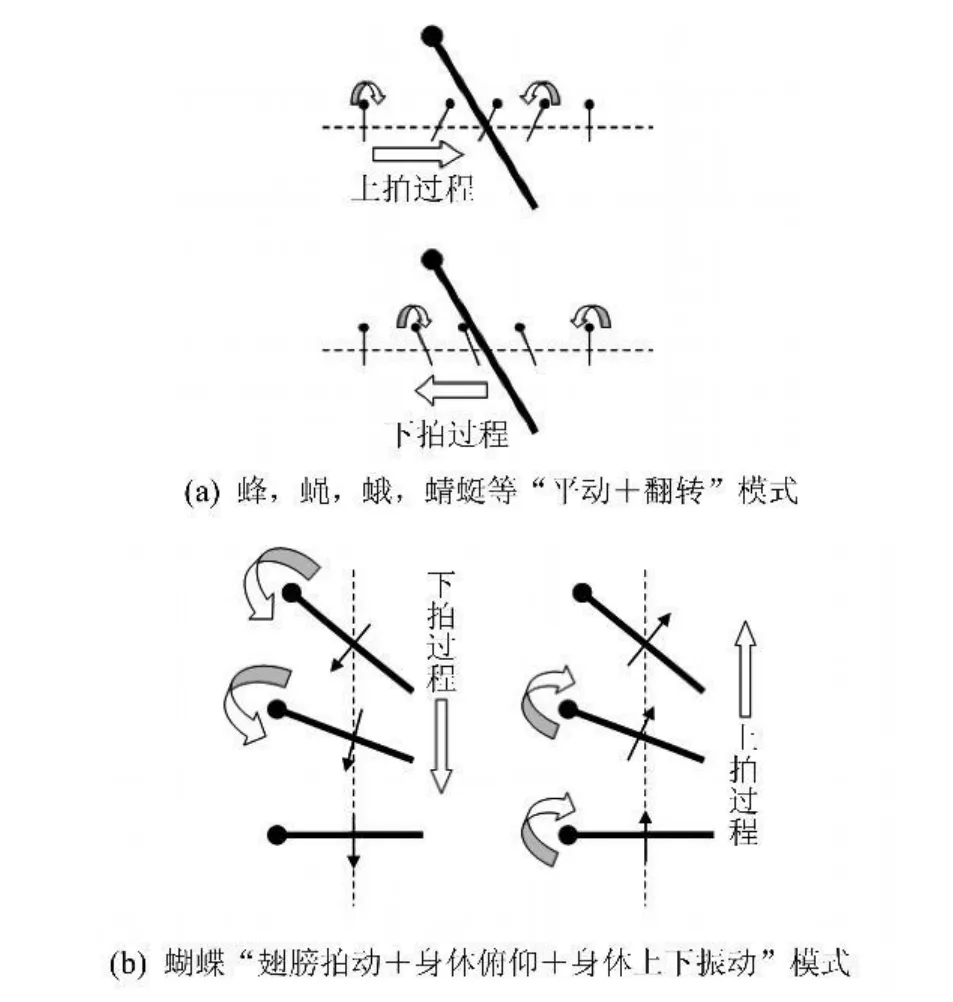
图1 两种运动模式示意简图Fig.1 Sketch of two flapping modes
1 实验方法
1.1 模型
选择了morpho peleides作为模型制作的原型,见图2,这是一种美丽的南美闪蝶,具有非常典型的翅膀形状,各种形态学参数也接近于蝴蝶的平均值,比较有代表性。由于主要目的在于考察蝴蝶独特的运动模式下产生的流动结构,实验中对模型进行了适当简化,首先将位于蝴蝶身体一侧的两个翅膀简化成一个,其次,将翅膀和身体都视为刚体。在确定模型的形态学及运动学参数时,主要参考了黄华,孙茂的计算结果[6]及Dudley的实验测量结果[7,8]。模型的具体外形及相关参数列于表1。
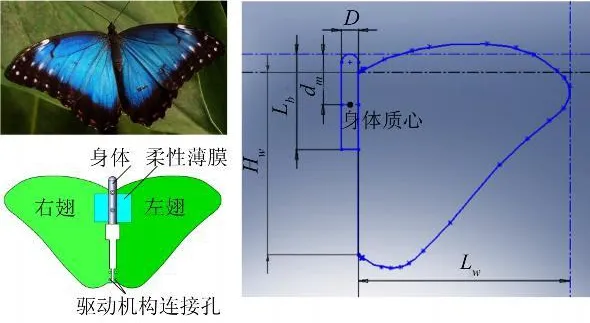
图2 实验模型Fig.2 Experimental model
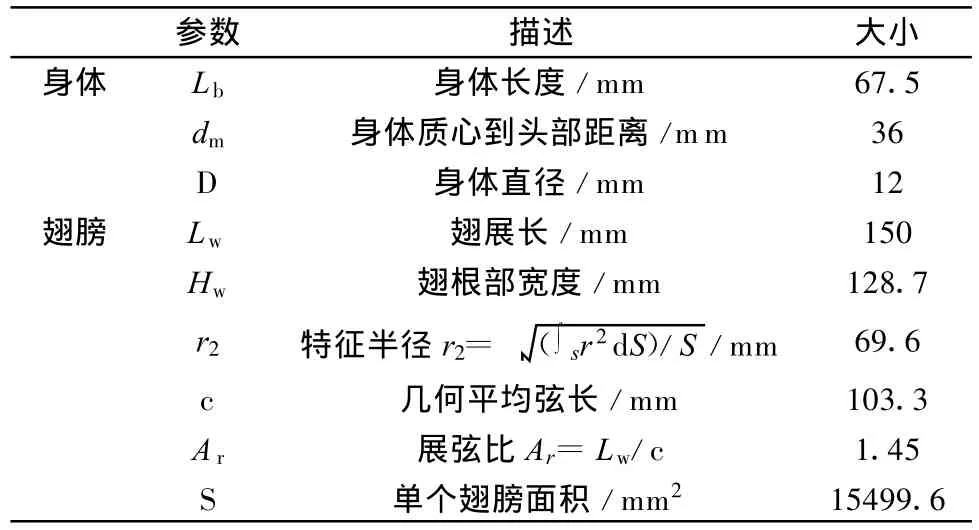
表1 模型外形参数Table 1 Parameters of morphology
1.2 机构及运动函数介绍
如前言中所介绍,蝴蝶的运动模式很特殊,也比较复杂,同时包括了翅膀与身体的运动,即:翅膀的拍动、身体绕质心的俯仰及身体的上下振动。
为了确定这3种运动,在图3中定义了坐标系:以质心点O为坐标原点,OXEYEZE为对地坐标系,OXbYbZb为固定在身体上的坐标系,Xb轴为身体的对称轴且指向头部为正,Yb轴位于身体的水平对称面内且向右为正,Zb轴位于身体的垂直对称面且向下为正。
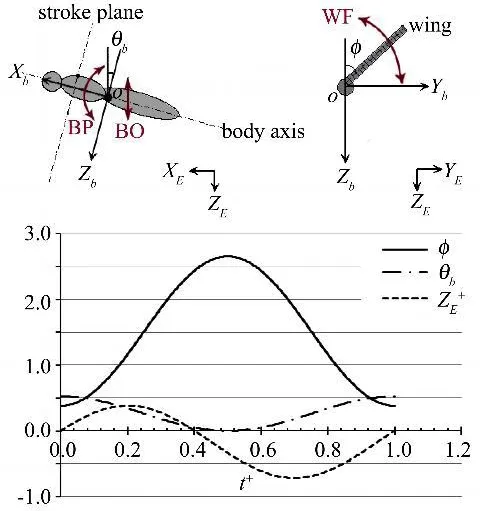
图3 坐标及运动函数定义Fig.3 Definitions of coordinate systems and kinematics functions
翅膀的拍动运动WF(Wing Flapping)由翅膀与身体垂直面(轴Zb所在平面)的夹角φ决定;身体绕质心的转动BP(Body Pitching)由身体轴线与水平面(轴 XE所在平面)的夹角θb定义;而身体的上下振动BO(Body Oscillating)则由竖直面内的移动距离ZE来表示(图4)。运动根据黄华、孙茂的计算结果[6]简化成了正、余弦函数(运动曲线见图3)。

图4 运动机构简图Fig.4 Sketch of mechanical driving system
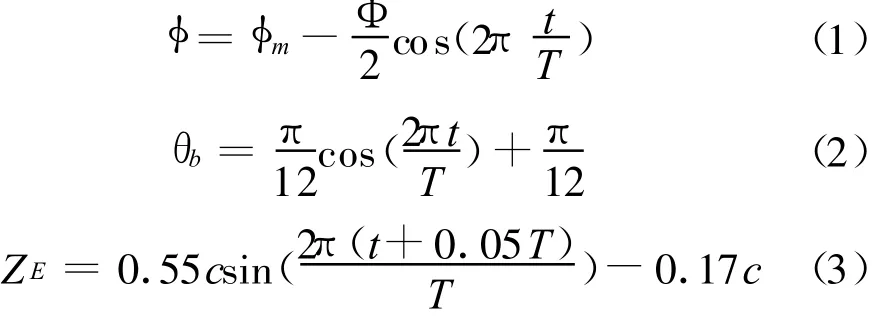

表2 运动参数的定义及取值Table 2 Parameters of kinematics
式中的 φm=87°为拍动平均角 ,Φ=130°为拍动幅度,T=10s为运动周期(3种运动周期相同),而c为几何平均弦长。按此定义,在本实验中,模型的初始状态为身体轴线与水平面呈30°角(即θb=30°),两翅夹角(上表面)为 44°(即 φ=22°),而下拍结束时(0.5T)体轴与水平面夹角变为0°,两翅夹角(下表面)变为 56°(即 φ=152°)。运动参数的具体定义及取值列于表2,其中雷诺数Re=3264,在蝴蝶的范围(1000~5000)内;斯特劳哈数St=An/U中,A=312mm为非定常尾迹在拍动平面上的流场宽度,用蝴蝶拍动时双翅尖端达到的最远距离(即双翅夹角呈180°时)近似代替,n=0.1Hz为翅膀拍动频率,U=31.6mm/s为翅膀特征半径处的拍动线速度。
实际实验中,模型被头朝下安装,如图4所示(坐标定义与图3对应)。值得一提的是,绕身体质心的俯仰实际上为一个绕空间虚点的运动,通过绕点O'的转动及两个电移台的补偿运动拟合实现。
1.3 染色液流动显示系统
如图5所示,实验在一个无来流的水缸中进行(长宽高:1200mm×1000mm×950mm);染色液容器被固定在水缸侧上方的一个可垂直升降的吊架上,可通过调节其高度来控制染色液的流出速度;染色液输入管沿着两根驱动杆进入左右翅膀的后缘,尽可能地减小了对流场的扰动。流动显示系统的采集设备为两台日本JVC的GZ-MG255摄像机,拍摄时采用了PAL制式标准,即每秒25帧图像,每帧图像分辨率为720×576。

图5 染色液流动显示系统布局Fig.5 Experiment setup
1.4 实验内容
实验中对蝴蝶的3个基本运动:翅膀拍动(F)、身体绕质心的俯仰(P),及身体上下振动(O),进行了不同的组合研究,以翅膀拍动运动(F)为基础,得到了4种不同的拍动模式,分别为:‘F'模式(Flapping),即只有翅膀拍动的运动模式;‘FP'模式(Flapping&Pitching),翅膀拍动加上身体绕质心的俯仰运动;‘FO'模式(Flapping&Oscillating),翅膀拍动的同时身体做上下振动。在振动中过程中,模型体轴始终和水平面保持 15°(俯仰平均角);‘All'模式(Flapping&Pitching&Oscillating),即3个动作都包括的运动模式--此模式最接近于蝴蝶的真实飞行状态。
2 实验结果及分析
2.1 ‘F'模式
图6给出了‘F'模式下的流动显示结果。可以看到,只存在翅膀拍动时,上、下翼面分别在下拍和上拍的过程中产生了前缘涡。与以往在拍动翼实验中观测到的从翼根部发出、螺旋或锥状的前缘涡不同,这里产生的前缘涡为横跨在身体及左右两翅之上的单个联通结构,且下翼面在上拍过程中形成的“联通”较上翼面在下拍过程中形成的弱。这种现象类似于Weis-Fogh的“合拢-打开”(clap-and-fling)效应,但又有所区别——拍动之初,双翅并不是合拢在一起而是存在一定夹角(下拍时上翼面起始夹角为44°,上拍时下翼面的起始夹角为56°),当双翅从一个小的夹角加速向两边打开时,会在两翅中间形成一个低压区,空气绕过头部、翅缘涌入这个低压区时就形成了联通的前缘涡——两翅的初始夹角越小,这种"打开"的效应就越强,形成的前缘涡也就越强。另外,在拍动加速结束时(下拍时0.25T,及上拍时0.75T),前缘涡出现了破裂,而在此之前,前缘涡有一个明显的展向发展过程。
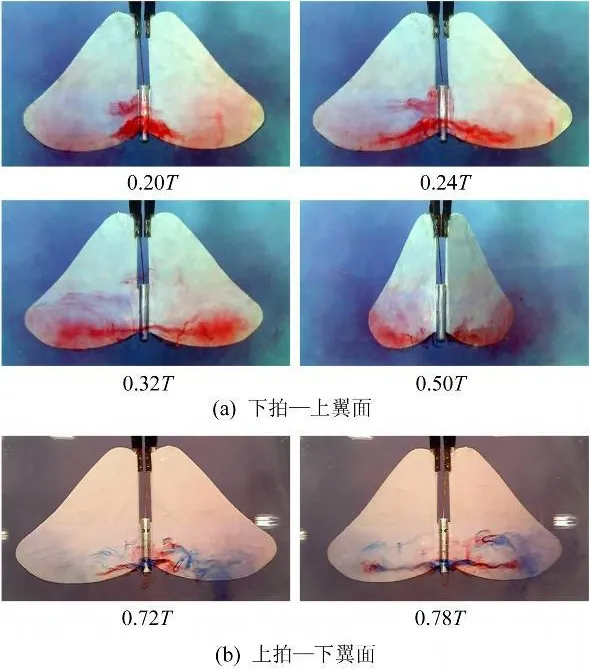
图6 拍动模式‘F'Fig.6 Flapping mode‘F'
2.2 ‘FP'模式
从图7的流动显示结果中可看到,在‘FP'模式中,下拍时前缘涡的总体发展过程与‘F'模式相似,经历了“形成联通涡→涡沿展向发展→涡出现破裂→破裂向翼根部扩散”这样一个发展变化的过程,但在下拍及下俯达到最大速度的时候(0.25T)出现了一个大的前缘联通涡(Lp)及两个从翼根部发出的小的锥状前缘涡(Lm-s),其中的锥状次涡类似于陆远观测到的结构[9]。但在上拍过程中,两侧翅膀上分别生成了与传统锥状涡相似的前缘涡,没能在身体上方形成“联通”,这可能是由于身体的俯仰运动使身体头部相对于翅膀拍动引起的来流迎角减小导致的。
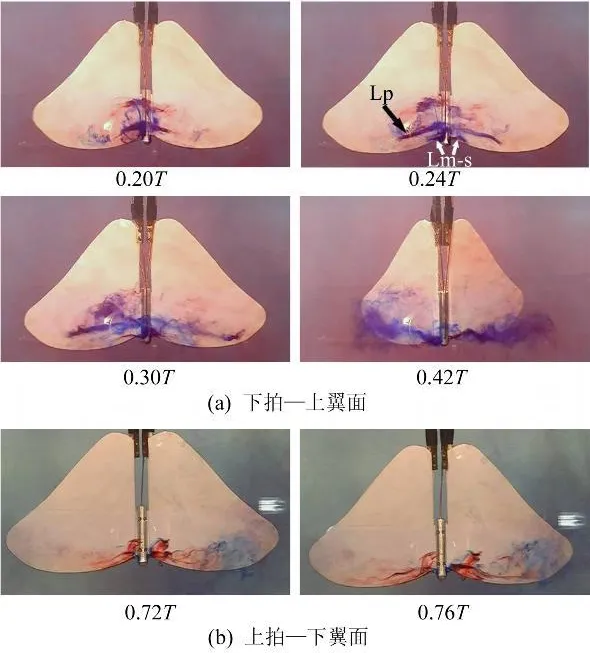
图7 拍动模式‘FP'Fig.7 Flapping mode‘FP'
2.3 ‘FO'模式
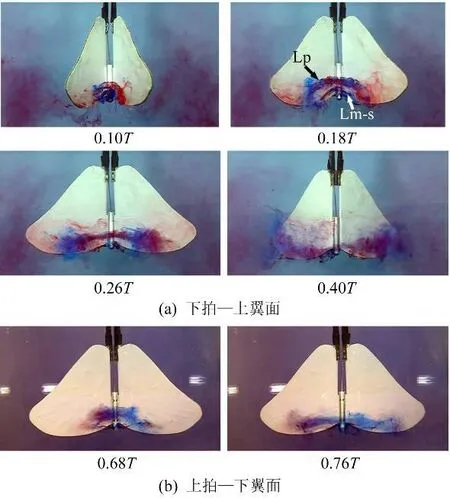
图8 拍动模式‘FO'Fig.8 Flapping mode‘FO'
图8显示了前缘涡在拍动模式‘FO'中的发展变化过程,可见,上下拍时都形成了跨越身体的联通结构,且涡的尺寸及在弦向(Xb轴方向)上的发展范围较之‘F'、‘FP'两种模式都有显著增加;而且在下拍加速、且身体向下运动的阶段同时出现了两个呈倒扣的‘U'状、且相互平行联通涡--类似于Tomas等人观测到的蝴蝶在需要大的加速度时产生的结构[10],此情况下产生的升力应该较单前缘涡时大,即身体振动运动对前缘涡起到了加强的作用。
2.4 ‘All'模式
从图9中可以看到,拍动模式‘All'中前缘涡的发展情况与‘FO'模式中类似,上下拍时都形成了跨越身体的联通结构,但涡在弦向(Xb轴方向)上的发展范围较之‘FO'更大,在下拍加速、且身体向下运动的过程中最多同时出现了3个联通涡(见图9(a),0.18T)。而且产生的涡几乎在整个下拍过程中都附着于翼面。
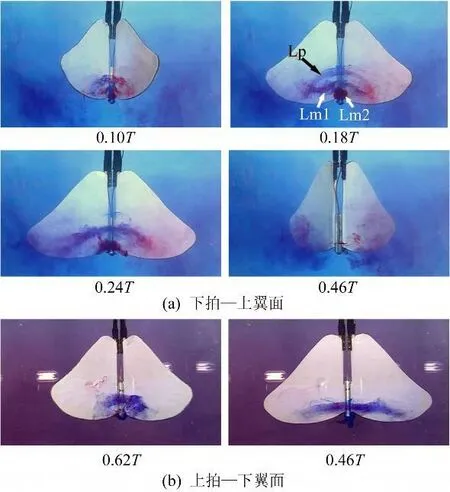
图9 拍动模式‘All'Fig.9 Flapping mode‘All'
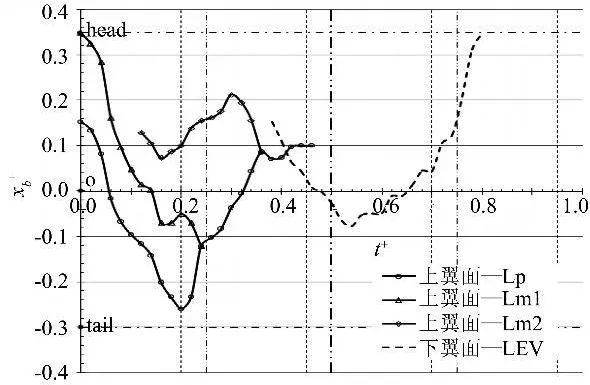
图10 拍动模式‘All'下的前缘涡在身体上方的弦向位置变化Fig.10 The chordwise position of LEV over the body in‘All'mode
另外,涡在弦向上运动的范围在某种意义上可以作为衡量其从前缘卷起强度的另一指标。图10统计了拍动模式‘All'下产生前缘涡在身体轴线(Xb轴)上的位置变化情况。其中横坐标t+=t/T,为无量纲时间;纵坐标=xb/c(c为翼型的平均气动弦长),因为原点O位于身体质心,所以xb表征的是到质心的距离。
从图中可看到,在‘All'模式中,前缘涡在弦向(轴Xb向)上的运动一度越过了质心。可以想象:在下拍过程中,当前缘涡(升力作用点)位于质心之前(即在头部和质心之间)时会产生一个抬头力矩,而若其位于质心之后(质心与尾部之间),则会产生一个低头力矩;上拍时与下拍的情况刚好相反。基于这个结论再对图10进行观察,发现由前缘涡在弦向运动导致的身体俯仰运动趋势与实验设定的俯仰函数(0~0.25T:下俯加速;0.25~0.5T:下俯减速;0.5~0.75T:上仰加速;0.75T~1T:上仰减速)基本一致:当下拍开始时,身体首先会获得一个瞬时的下俯加速度(可能由打开时两侧翼尖涡形成的射流导致);0.05~0.3T时上翼面上的前缘涡运动到质心之后,产生的低头力矩使身体的下俯运动继续加速;0.3~0.5T时上翼面上的前缘涡运动回质心之前,而后,0.5~0.6T时下翼面的前缘涡运动到质心之后,这两个过程产生了一个持续的抬头力矩,对应着身体的下俯减速及接下来的上仰加速;0.6~1T期间下翼面上的前缘涡位于质心之前,产生的低头力矩使身体上仰减速。由此我们可以大胆推测:身体的俯仰运动可能并不像翅膀拍动一样,是一种主动控制运动;而是蝴蝶在飞行过程中,随着前缘涡位置变化产生的一种自适应的运动,它能够增强前缘涡并使其更靠近身体,从而增加升力;另外,蝴蝶可以通过曲卷身体等方式改变质心位置,从而对俯仰运动进行间接控制。值得一提的是,东京大学的 Hiroto Tanaka等人曾对其自制的蝴蝶仿生飞行器进行了放飞拍摄,发现其在只驱动翅膀拍动这一种运动的情况下仍会在飞行过程中发生身体俯仰角度的变化[11],这一结果也可作为本推论的一个佐证。
还根据染色液实验中拍摄到的图像进行了统计分析,将各种模式中上、下拍产生的前缘涡发展情况进行了量化,结果列于表3。当然,染色液主要的功能是定性,由于无法去除染色液的释放速度等可能造成误差的原因,表3中得到的统计数据仅能作为反映流动趋势的一个参考。
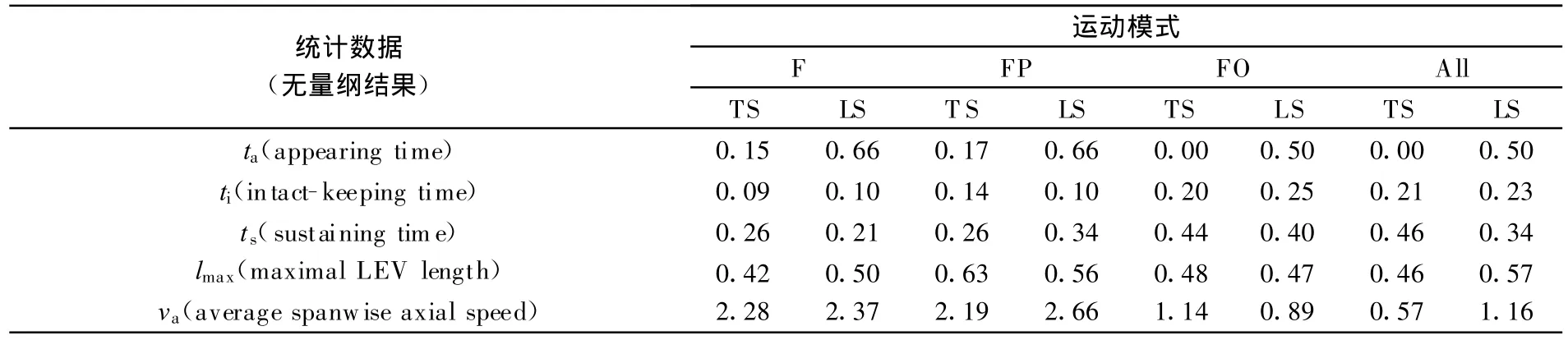
表3 4种拍动模式下的前缘涡发展情况统计(多前缘涡的情况只考察主涡)Table 3 Development of leading edge vortex at four flqpping modes
3 结 论
(1)在蝴蝶的飞行过程中(拍动模式‘All'),前缘涡以跨越身体的柱状联通结构出现,而不是传统的螺旋或锥状结构--在下拍的0~0.2T(拍动加速且身体向下运动)期间,会出现多前缘涡结构;其显著特征为:拍动加速阶段(0~0.25T和 0.5~0.75T):具有明显的展向速度;而拍动开始减速后(0.25T~0.5T和0.75T~1T)会出现破裂;
(2)翅膀拍动(F)导致的"打开"效应,是出现联通结构的关键;而身体俯仰运动(P)能使前缘涡形成的位置更靠近翼面及质心,身体上下振动(O)则能对涡起到增强作用,皆有利于增大升力;
(3)下、上拍过程中,前缘涡分别形成于上、下翼面--这种气动力周期性的换向,正是导致蝴蝶在飞行时上下振动的原因;
(4)通过统计前缘涡在身体弦向上的运动位置,发现身体俯仰很可能是一种自适应运动,但可以通过在飞行中改变质心位置进行运动调节。
[1]WEIS-Fogh T.Quick estimates of flight fitness in hovering animals,including novel mechanisms for lift production[J].J Exp Biol,1973,59:169-230.
[2]ELLINGTON C P,VAN Den Berg C,WILLMOT T A P,et al.Leading edge vortices in insect flight[J].Nature.1996,384:626-630.
[3]VAN Den Birg C,ELLINGTON C P.The three-dimensional leading edge vortex of a‘hovering'model howkmoth[J].Phil Trans R Soc Lond B,1997,352:329-340.
[4]DICKINSON M H,LEHMANN F O,SANE S.Wing rotation and the aerodynamic basis of insect flight[J].Science,1999,284:1954-1960.
[5]SUN M,HAM ADANI H.High-lift generation by an airfoil performing unsteady motion at low Reynolds number[J].Acta Mech Sinica,2001,17(2):1-18.
[6]黄华.蝴蝶飞行的气动力及能耗特性研究[D].北京航空航天大学,2007.
[7]DUDLEY R.Biomechanics of flight in neotropical butterflies:Morphometrics and kinematics[J].1990,Journal of Experimental Biology,150:37-53.
[8]DUDLEY R.Biomechanics of flight in neotropical butterflies:Aerodynamics and mechanical power requirements[J].1991,Journal of Experimental Biology,159:335-357.
[9]LU Y,SHEN G X,LAI G J.Dual leading-edge vortices on flapping wings[J].Journal of Experimental Biology 2006.209:5005-5016.
[10]SRYGLEY R B,THOM AS A L R.Unconventional liftgenerating mechanisms in free-flying butterflies[J].2002,Nature,420:660-664.
[11]HIROTO TANAKA,KAZUNORI HOSHINO,KIYOSHI MATSUMOTO,at al.Flight dynamics of a butterfly-type orithopter[A].2005 IEEE/RSJ International Conference on Intelligent Robots and Systems[C].Edmonton,Canada,2005:2706-2711.
猜你喜欢
探测与控制学报(2022年4期)2022-08-30
汽车实用技术(2022年14期)2022-07-30
兵器装备工程学报(2021年12期)2022-01-11
北京航空航天大学学报(2021年4期)2021-11-24
民用飞机设计与研究(2020年1期)2020-05-21
中国惯性技术学报(2019年6期)2019-03-04
宝藏(2019年6期)2019-01-15
探测与控制学报(2018年3期)2018-07-09
航空模型(2017年10期)2018-02-23
南方文学(2016年4期)2016-06-12
