
温度和光照时间对3个秋眠型紫花苜蓿品种形态特征的影响
2010-03-31 06:12陈玮玮万里强何峰李向林刘树军
草业科学 2010年12期
陈玮玮,万里强,何峰,李向林,刘树军
(1.兰州大学草地农业科技学院,甘肃兰州 730020;2.中国农业科学院北京畜牧兽医研究所,北京 100094)
紫花苜蓿(Medicago sativa)为豆科多年生植物,以多年生品种最多。紫花苜蓿是一种比较耐盐的多年生豆科牧草,它不仅产量高,耐旱性强,可为家畜养殖提供足够的优质牧草[1],而且具有明显的土壤改良效果[2],成为改良中轻度盐碱荒地的一种重要植物[3-5]。
秋眠性是苜蓿的一种生长特性,是指在较高纬度地区,光照减少和气温下降导致紫花苜蓿形态类型和生产能力发生变化的现象[6-7]。秋季日照时数的减少引起苜蓿生理休眠,植株由向上生长转向匍匐生长,导致总产量减少,它可以发生在任何温度和湿度水平上。卢欣石[8]认为苜蓿秋眠性是一种生长特性,这种特性与苜蓿的耐寒力和生产性能有直接的关系。
紫花苜蓿的形态结构与生境有关,其结构变化与功能的统一是其适应生长环境的生物学基础。生境变化时不同秋眠级苜蓿品种生物学特性和生产性能差异比较大,当日照长度变短或(和)温度变低时,秋眠型品种将被触发,地上部分不生长或极少生长,非秋眠品种则继续旺盛生长[9-10]。有研究表明,当光照减少时,秋眠型苜蓿品种的生长开始矮化,植株停止向上生长,生长趋于匍匐,日照长度越短,植株矮化越明显[11]。卢欣石和申玉龙[12]认为,不同秋眠型苜蓿受光照和温度影响不同,秋眠型品种受光照时数影响较大。此外,秋眠型品种受温度影响远远大于非秋眠型品种。本研究通过模拟不同温度与日照长度,对3个不同秋眠级紫花苜蓿品种的形态特征的变化进行分析,探讨短日照和低温对不同秋眠级紫花苜蓿形态建成的影响,为实际生产中苜蓿的区划种植提供参考。
1 材料与方法
1.1 试验材料3个紫花苜蓿品种为阿尔冈金(Algonqiun)、德宝(Derby)和 CUF101,秋眠级分别为2、5和9级。
1.2 试验处理在温室中,采用统一标准的花盆(上直径 11 cm,下直径 7.5 cm,高9 cm),花盆中装沙质壤土,播种时充分浇水,每个品种播种40盆,每盆播种7~8粒。幼苗期间,每盆留生长健壮植株3~5株,并进行正常的浇水、除草、防病虫。幼苗在温室生长40 d后,每个品种挑选生长健壮、长势一致的植株16盆,3个品种共计48盆进行进一步培养。
将48盆紫花苜蓿分别放入4个光照培养箱(均匀光照,光照强度为24 000 lx,湿度为60%),每个品种4次重复,光照培养箱设置4个不同的光长(16、14、10和 8 h)和5 个温度条件(30、25、20、15、10和5℃)。其中,对照中光照时数和温度保持不变(白天和黑夜光照均为12 h,温度分别为25和20℃),其余3个光照培养箱在不同周期分别进行光照时数和昼夜温度的变化,每个周期7 d。人工气候室实验条件见表1。
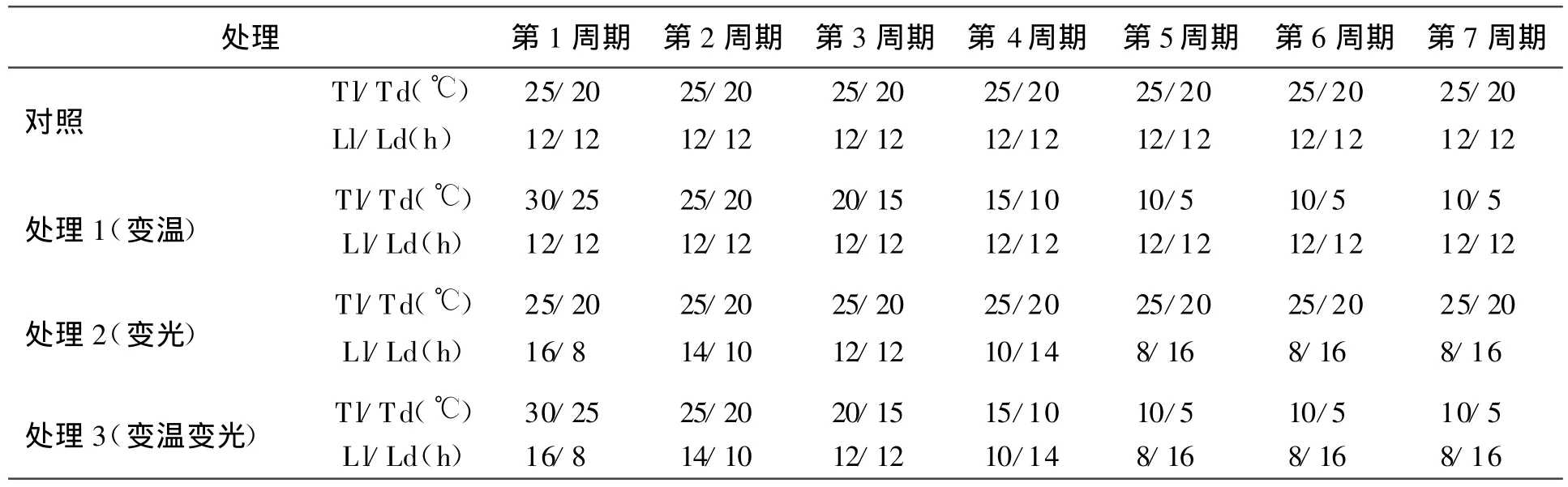
表1 人工气候室实验条件
1.3 测定指标及方法放入光照培养箱开始测量,记为第1天,每隔4 d进行测量。
茎粗:选取10株健康植株,测定基部以上的最大直径处,用游标卡尺测量,精确到0.001 cm。
叶片长宽:选择3小叶中间叶片叶梗到叶尖处的距离为叶片长,叶片最宽处距离为叶片宽,精确到0.1 cm。
直立性:植株直立性用植株自然株高和绝对株高的比值来表示。每个培养箱每个品种选取生长健壮的植株10株,用卷尺测量自然株高和绝对株高,精确到0.1 cm。
节间长:每个培养箱每个品种选取10株生长旺盛的植株进行测量,测定从根冠到顶端主茎长度内的茎节数目。

茎叶比:第7周期结束后(即第49天)把同一培养箱中同一品种植株齐地完整剪下,快速将叶片和茎秆分离,分别用天平称量(花蕾也算为叶片部分)。称鲜质量后,叶片和茎秆分别在80℃下烘干至质量恒定,称量。

以上指标测定从苜蓿放入人工气候室开始,测量、记录采集数据,直至第7周期结束。
1.4 数据统计分析实验数据采用SPSS 13.0统计软件one-way ANOVA过程进行方差分析,Excel 2003进行图形绘制等工作。
2 结果与分析
2.1 不同秋眠型品种分枝数的变化在对照、降温处理和缩短光长处理中,阿尔冈金和德宝分枝数在第17天以前均缓慢增加,从第25天开始3个品种分枝数明显增加(表2)。对照处理中,阿尔冈金分枝数最多,德宝次之,CUF101最少,这说明适宜的光温条件更有利于秋眠型品种分枝的生长。在降温处理中,各个品种分枝数呈现出一致的变化趋势。缩短光长条件下,在第25天以前,3个品种分枝数变化趋势一致,第25天后,随着光长缩短阿尔冈金和德宝分枝数变化一致,和对照相比差异均不显著(P>0.05)。从表2可以看出,当降温和光长缩短互作时,阿尔冈金分枝数最多,德宝分枝数最少,从第25天开始3个品种分枝数显著(P<0.05)少于对照。
2.2 不同秋眠型品种茎粗增长量、平均茎节长和直立性的变化对照中3个紫花苜蓿品种茎粗增长量维持在一个稳定的水平上,其中CUF101茎粗增长量值最大,德宝和阿尔冈金的茎粗增长量基本一致。当温度降低或光长缩短时,3个品种茎粗变化趋势相同,第33天之前呈增加趋势,第33天以后呈下降趋势;第33天以后,经过变温或变光处理的各个品种的茎粗增长量均显著低于(P<0.05)对照,在第49天,除CUF101的缩短光长处理外,3个品种的茎粗增长值无显著差异(P>0.05)。当降温和光长缩短互作时3个品种茎粗波动较大,第25天以后,3个品种茎粗变化趋势基本相同,阿尔冈金的茎粗最小。
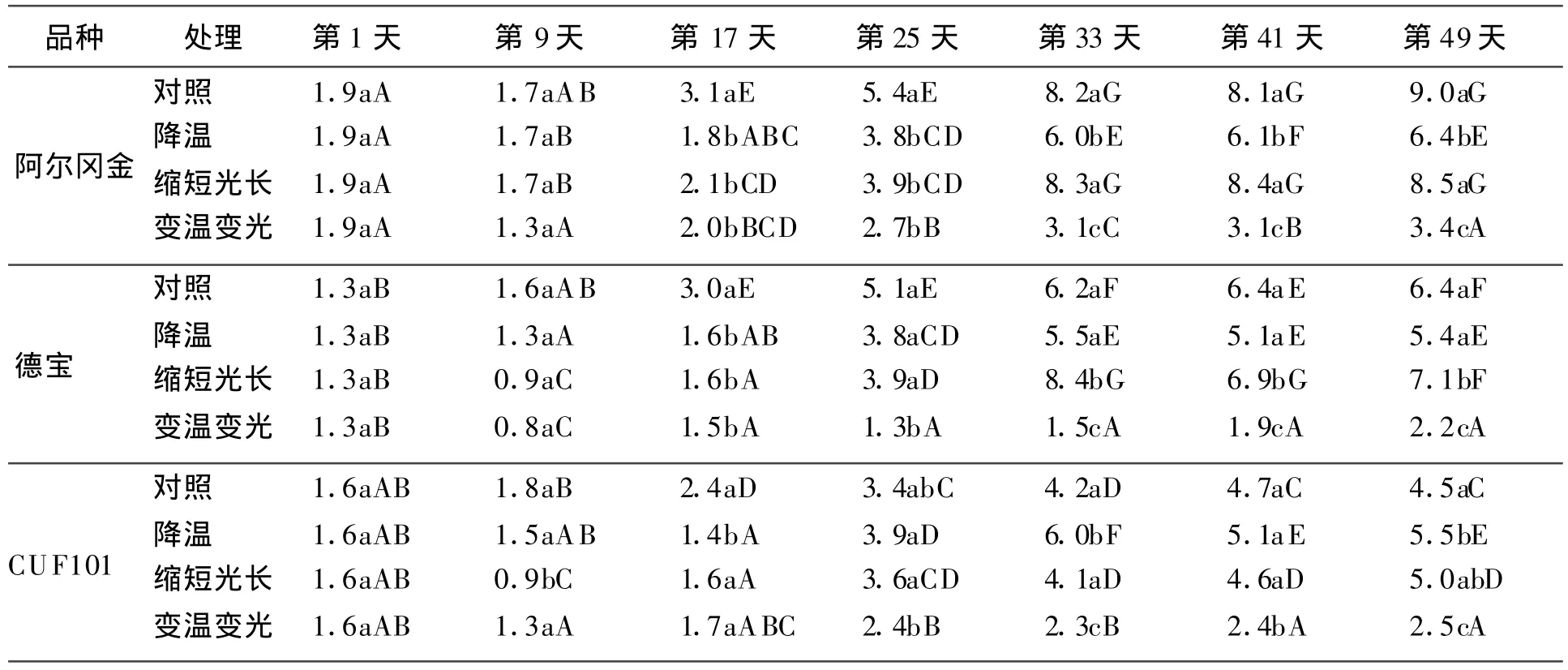
表2 低温和短日照对分枝数的影响
在对照中,3个紫花苜蓿品种平均节间都缓慢伸长,在光温适宜条件下,阿尔冈金的节间长最长。当降温或光长缩短时,3个品种平均节间长都是先缓慢增加,第33天以后快速降低,说明不同秋眠型紫花苜蓿在光长缩短或降温时茎节都呈缩短趋势;当温度降低时,阿尔冈金和德宝的平均节间长变化一致,CUF101的平均节间长变化幅度最大,从第33天开始明显低于其他2个品种的平均节间长;在整个观察过程中,阿尔冈金对光长缩短反应最为敏感,变化幅度最大,CUF101受光长影响节间长变化幅度最小。当降温和光长缩短同时作用于植株时,3个品种变化趋势一致,阿尔冈金的平均茎节长低于其他 2个品种(P>0.05)。

表3 低温和短日照对茎粗增长量的影响 cm
对照中,3个品种直立性变化曲线一致,并且都维持在一个较高的水平上,波动范围很小。温度降低时CUF101直立性最小,匍匐生长的趋势明显大于阿尔冈金和德宝,从第 25天开始CUF101直立性显著低于对照(P<0.05),阿尔冈金和德宝直立性和对照差异不显著(P>0.05)。在光长缩短影响下3个品种直立性都呈下降趋势,第33天以后显著低于对照(P<0.05);从第33天开始,阿尔冈金直立性显著低于其他2个品种,德宝和CUF101直立性基本相同。当降温和光长缩短互作时3个品种植株直立性变化一致,都是直立性迅速降低,显著低于对照和其他2个处理(P<0.05),从第25天开始,3个品种直立性都维持在同一个水平。
2.3 不同秋眠型品种叶长/叶宽、茎叶比和地上生物量/地下生物量的变化在光温适宜条件下,3个品种叶长叶宽比保持在一个稳定的水平,阿尔冈金的叶长/叶宽最大。当温度、光长中1个或2个因素降低(缩短)时,3个品种叶长/叶宽都呈现基本一致的增长趋势,即第25天以前缓慢增大,第25天至第41天快速增大,第41天以后缓慢增大,最终维持在一定的水平;当降温或光长缩短作用于植株时,阿尔冈金的叶长/叶宽比值最大,德宝的比值最小。

表4 低温和短日照对节间长的影响 cm
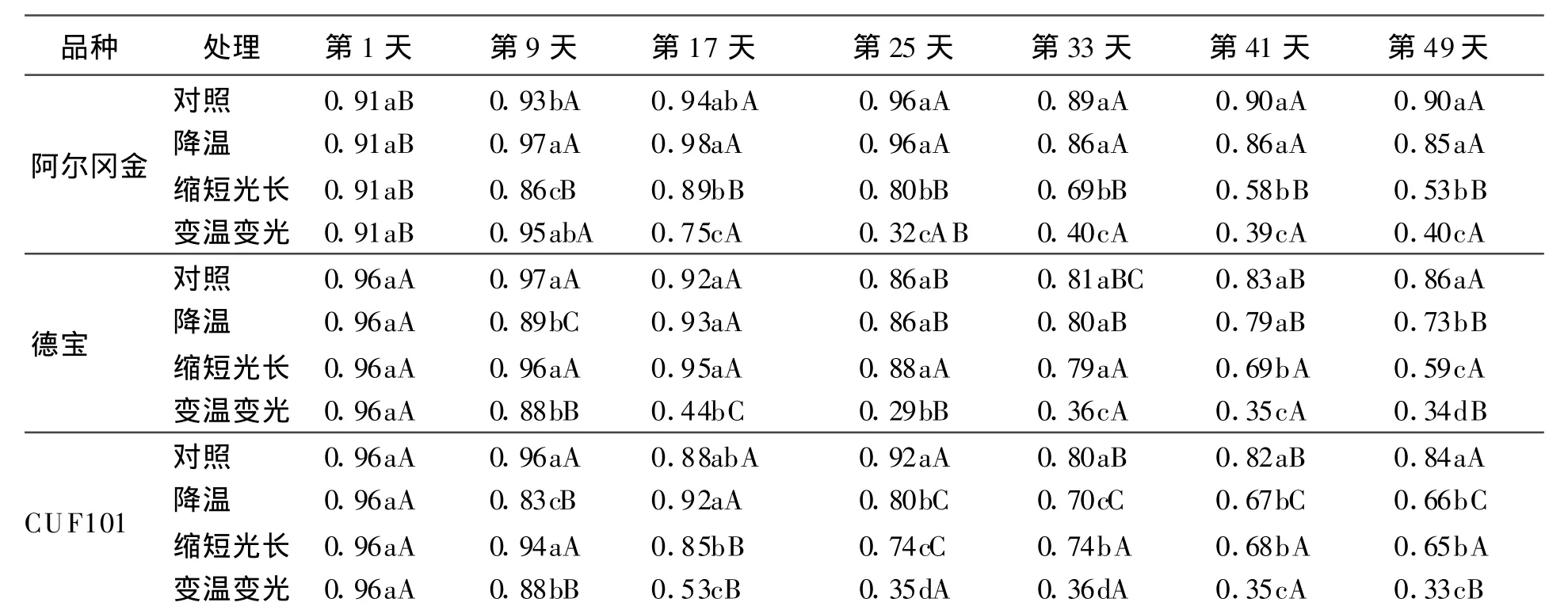
表5 低温和短日照对直立性的影响 cm
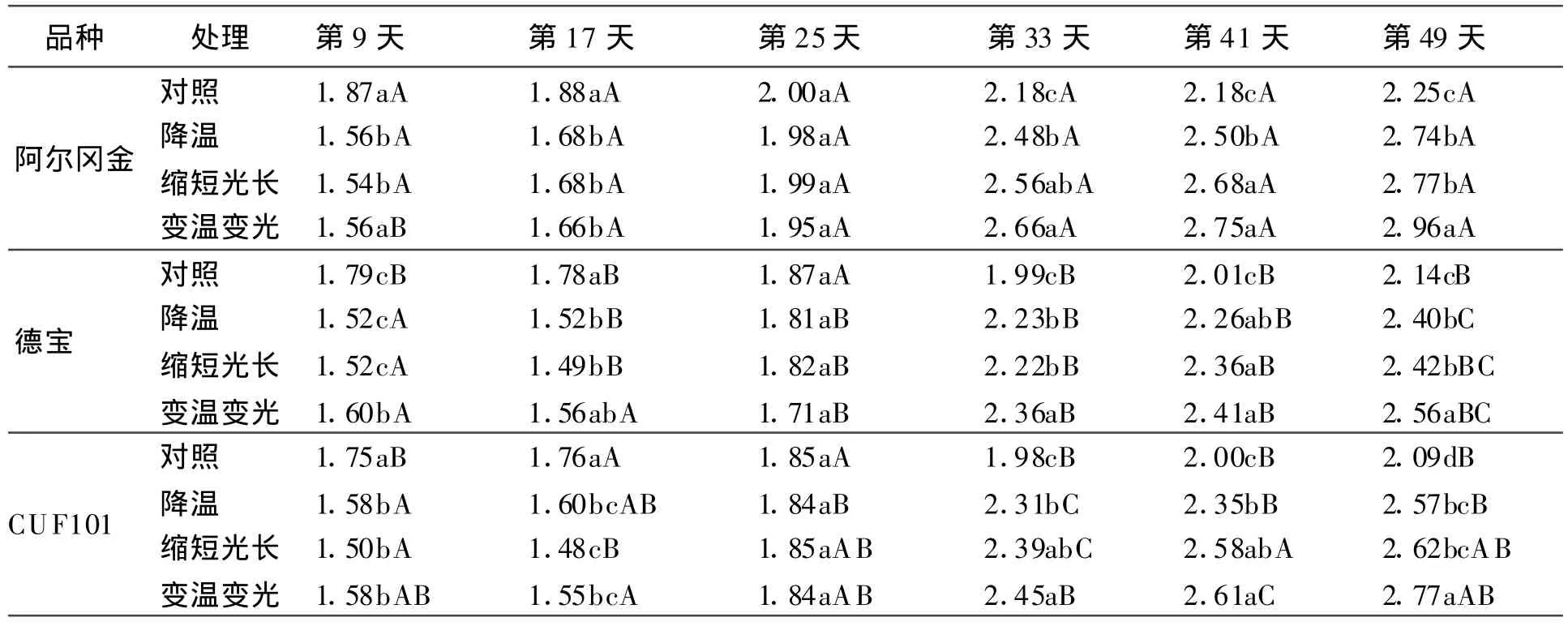
表6 低温和短日照对叶长/叶宽的影响 cm
由图1可以看出,在经过不同处理后,3个品种紫花苜蓿的茎叶比和地上生物量/地下生物量都呈现出一致的变化趋势。经过降温、缩短光长和变温变光处理后,3个品种紫花苜蓿茎叶比都高于对照,地上生物量/地下生物量都小于对照;在同一处理中,阿尔冈金和德宝茎叶比基本相同,都高于CUF101,在各个处理中德宝和阿尔冈金地上生物量/地下生物量的值相差不大,都低于CUF101。

图1 降温和短日照对茎叶比、地上生物量/地下生物量的影响
3 讨论
3.1 不同秋眠型品种分枝数变化分析苜蓿的快速增长往往以出现分枝为标志,当苜蓿到达三叶期后,即可从腋芽上发育分枝,主茎上腋芽不断形成分枝而成为株丛[11]。因此,分枝数可以反映植株的生长发育状况。光长和温度是引起休眠的关键因子,强烈影响腋芽的发育,充足的光照和适宜的温度能够通过促进细胞分裂来促进更多腋芽发育,使分枝数目和长度不断增加。光照和温度对分枝数的影响还与苜蓿的品种有很大关系,邓蓉等[13]通过对紫花苜蓿秋眠性的研究发现秋眠级数与分枝数有显著的相关性。本研究表明,光温条件适宜时,秋眠型品种分枝数要远远多于半秋眠型和非秋眠型品种。非秋眠型品种对温度降低更敏感,降温能更明显抑制非秋眠性品种的分枝发育,使分枝数减少;秋眠型品种分枝数只在降温和光长缩短互作时才显著减少(P<0.05),而光长缩短时各个品种分枝数变化不明显。这是因为温度变化直接影响物质转移运输和对水分的利用效率以及腋芽发育的生理生化反应,甚至促使腋芽进入休眠,从而影响分枝的发育,而光照变化对秋眠型品种的影响并不是马上发生,是经过体内许多基本变化后表现出的综合效应。
3.2 不同秋眠型品种茎粗增长量、平均节间长和直立性变化分析茎的生长反映了苜蓿生长的健壮程度,是综合性状的体现,生长健壮的植株会有较高的生物量。光照条件不仅影响光合作用的速度和能力,还直接影响植物的发育,在长日照植物中表现尤为突出[10]。紫花苜蓿是长日照植物,对光长反应敏感。研究表明,随光照时间从8 h延长到16 h,枝条变的更长更直立,并有较长的茎节和较大直径[14-15]。本研究显示,光温适宜条件下,不同秋眠级紫花苜蓿茎粗,节间长,直立性都维持在一定水平,非秋眠型品种茎秆最粗,秋眠型品种节间长和直立性最大;当降温或光长缩短时,各个品种的茎粗增长量、节间长、直立性都呈下降趋势,这是由于温度降低影响了细胞分裂所需酶的活性,阻碍了细胞分裂和生长,而光长缩短影响了植物有机质的产生和运输,细胞分裂得不到足够的能量,影响植株组织分化和生长发育,导致植株茎杆增长缓慢,植株向上生长减缓或停止,匍匐生长增强。非秋眠型品种节间长、直立性的变化对温度降低反应敏感,秋眠型品种节间长、直立性对光长反应敏感,这可能是由品种间的差异引起的;当降温和光长缩短互作时植株茎秆发育、节间伸长严重受阻,各个品种直立性都迅速降低,植株明显趋向于匍匐性生长。
3.3 不同秋眠型品种叶长/叶宽、茎叶比和地上生物量/地下生物量变化分析苜蓿叶片是苜蓿的同化器官,象征截取光能和制造干物质的能力,叶的生长直接影响到光合面积和光能的利用[11]。光照和温度是影响植物光合作用的主要因素,同时也影响着叶片的形态建成,本研究表明温度降低和光长减少使不同秋眠级品种叶长/叶宽都呈增加趋势,这说明低温和低光照使叶片变薄,变狭长,这样有利于增加光的透射率,尽可能多的弥补损失的光合作用。茎叶比、地上生物量/地下生物量是牧草品质、干物质积累和利用价值等方面的重要指标[16-17]。在光温适宜条件下,植物光合作用、呼吸作用等生理生化活动旺盛,物质转移运输和水分利用效率较高[18]。本研究表明,光温变化使得茎的比例变大,茎叶比增大,这主要是因为降温和光长缩短加速了同化器官(主要是叶)将光合产物向异化器官(主要是茎)的运输;植物地下部分受环境条件改变的影响大于地上部分,因此当降温或光长缩短时地上生物量/地下生物量减小。对于不同秋眠级紫花苜蓿来说,茎叶比和地上生物量/地下生物量对光长缩短的反应比对降温的反应更敏感,这是由于光长缩短直接影响光合作用,影响干物质积累。本研究还表明,当温度降低或光长缩短时,秋眠型品种和半秋眠型品种茎叶比、地上生物量/地下生物量的变化大于非秋眠型品种的变化,这可能是因为光长变短和(或)温度降低时,秋眠型品种更容易被触发,地上部分生长减缓,为地下部分储存营养。
4 结论
光温适宜条件下,秋眠型紫花苜蓿分枝数最多,半秋眠型品种次之,非秋眠型品种最少。非秋眠型紫花苜蓿分枝数变化对降温的反应比对光长缩短反应敏感,秋眠型品种只有在降温和光长缩短互作时,分枝数显著减少(P<0.05)。
降温和光长缩短时,茎秆变细,茎节缩短,苜蓿植株直立性降低。温度和光长交互作用对茎秆生长作用最为明显。秋眠型品种这些形态的变化受光长变化影响较大,非秋眠型品种则受温度变化影响较大。
降温或光长缩短使不同秋眠型品种叶长/叶宽增大,茎叶比增加,地上生物量/地下生物量降低,其中降温和光长缩短交互作用影响最明显,光长缩短对于3个品种的影响大于降温的影响。
[1]马其东,叶建敏.紫花苜蓿栽培管理技术[J].中国奶牛,2001(2):23-24.
[2]岳常彦,郭金龙,王洪水.种植牧草改良盐碱地和油污地[J].山东畜牧兽医,2001(6):49-50.
[3]张少华,张敦新,张银会,等.论陇东紫花苜蓿的产业化[J].草业科学,2000,17(2):19-22.
[4]阎旭东,翟玉柱,朱志明,等.八个苜蓿品种的耐盐性分析[A].见:刘小京,刘孟雨.盐生植物利用与区域农业可持续发展[C].北京:气象出版社,2002:275-277.
[5]马晖玲,卢欣石,曹致中,等.紫花苜蓿不同栽培品种植株再生的研究[J].草业学报,2004,13(6):99-105.
[6]李向林,万里强.苜蓿秋眠性及其与抗寒性和产量的关系[J].草业学报,2004,13(3):57-61.
[7]毕玉芬.新疆北部苜蓿属植物秋眠性[J].安徽农业大学学报,2002,29(4):382-386.
[8]卢欣石.中国苜蓿审定品种秋眠性研究[J].中国草地,1998(3):1-5,12.
[9]于林清,王照兰.中国新疆苜蓿野生种群秋眠性的研究[J].中国草地,2001(3):13-16.
[10]张世君.紫花苜蓿秋眠级及其应用[J].草业科学,2002,19(4):79.
[11]洪绂曾.苜蓿科学[M].北京:中国农业出版社,2009.
[12]卢欣石,申玉龙.苜蓿秋眠性研究与利用[J].草原与牧草,1991(4):1-4.
[13]邓蓉,向清华,陈武,等.紫花苜蓿秋眠性的研究[J].草业科学,2005,22(5),41-44.
[14]Sato K.Growth and development of Lucerne plants in a controlled environment.The effects of daylength and tempreature on growth and chemical composition[J].Proceedings of the Crop Science Society of Japan,1971,40:120-126.
[15]Sato K.Growth and development of Lucerne plants in a controlled environment.1.The effects of day length and temperature before and after cutting on the regrowth process[J].Journal of The Japanese Society For Grass Science and Technology,1971,17:127-132.
[16]郞炳耀.苜蓿生长发育及产量形成与温度和水分的关系[J].内蒙古草业,1990(2):76-80.
[17]郭孝.苇状羊茅生产性能的综合研究[J].草业科学,1998,15(2):24-26.
[18]贾慎修.草地学[M].第二版.北京:中国农业出版社,2001.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
语文周报·教研版(2021年28期)2021-08-19
现代畜牧科技(2021年4期)2021-07-21
小哥白尼(神奇星球)(2020年7期)2021-01-18
陕西农业科学(2019年4期)2019-05-13
老友(2017年7期)2017-08-22
中国三峡(2017年4期)2017-06-06
能源(2016年1期)2016-12-01
为了孩子(3~7岁)(2016年6期)2016-05-14
新疆农业科技(2016年6期)2016-02-19
