
eha,迟缓爱德华菌一个毒力调控基因*
2010-01-24 07:43:30郑恩金高大庆陆承平
中国人兽共患病学报 2010年11期
郑恩金,高大庆,洪 捷,陆承平
eha,迟缓爱德华菌一个毒力调控基因*
郑恩金1,高大庆1,洪 捷1,陆承平2
目的 探讨迟缓爱德华菌(Edw ardsiella tarda,E.tarda)溶血相关基因(E.tarda haemolysin activator gene,eha)基因调控该菌毒力基因的作用。方法利用自杀质粒p HM 5,缺失 E.tarda的eha基因,得到△eha缺失菌株,再构建△eha株的互补菌株。通过平板溶血性法、接触溶血法、上清溶血法,观察野生株、△eha缺失株与及互补株溶血性的差异。利用M TT法比较三种细菌培养物的过滤液对Vero细胞的毒性的差别。利用H2O2抗性纸片扩散法,比较三种菌对过氧化氢抵抗力的差异。利用RT-PCR和SDD-PAGE电泳超速酸化离心法提取的鞭毛蛋白,比较三种细菌鞭毛基因的转录和表达的差异。结果△eha缺失株和互补株不溶血,而野生株溶血。eha基因的缺失降低E.tarda菌对过氧化氢的抵抗力,△eha缺失株较野生株的细胞毒性明显减弱,eha基因可以调控迟缓爱德菌鞭毛基因的转录和表达。结论eha基因可以调控迟缓爱德华菌毒力基因的表达,是一个毒力调控基因。
迟缓爱德华菌;eha基因;基因缺失;基因调控
E.tarda属于肠杆菌科爱德华菌属是一种既感染动物又感染人的致病菌,特别会导致许多水生动物致病,给水产养殖业带来了很大的危害〔1〕。国内外对 E.tarda的感染报道较多,但对其致病机制研究甚少,高大庆等通过鸟枪法,从 E.tarda中发现了一个新的溶血相关基因eha〔2-3〕,有关研究表明,eha不是溶血素结构基因,而是激活大肠埃希菌中clyA(cytolysin A)基因的表达〔4〕。eha基因和伤寒沙门氏菌转录调控基因slyA有68%同源性。因此,eha基因可能对 E.tarda其它基因具有调控作用。本研究利用自杀质粒同源重组的方法,构建△eha缺失株。通过比较 E.tarda野生株、△eha缺失株和互补株毒力方面的差异,分析eha对 E.tarda毒力的影响,有助于进一步理解 E.tarda的致病机制。
1 材料和方法
1.1 质粒和菌株 受体菌株,E.coil DH5α由本实验室保存;E.coil SM 10(λpir)由中国疾病预防控制
中心的梁未丽教授惠赠;E tarda ET13株由南京农业大学的陆承平教授惠赠;质粒pACYC 184和自杀质粒p HM 5均由本实验室保存。
1.2 培养基 细菌培养选用LB(Luria Broth)培养基 抗生素使用浓度分别为:氨苄青霉素(Amp)100μg/m L,氯霉素 (Cm)50μg/m L,粘菌素 (Col)50μg/m L。
1.3 主要试剂 RNA提取试剂(Om iga公司);DNA提取试剂盒、胶回收试剂盒(上海化舜公司);RT-PCR试剂 (TaKaRa公司);引物由上海英骏生物有限公司合成。
1.4PCR扩增 以提取 ET-13株基因组为模板,以F1-F:GAAGA TCTTA TTGCTGAGGGAACCGTCA和F1-R:GCTCTAGA CGTCGACCACTCGTACCAACCGTGA为引物序列,扩增 eha基因上游F1片段。以F2-F:GCTCTAGA TTTGGTAGA GCGA TTGGAAC和F2-R:CGAGCTCGCTTCT TAACCCACC GA TTT为引物序列,扩增eha基因下游F2片段。PCR反应条件为:94℃3min,94℃30s,58℃(F1)52℃(F2)45s,72℃60s,扩增反应为35个循环,最后72℃延伸10min。PCR产物经琼脂糖凝胶电泳鉴定。
1.5 重组自杀质粒的制备 双酶切F1和F2片段及自杀质粒载体p HM 5,用胶回收试剂盒回收,16℃连接过夜,用氯化钙法转化大肠杆菌SM 10(λpir)感受态细胞。在含AmprLB平板上挑取阳性克隆。提取重组自杀质粒,酶切鉴定,命名为p HM-F。
1.6 缺失菌株构建和鉴定 用接合的方法将重组自杀质粒p HM-F从 SM 10λpi r转移到 E.tarda ET13。在Ampr和ColrLB平板上,筛选含p HM-F的 ET13菌落。再通过二次交换,在Col和10%蔗糖的LB平板上,筛选到缺失株。再将缺失株反复稳定传代。另设计鉴定引物-F:GGCGTCTTACGCCGTTTAGC和鉴定引物-R:GTACCGA TGGGGTGCAGTTG,缺失株经 PCR扩增鉴定,命名为△eha缺失株。
1.7 互补菌株的构建 以ET13株基因组DNA为模板,PCR扩增 eha基因的片段,引物序列为eha-F:CCAAGCTTTTGGAA TCGACA TTGGGCT和eha-R:CGGGA TCCTTA TTTA TTCTGTAAGTGA,eha基因和载体pACYC184连接,得到重组质粒p ACEHA。通过电击法将p ACEHA转入到△eha缺失株,得到互补株,命名为ehaComp。
1.8 H2O2抗性纸片扩散法 一定浓度的细菌均匀涂到LB平板,再将不同浓度的 H2O2纸片置于平板上,16小时观察结果,测量抑菌圈直径(mm)。
1.9 溶血性分析 平板检测法:参照文献〔5〕,接触溶血法将100μL过夜培养菌液,在“V”形96孔微量滴定板上与等体积1%兔红细胞混合,37℃作用1h,用酶联免疫检测仪在OD540nm和OD600nm处测量各孔的吸光值,根据计算公式:溶血活性单位(hemolytic activity units,Hem.act.)= A 540/A 600×100(U)。上清检测法将过夜培养菌液离心,上清过滤除菌,将100μL上清和等体积1%兔红细胞混合,同上法测定上清的溶血性。
1.10 Vero细胞毒性试验 取培养至OD600约0.8的菌液5 m L,离心 5 000g×30 min。上清经过滤菌器过滤。同时收集对数期Vero细胞,至细胞单层铺满96孔底,分别加入10μL,50μL和100μL的滤液,再加 M TT溶液,用在酶联免疫检测仪在OD490nm处测量各孔的吸光值,计算Vero细胞的存活率。
1.11 酸化超速离心法法 取野生株和突变株的单个菌落接种至LB中,摇床(37±2)℃孵育过夜后,移种至500m L LB培养基中,放入恒温摇箱 (37±2)℃孵育18~24 h,测定OD值,使其OD值基本一致。提取细菌鞭毛蛋白方法参考文献〔6〕,分别溶于5 mL PBS,保存-20℃备用。
1.12 SDS– PAGE蛋白电泳,检测细菌鞭毛蛋白 ,方法参考文献〔6〕。加样量 15μL/每孔 ,在 1 ×Tris一甘氨酸凝胶电泳缓冲液中电泳。样品在浓缩胶中的电泳电压为80伏,在分离胶的电泳电压为120伏。电泳后的凝胶置于考马斯亮兰 R-250染色液中染色过夜,脱色液脱色后观察结果。
1.13 RT-PCR法 提取细菌 ET-13株总RNA,采用随机引物,按照Promega公司反转录试剂盒说明书操作,逆转录合成 cDNA。用引物 Flic-F:TAGAAGCAGGAAAA TGGGG和Flic-R:ACGC GGTAA TGCGCGA TAACG扩增鞭毛基因,同时扩增16SrRNA基因作为内标参照,引物序列为16srRNA引物-F:TGAA GAAGGCCTTCGGGTTG和 16srRNA引物-R:TTACTAGCGA TTCCGACTTC,半定量检测鞭毛基因的转录。
2 结 果
2.1 △eha缺失菌株的构建及鉴定 本实验构建的自杀性重组质粒p HM-F含有eha基因两侧 F1和F2片段,将其通过接合转入 E.tarda ET13株,其两侧片段与 ET-13株染色体上eha基因两侧片段可发生同源重组,质粒上的Amp基因被随之带入 ET-13株中,在Ampr和 ColrLB平板上,筛选含p HM-F的 ET13菌落。在Col和10%蔗糖的LB平板上,筛选到自杀性质粒脱离的△eha缺失株。
用 PCR扩增进一步鉴定突变株,即扩增片段478bp为缺失株,扩增片段825bp为野生株,PCR鉴定 。经过十几代传代后,可得到稳定的△eha缺失株。
2.2 互补株的构建 根据eha基因序列设计Eha-F和 Eha-R为引物,以 ET-13的DNA为模板,扩增eha基因 432bp。用 Bam H I和 H in d III双酶切产物,与载体pACYC184相同酶切产物连接,转化E.coil DH5α,在氯霉素LB平板上筛选阳性转化子,抽提重组质粒进行酶切和PCR鉴定,重组质粒命名为pACEHA。将该质粒电击入△eha缺失株,在氯霉素LB平板上获得互补菌株,命名为ehaComp互补株。
2.3 溶血试验 从图1可以看出,野生株较△eha突变株和互补株在血平板有明显溶血现象。

图1 细菌血平板溶血现象Fig.1 The bacterial hemolytic crisis on blood plate 1.the△eha strains;2.the ET-13 wild strains;3.the comp lement strains
从表1可以看出,用接触溶血(Contact Hemolysis,CH)法检测,野生株的溶血活性明显高于缺失株和互补株,有统计学意义(P<0.05),缺失株和互补株的溶血活性无统计学意义(P>0.05)。用上清检测(Supernatant A ssay,SA)法检测,三种菌株均无溶血活性。
2.4 Vero细胞毒性实验 细胞的存活率如表2所示。经进行统计学分析,加入野生株与缺失株、互补株各 50μL滤液量时,细胞存活率有差异(P<0.05),加入50μL滤液,野生株与缺失株和互补株的细胞存活率差异明显,说明缺失株和互补株较野生株分泌到培养基的细胞毒素少;而加入各10μL和100μL滤液量时,细胞存活率差异均不显著。
2.5 RT-PCR 结果见图2:分别以 ET-13野生株、△eha缺失株及ehaComp互补株的RNA逆转录合成的cDNA为模板,同时扩增16SrRNA的基因和鞭毛基因 f lic。根据电泳结果,以扩增的16SrRNA基因为内标参照,野生株和缺失株的扩增16SrRNA基因产物亮度基本一致前提下,野生株扩增f lic基因产物明显亮于缺失株。互补株扩增16SrRNA基因的产物亮度强于野生株,但是互补株扩增出的 f lic基因产物却明显弱于野生株。

表1 细菌接触溶血和细菌培养上清溶血性的结果Tab.1 The result of bacterial contact hemolysis and culture supernatant hemolysis
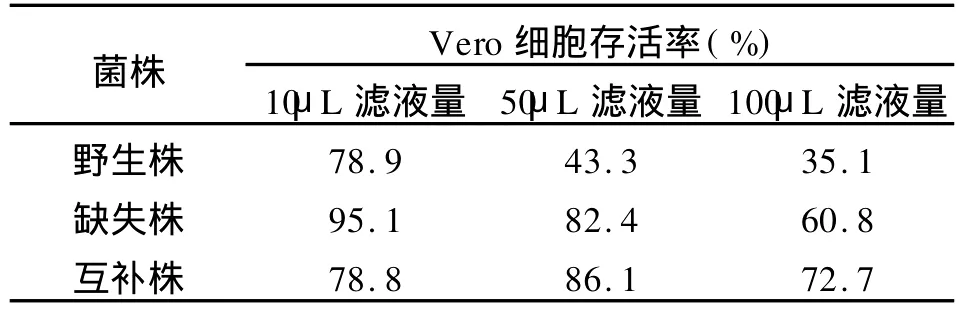
表2 细菌培养过滤液的Vero细胞毒性(M TT法)Tab.2 The Vero cell’cytotoxicity for the filtering culture of the bacteria by MTT

图2 RT-PCR扩增鞭毛基因和16Sr RNA基因Fig.2 Amplification of the flagellar gene and 16SrRNA gene by RT-PCR 1.DNA marker;2,6.the ET-13 wild strains;3,7.the△eha strains;4,8.the comp lement strains;2-4.amp lification of the flagellar gene;6-8.amp lification of 16SrRNA gene
2.6 通过蛋白SDS-PAGE电泳检测 结果见图3条带主要在43kD与56kD之间,与 E.tarda鞭毛蛋白分子量44kD一致。在 ET-13野生株和△eha缺失株细菌浓度一致的情况下,从野生株中提取的鞭毛蛋白FliC的量高于缺失株。

图3 鞭毛蛋白的SDS-PAGE电泳1.蛋白marker;2.△eha缺失株的鞭毛蛋白;3.ET-13野生株的鞭毛蛋白Fig.3 The SDS-PAGE electrophoresis of the flagellin protein 1.protein marker;2.the flagellin proteins from△eha strains;3.the flagellin proteins from the ET-13 strains
2.7 H2O2纸片扩散实验 通过测量抑菌环直径,并对结果(图4)进行统计学分析显示,4个不同 H2O2浓度作用下,突变株的抑菌环直径均大于野生株和互补株。抑菌环直径与纸片 H2O2浓度呈现出正比趋势。
3 讨 论
病原细菌侵袭宿主细胞,产生毒力因子使宿主致病的过程,是一个紧密调控的过程。即病原细菌从感应信号刺激,到接受刺激的调控子将激活或抑制相应毒力因子靶基因的转录和表达,这样组成了一个毒力调控系统。为了能适应各种环境,E.tarda菌已经进化出了一些精细的毒力调控系统对环境作出适宜反应。2009年中国科学家〔7〕公布了该菌EIB202菌株全基因组序列,测序结果表明:E.tarda菌染色体上存在和其他病原菌相似的毒力调控基因,如二元调控系统(Two-component Regulatory System s),密度感应(Quorum Sensing),铁摄取调控子(ferric up take regulato r,fur),Sly(Salmonella hemolysin,slyA)等,但这些毒力调控基因在 E.tarda菌中如何发挥作用,有待研究。在 Et-EIB202菌全基因组序列中,标签为 ETAE-1684基因则直接称为slyA,该基因序列和eha基因序列有100%同源性,即为eha基因。它和伤寒沙门氏菌转录调控基因 slyA有68%同源性。slyA初次发现时,被认为它是一种新的溶血素基因,以后的研究表明,slyA不仅能调控伤寒沙门氏菌毒力二岛 (Salmonella Pathogenicity island,SPI)等表面结构基因,而且调控伤寒沙门氏菌中产生细胞毒性的胶原酶表达、抗菌肽耐药性的产生,因此,slyA作为伤寒沙门氏菌重要转录调控子〔8〕。虽然 eha和slyA都属于 M arR〔9〕家族中的一员,它对 E.tarda其它基因是否具有调控作用,有待研究。
有研究〔10〕表明 E.tarda侵袭并进入宿主细胞,可能是由溶血素引起的。目前认为,至少存在两种溶血素,一种是细胞结合性的溶血素,另一种是使细胞穿孔的分泌型溶血素。本研究显示eha基因可以影响E.tarda毒素的产生,eha缺失后会影响细菌接触性溶血和血平板的溶血现象,但是eha基因是否影响E.tarda其它毒素的产生及对何种溶血素的表达有作用,还有待研究。另外,鞭毛介导的运动能力被认为在 E.tarda致病早期起着重要作用,它使细菌稳定地附着并在粘膜内大量繁殖。flic基因控制细菌鞭毛素蛋白(flagellin)的产生,本研究表明eha基因可以影响f lic基因的转录和表达。
宿主在抵抗病原菌侵袭的过程中,其巨噬细胞可以产生对微生物杀伤的反应性氧中间物,如H2O2等。本研究表明△eha缺失株相对于野生株对过氧化氢的抵抗力是降低的,所以eha基因可能影响 E.tarda在细胞内的存活。
因此,eha基因调控了 E.tarda菌一些毒力基因的表达,是细菌的一种毒力转录调控因子。本研究通过探讨eha基因调控E.tarda菌靶基因表达的机制,有助于加深对 E.tarda菌的宿主适应力和致病机制的认识。
〔1〕Xiao J,Wang Q,Liu Q,et al.Isolation and identification of fish pathogen Edwardsiella tarda from mariculture in China〔J〕.Aquac Res,2008,40(1):13-17.
〔2〕高大庆,陆承平,吴守一,等.迟缓爱德华氏菌的溶血相关基因的克隆〔J〕.中国人兽共患病杂志,2000,16(5):51-52.
〔3〕高大庆,阚飙,陆承平,等.迟缓爱德华菌溶血相关基因的测序和初步的功能分析〔J〕.遗传学报,2001,25(12):1162-1167.
〔4〕成静,韩晶晶,印文静,等.迟缓爱德华菌 eha基因对大肠埃希氏菌溶血基因cly A转录的调控〔J〕.中国人兽共患病学报,2007,23(12)15-18.
〔5〕高大庆,黄锡全,陆承平,等.迟缓爱德华氏菌的溶血特性的检测〔J〕.中国人兽共患病杂志,2000,16(4):66-68.
〔6〕Morohoshi T,Yokoyama Y,OuchiM,et al.Motility and the expression of the flagellin protein FliC are negatively regulated by quorum sensing in Edw ardsiella tarda〔J〕.JBioscience Bioengineer,2009,108(4),314-318.
〔7〕Wang Q,Yang M.,Xiao J,et al.Genome sequenceof the versatile fish pathogen Edw ardsiella tarda provides insights into its adaptation to broad host ranges and intracellular niches〔J〕.PLos One,2009,4(10):e7646.
〔8〕Buchmeier N,Bossie S,Chen CY,et al.SlyA,a transcriptional regulator of Salmonella typhimurium,is required for resistance to oxidative stress and is expressed in the intracellular environment of macrophages〔J〕. Infect Immun,1997,65(9):3725-3730.
〔9〕Haque MM,Kabir MS,Kabir MS,et al.SlyA,a MarR family transcriptional regulator,isessential for virulence in Dickeya dadantii 3937〔J〕.JBacteriol,2009,191(17):5409-5418.
〔10〕Wang F,Zhang M,Hu YH,et al.Regulation of the Edw ardsiella tarda hemolysin gene and luxS by EthR〔J〕.J Microbiol Biotechnol,2009,19(8):765-773.
eha,a regulating virulence gene of Edw ardsiella tarda
ZHENG En-jin1,GAO Da-qing1,HONG Jie1,LU Cheng-ping2
(1.Southeast University;2 N anjing A gricultural University,N anning 210009,China)
To exp lo re the function of eha gene on regulating E.tarda virulence geneswhich was deleted by suicide plasmid p HM 5 from E.tarda.The△eha mutant strain wasobtained and the△eha comp lementary strain was constructed.Differences among the wild strains,mutant strains and comp lementary strains were compared by p late hemolysis,contact hemolysis and supernatant assay,respectively.The filtering cultures of the three kindsof E.tarda were used to test verocellcyto toxicity with M TTmethod.Comparisonsof the three kindsof E.tarda against hydrogen peroxidewere conducted by scrip diffusion and the bacterial flagellin protein was purified by acidification excessive speed centrifugation.The transcription and expression on the bacterial flagellin gene of them were also compared by RT-PCR and SDS-PAGE electropho resis.Results indicated that the mutant and comp lementary strains were non-hemolytic,while the wild strains were hemolytic.E.tarda decreased its resistance against hydrogen peroxide due to deletion eha and mutant strains produced less cytotoxicity than the wild ones.The transcription and expression of flagellin gene were regulated by eha gene.It’s concluded that eha gene p lays a regulato ry role on the expression of virulence gene of E.tarda and the eha gene was a regulating virulence gene of E.tarda.
E.tarda;eha gene;gene deletion;regulation
R378.2
A
1002-2694(2010)11-0999-05
*东南大学人才引进科研启动基金(No.4023001015)资助
高大庆,Email:dgao2@yahoo.com
1.东南大学医学院,南京 210009;
2.南京农业大学动物医学院,南京 210095;
2010-06-17;
2010-08-31
猜你喜欢
云南化工(2021年6期)2021-12-21 07:31:04
数学大王·中高年级(2021年4期)2021-04-27 11:21:14
农药科学与管理(2019年6期)2019-11-23 08:17:12
家庭影院技术(2019年8期)2019-08-27 02:44:52
食品科学(2018年10期)2018-05-23 01:27:28
华南农业大学学报(2015年5期)2015-12-04 03:04:38
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
计算机与网络(2014年9期)2014-03-25 10:57:13
