
基于MaxEnt模型预测瓜类细菌性果斑病在中国的潜在地理分布
2025-02-19 00:00:00张文瑶陈雪荣谢洪芳田艳丽胡白石
植物保护 2025年1期

摘要
瓜类细菌性果斑病(bacterial fruit blotch)是瓜类作物上重要的种传细菌性病害, 病原菌为西瓜噬酸菌Acidovorax citrulli。我国是全球西甜瓜的主要生产区。近年来, 瓜类细菌性果斑病的频繁发生已严重影响我国西甜瓜产业的健康发展。为明确瓜类细菌性果斑病在我国的适生性, 根据其在全球的最新分布数据, 本研究利用MaxEnt模型结合ArcGIS软件预测了瓜类细菌性果斑病在我国的潜在地理分布。结果表明, MaxEnt模型的平均AUC (area under curve, AUC)值均大于0.9, 预测结果的准确性较高。在历史气候条件下,瓜类细菌性果斑病适生区分布广泛, 主要包括华中、华南和华东地区, 以及部分华北、东北地区, 占我国面积的47.36%。影响瓜类细菌性果斑病在我国潜在分布区域的主要气候因子包括最热月份最高温度、月平均昼夜温差、最干月份降水量和最干季平均温度。未来气候情景无论是低环境强迫还是高环境强迫, 适生区面积均呈现增长的趋势, 预示着随着气候的变化, 瓜类细菌性果斑病在我国发生的风险不断增加, 因此建议应加强检疫监测和防控, 严防其扩散。
关键词
瓜类细菌性果斑病;" MaxEnt模型;" 气候变化;" 潜在地理分布
中图分类号:
S 432.1
文献标识码:" A
DOI:" 10.16688/j.zwbh.2024066
收稿日期:" 20240202""" 修订日期:" 20240312
基金项目:
江苏现代农业产业技术体系建设项目(JATS[2023]315)
致" 谢:" 参加本试验部分工作的还有江代礼、谭翰杰、张能和纪烨斌等同学,特此一并致谢。
* 通信作者
E-mail:
tianyanli@njau.edu.cn
#
为并列第一作者
Potential distribution prediction of bacterial fruit blotch of melons in China based on MaxEnt
ZHANG Wenyao1," CHEN Xuerong2," XIE Hongfang3," TIAN Yanli1,4*," HU Baishi1,4
(1. College of Plant Protection, Nanjing Agricultural University, Key Laboratory of Surveillance and Management
for Plant Quarantine Pests, Ministry of Agriculture and Rural Affairs, Nanjing" 210095, China; 2. Agricultural
Technology Extension Service Center of Pengyang County, Guyuan City, Ningxia Hui Autonomous Region,
Guyuan" 756599, China; 3. Plant Protection and Quarantine Station of Nanjing, Nanjing" 210008, China;
4. Xinjiang Key Laboratory of Agricultural Biosafety, Urumqi" 830091, China)
Abstract
Acidovorax citrulli, is the causal agent of bacterial fruit blotch (BFB), a serious seed borne disease affecting cucurbit crops worldwide, particularly watermelon and melon. China is a major global producer of these crops. In recent years, frequent BFB outbreaks have significantly impacted the healthy development of China’s watermelon industry." This study predicted the potential distribution of BFB in China, using the MaxEnt model combined with ArcGIS, based on recent global distribution data. The average AUC values of all models exceeded 0.9, indicating high prediction accuracy. Under historical climate conditions, suitable areas for BFP cover 47.36% of China’s area, including most of central, south and east China, as well as parts of north and northeast China. The main climate factors influencing the potential distribution were the maximum temperature of the warmest month, mean diurnal range monthly, precipitation of the driest month, and mean temperature of the driest quarter. In future climate scenarios, both low and high environmental pressure scenarios, the suitable areas will expand, indicating an increased risk of BFB occurrence with climate change. Therefore, it is recommended to strengthen quarantine monitoring and prevention efforts to prevent its spread.
Key words
bacterial fruit blotch of melons;" MaxEnt model;" climate change;" potential distribution
瓜类细菌性果斑病菌(西瓜噬酸菌)Acidovorax citrulli[1]是我国进境植物检疫性有害生物及全国农业植物检疫性有害生物,主要危害西瓜、甜瓜等葫芦科作物。该病自1965年首次在美国报道以来,已在世界上许多西甜瓜产区相继发生,造成巨大的经济损失[2]。我国在20世纪90年代首次报道瓜类细菌性果斑病发生,随后在我国内蒙古、广东、福建、山东等地均有发生,造成大田西瓜和甜瓜减产甚至绝收[37]。带菌种子是瓜类细菌性果斑病最主要的初侵染源和远距离传播的病菌载体,也是近10年来该病在我国传播的主要原因。然而,调查研究发现,国内西甜瓜以及砧木种子携带瓜类细菌性果斑病菌的带菌率较高[8]。因此,瓜类细菌性果斑病的防控技术应以种子处理为主,尤其是在病害发生的适生区加强种子检疫和消毒,降低病害发生风险。综上所述,明确瓜类细菌性果斑病菌在中国的潜在地理分布,对于该病害的精准防控具有重要意义。
物种分布模型(species distribution model, SDM)也称生态位模型(ecological niche models, ENM), 是一种基于物种分布点或丰富度及分布地区环境变量(气候、海拔、植被等)的数学模型[9]。模型依据分布信息估测物种所需的生态环境, 将结果投射到不同的时空中, 以概率的形式预测物种当前或未来的分布情况[10], 模型结果通常反映大尺度空间上物种适宜生境的分布。作为研究环境变量对物种分布影响的重要方法, SDM被广泛应用于入侵物种潜在分布区预测、气候变化对物种丰富度的影响、物种生境的适宜度评价等[11]。MaxEnt (maximum entropy, MaxEnt)模型作为当下应用较多的一种SDM模型, 主要利用物种分布点和环境变量及基于机器学习和最大熵原理来模拟物种潜在分布[1213], 即使在物种分布信息较少的情况下也能有较好的预测结果[14], 被广泛应用于多种有害生物的适生性分析, 包括柑橘木虱Diaphorina citri[15]、腐烂茎线虫 Ditylenchus destructor[16]、玉米褪绿斑驳病毒Maize chlorotic mottle virus[17]、落叶松枯梢病菌Neofusicoccum laricinum[18]和梨火疫病菌Erwinia amylovora[19]。目前, 关于瓜类细菌性果斑病菌的研究报道主要集中于快速检测方法建立[2022]、致病机制解析[2326]、防控药剂筛选[2729]等方面, 尚未有关于其在我国潜在地理分布的研究。
本研究利用MaxEnt模型, 对瓜类细菌性果斑病在我国的适生区进行了预测, 利用刀切法探究了影响因素较高的气候因子, 对这些气候因子的响应曲线进行了分析, 定量描述了
瓜类细菌性果斑病适生的环境条件, 分析不同环境情景下的气候数据,预测未来适生区域的转移情况, 为病害的预测预报和有效防控提供了一定的理论基础。
1" 材料与方法
1.1" 材料
1.1.1" 气候数据
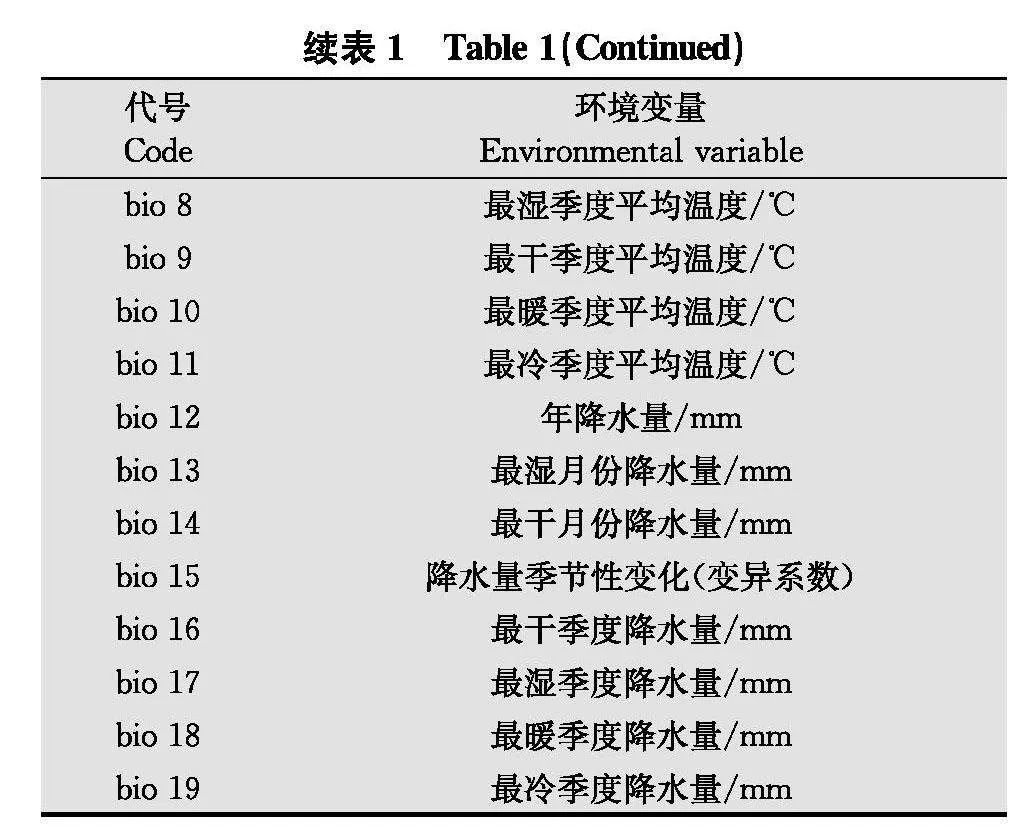
本研究中过去气候采用1970年-2000年的19个气候因子bio 1~bio 19(表1), 数据来源于WorldClim 2.1数据库(https:∥worldclim.org/), 分辨率为2.5 arc-minutes。未来气候情景选择第6次国际耦合模式比较计划(coupled model intercomparison project phase 6, CMIP6), 国家(北京)气候中心气候系统模式的中等分辨率(Beijing climate center climate system model 2 medium resolution, BCC-CSM2-MR)[30]。BCC-CSM2-MR主要包含4种情景, 即低强迫情景SSP126、中等强迫情景SSP245、中高等强迫情景SSP370和高强迫情景SSP585, 并且相较于CMIP5中的全球模式,在气温和降水等方面的模拟能力有明显的提高[30]。本研究针对2021年-2040年、2041年-2060年、2061年-2080年、2081年-2100年这4个时间段,选取低强迫SSP126和高强迫SSP585两种情景,预测未来瓜类细菌性果斑病适生区的变化情况。
1.1.2" 地图数据
从标准地图服务系统(http:∥bzdt.cn.mn.gov.cn/)下载中华人民共和国国界和省界以及省级行政区划图的矢量文件。
1.1.3" 分布点数据
查阅公开发表的文献和历年国家公布的《全国农业植物检疫性有害生物分布行政区名录》, 共获得143个物种分布点信息数据。
1.1.4" 软件版本
MaxEnt 3.4.4 来源于(https:∥biodiversityinformatics.amnh.org/open_source/maxent/)。ArcGIS使用10.8版本, 插件SDM Toolbox采用2.6版本, ENMTools采用1.0版本。
1.2" 数据处理方法
1.2.1" 气候数据处理
过去气候数据使用ArcGIS中SDM Toolbox插件, 以中国矢量地图为掩膜裁剪, 并转为asc格式。未来气候数据首先需要在ArcGIS中按照波段分别提取19个气候因子的信息, 再裁剪出中国的数据保存为asc格式。
1.2.2" 地图数据处理
所有使用的地图相关数据都在ArcGIS中统一坐标系, 地理坐标系选择WGS 1984, 投影坐标系选择Albers。
1.2.3" 分布点数据处理
将所有分布点在Excel中输入十进制格式的经纬度信息, 并在ArcGIS中使用SDM Toolbox除去自相关的点位, 保证每一个栅格(raster)中只保留离中心最近的一个点, 防止过拟合的分布点信息对模型数据产生影响[32]。将最终得到的135个点位分别按物种名(species)、经度(longitude)、纬度(latitude)顺序记录, 并保存为csv格式。
1.3" MaxEnt模型构建和检验
1.3.1" MaxEnt模型构建
首先使用ENMTools计算每两个气候变量之间的皮尔逊系数,进行多重共线性检验[33]。当两个气候之间的相关系数绝对值≥0.8时,说明两者高度相关,需要比较两者的贡献度,删去贡献度低的气候因子[14]。在MaxEnt软件中分别输入删去过拟合数据的分布点经纬度和筛选过后的气候因子,采用自举法(bootstrap),选取25%点位作为测试集,剩余75%分布点作为训练集,勾选刀切法(jackknife)和响应曲线(response curves),重复次数为10次。刀切法被用于测试各个环境变量对模型构建的重要程度以及影响力[14]。输出格式(output format)中,cloglog输出是当前预测适生区最优的输出方式[34],所以本研究采用该输出形式。多次重复运行后,输出我国瓜类细菌性果斑病适生区分布的预测结果,输出格式为ASCII,利用ArcGIS软件转化成栅格形式显示。
1.3.2" MaxEnt模型检验
AUC值(area under curve, AUC)为受试者工作特征曲线(receiver operating characteristic curve, ROC曲线)下面积值。AUC的取值范围为[0,1], AUC值越接近1表明预测结果精度越高[35]。当0.6lt;AUC值≤0.7,模型预测结果较差;当0.70lt;AUC值≤0.80,模型预测结果一般;当0.80lt;AUC值≤0.90,模型预测结果好;当0.90lt;AUC值≤1.00,模型预测结果非常好[36]。
2" 结果与分析
2.1" 模型精度检验
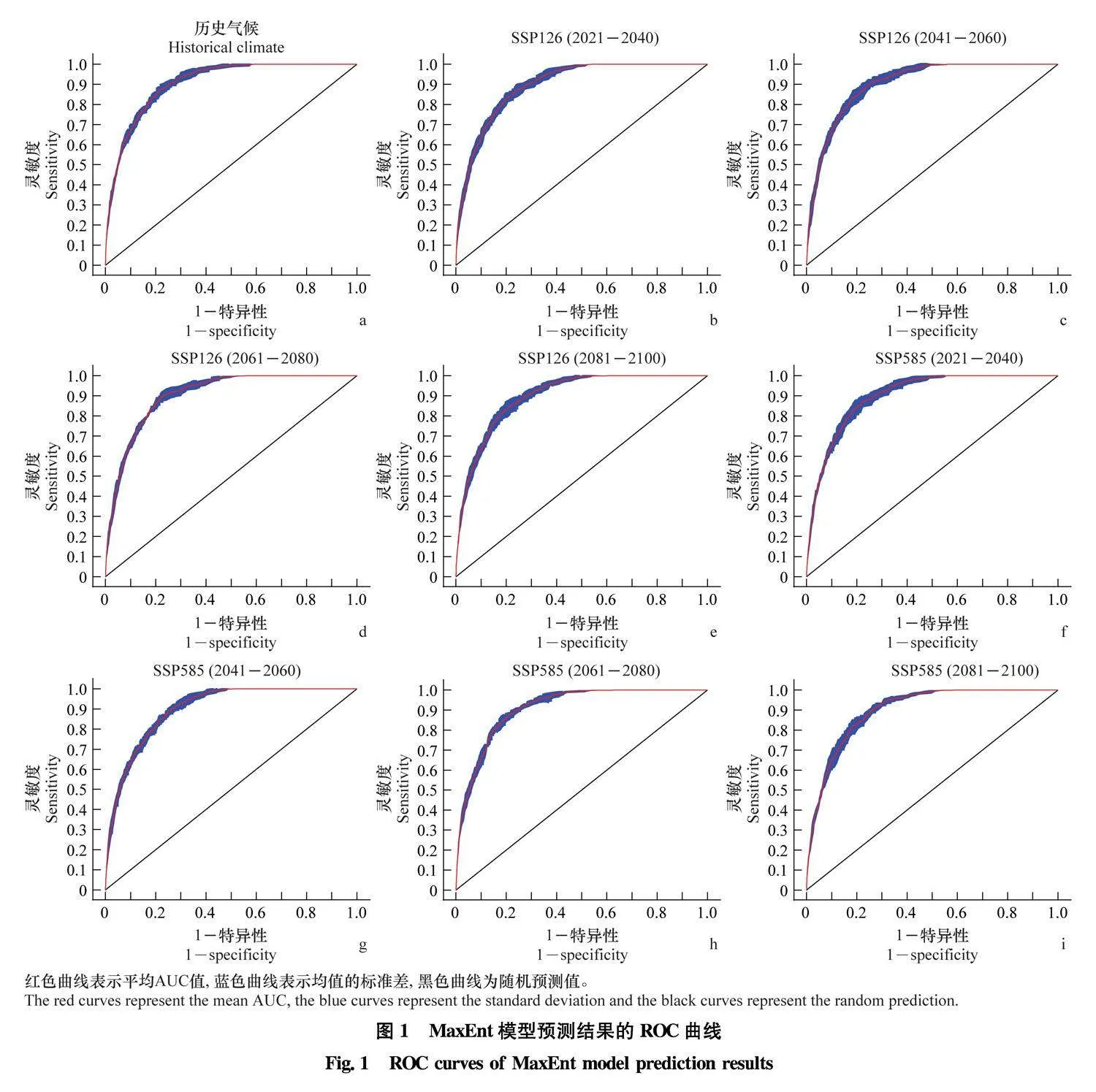
MaxEnt模型构建时重复运行10次,过去气候条件下AUC平均值达到0.907 (图1a)。对2021年-2040年、2041年-2060年、2061年-2080年、2081年-2100年这4个时期,SSP126情景模式下,AUC平均值分别为0.901、0.904、0.904、0.900 (图1b~e);SSP585情景模式下, 各模型AUC平均值分别为0.900, 0.900、0.909、0.901 (图1f~i), 说明模型构建优秀, 预测结果准确。
2.2" 历史气候条件适生性分析
2.2.1" 适生区分布
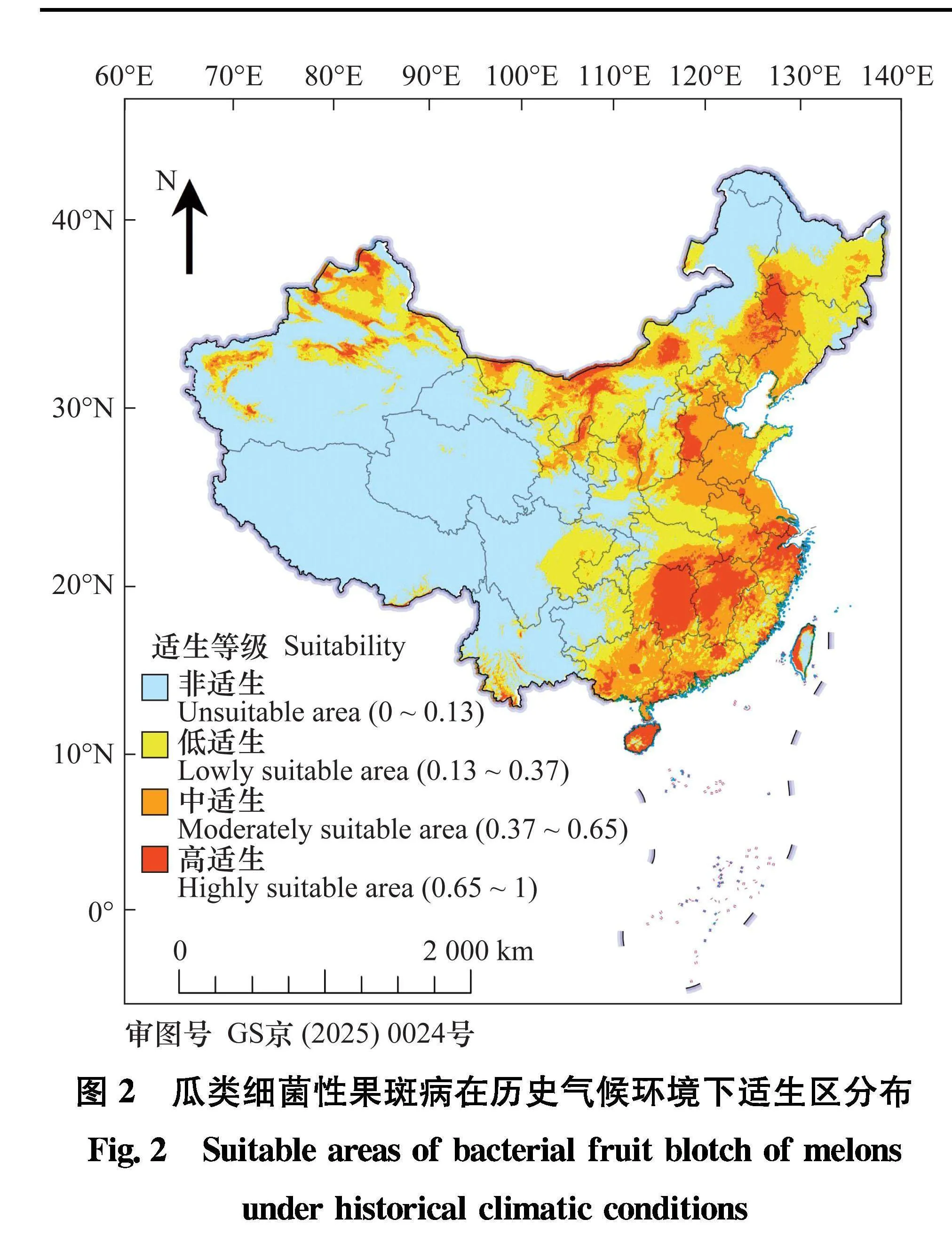
10次运行结果平均值的asc格式文件导入ArcGIS, 采用自然断点法(Jenks’ natural breaks)将适生区分为4类[36]。P≤0.13为非适生区, 用蓝色表示;0.13lt;P≤0.37为低适生区, 用黄色表示;0.37lt;P≤0.65为中适生区
, 用橙色表示;P>0.65为高适生区,用红色表示(图2)。模型结果显示,瓜类细菌性果斑病在我国适生范
围很广, 在ArcGIS
中计算适生区面积, 适生区面积占我国总面积的47.36%, 高适生区面积为66.95万km2, 中适生区为162.93万km2, 低适生区为225.59万km2。吉林、辽宁、河北、北京、天津、宁夏、河南、山东、江苏、上海、安徽、浙江、湖北、湖南、重庆、福建、江西、广东、广西、海南这些地区适生范围几乎遍布全境, 内蒙古、新疆、甘肃、陕西、台湾等地区也有较大范围的中、高适生区,非适生区主要分布在我国的西南地区和部分东北地区。
2.2.2" 主要气候因素分析
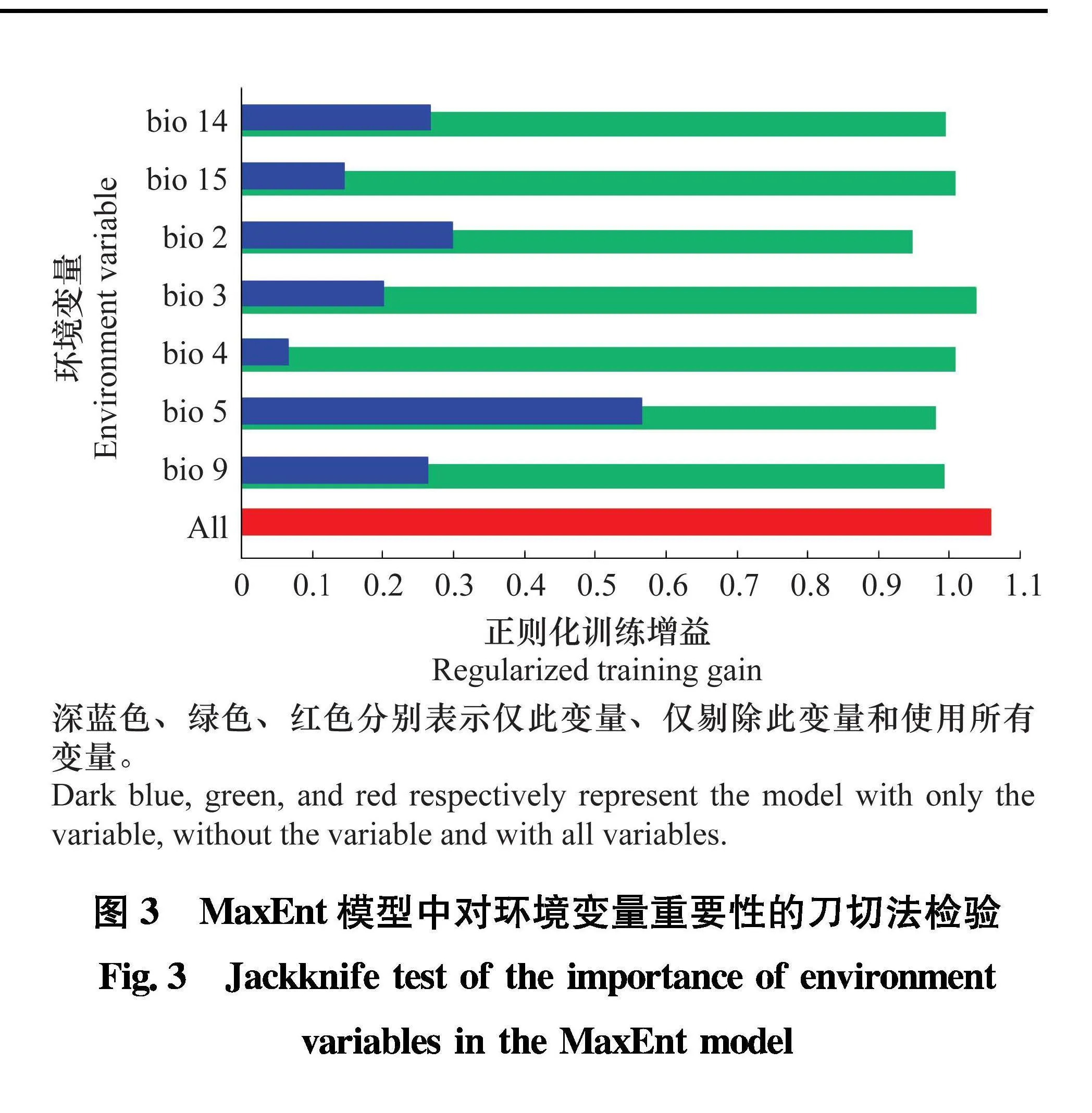
19个气候因子采用皮尔逊系数和变量贡献值筛选后, 剩余7个气候变量数据, 分别是: bio 2、bio 3、bio 4、bio 5、bio 9、bio 14和bio 15。对这7个
气候变量使用刀切法检验对模型构建的重要程度。如图3所示, 所有气候变量中最热月份最高温度(bio 5)训练增益值最高, 是模型构建最为重要的气候因子, 其次是月平均昼夜温差(bio 2)、最干月份降水量(bio 14)和最干季度平均温度(bio 9), 这4个变量是影响瓜类细菌性果斑病
在我国适生区分布的主导因素。在仅剔除bio 2
构建模型时, 条带最短说明这个气候因子拥有最多其他变量不存在的信息。
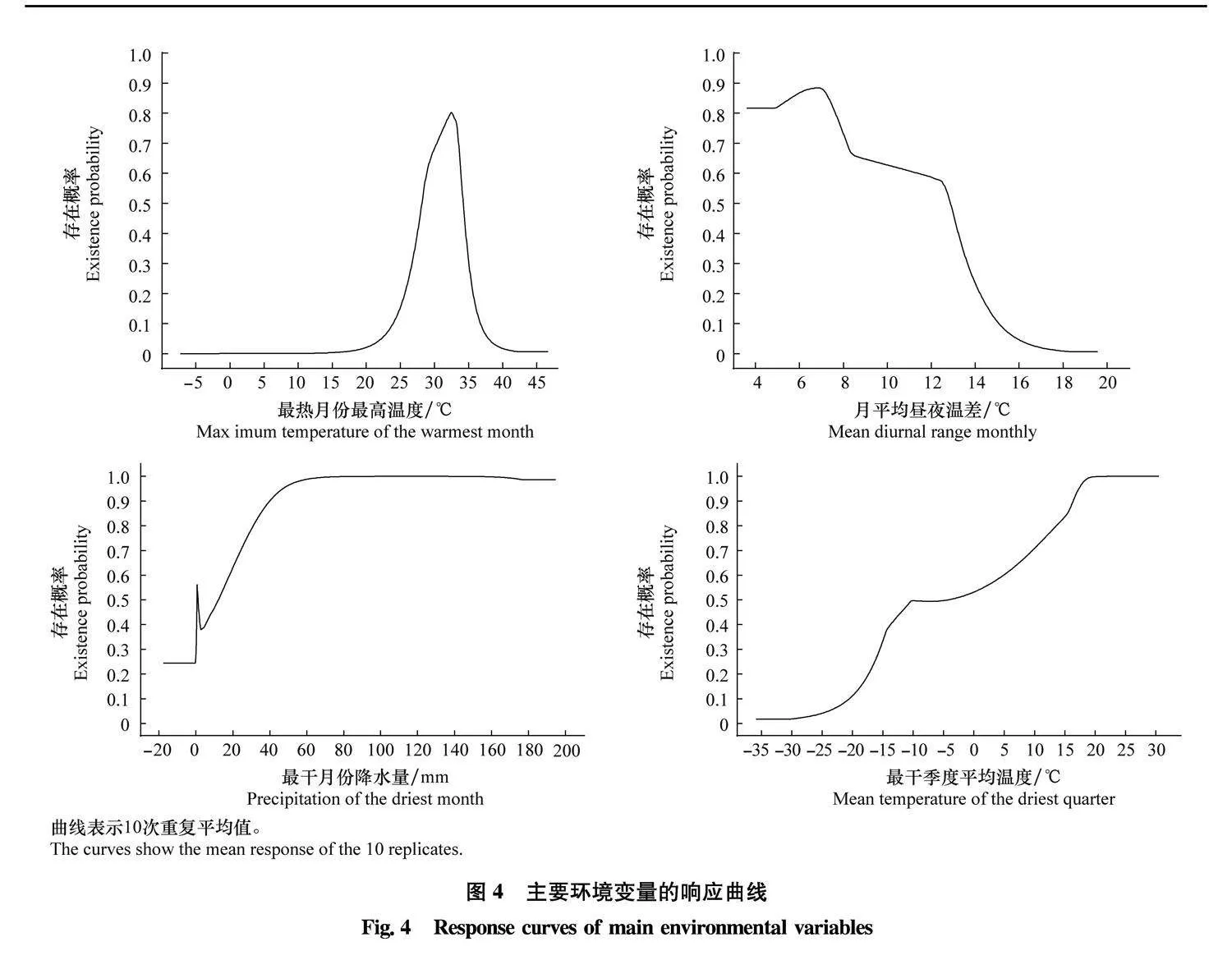
分别对这4个气候变量的响应曲线(图4)分析
可得: 最热月份最高温度(bio 5)在30~35℃时, 存
说明过高的气温使得病原菌不能存活;最适宜的月在
概率最高, 超过最适温度以后, 存在概率骤减,
平均昼夜温差(bio 2)在6~8℃, 大于8℃后分布概率随着月平均昼夜温差增加而减少;最干月份降水量(bio 14)越高, 分布概率越大, 当降水量大于60 mm后存在概率达到最高;分布概率随着最干季度平均温度(bio 9)的升高而升高, 并且当温度大于20℃后, 存在概率达到最高。
2.3" 未来气候情景模式下适生区变化
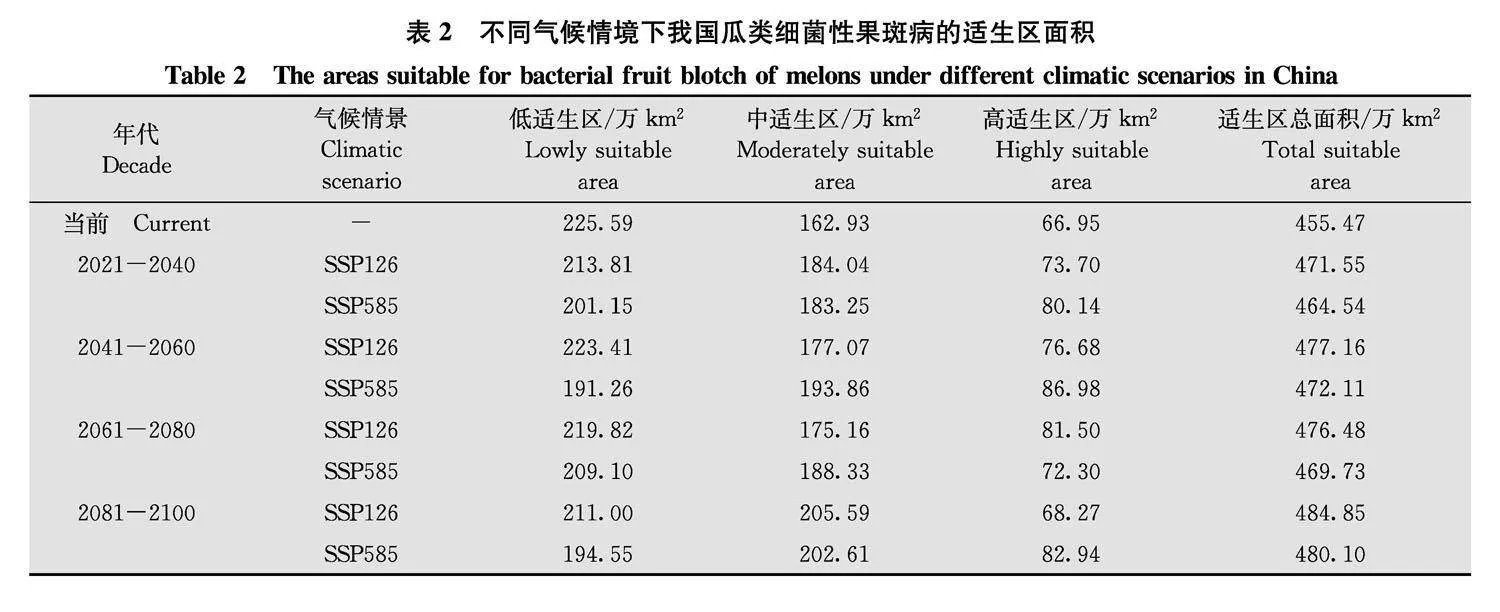
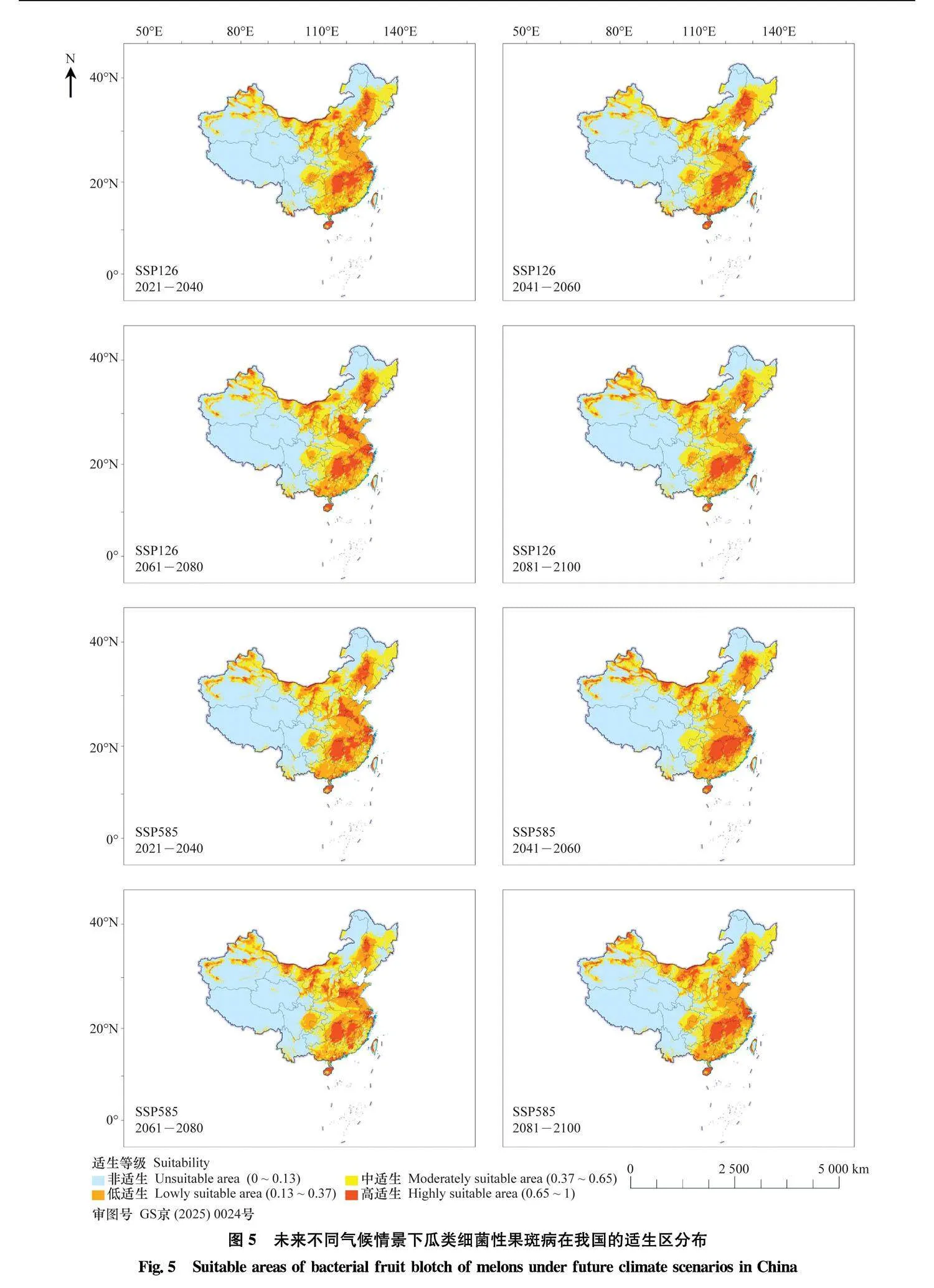
在SSP126和SSP585 2种未来气候情景下, 分别预测了2021年-2040年、2041年-2060年、2061年-2080年、2081年-2100年4个时间段里, 适生区的变化(图5), 并使用ArcGIS统计面积变化(表2)。
SSP126气候情景下,高适生区面积先增加后下降,2061年-2080年时最高,面积增长率达21.73%,
到2081年-2100年高适生区面积下降到当前水平。总适生面积基本呈现逐渐增加的趋势,到2081年-2100年时,总适生面积增加了29.38万km2。
在SSP585情景下,每一个时间段相较于当前气候模式,高适生区面积有所增加,并在
2041年-2060年时达到最高,增长率为29.92%;中适生区在2081年-2100年时面积最大,增长率为24.35%;低适生区相较于当下,面积都有所减少。总适生区面积在每个时间段与过去气候预测适生区相比都有所增加,2081年-2100年时最高,增加了24.63万km2。
3" 讨论与展望
随着社会的发展和进步,各国家、地区之间的贸易往来更加频繁,外来生物入侵的风险也随之增加[37],对外来入侵生物的风险分析在防控管理策略中显得尤为重要。瓜类细菌性果斑病作为一种我国发生面积广泛的外来入侵病害,每年对西甜瓜产业造成较大的经济损失。由于带菌种子是主要初侵染源,并且西甜瓜整个生育期都易受到侵染,对于调入疫情发生概率较高地区的种苗,需要严格检疫措施,并在植株整个生育过程中做好监控监测。MaxEnt模型因其准确性较高,但对分布点数据要求不高,常被用于适生性分析[38]。本研究主要利用MaxEnt模型与ArcGIS结合,使用历史气候和不同强迫场景下的未来气候,对瓜类细菌性果斑病在我国的适生区域进行了预测,预测结果能够为病害的科学防控和及时监测提供一定的帮助。
ROC曲线下面积即AUC值是MaxEnt精度检验的重要指标。ROC曲线根据一系列不同的二分类方式,分别以假阳性率(1-特异率)和真阳性率(1-遗漏率)为横纵坐标绘制而成,曲线下面积值即AUC值。由于MaxEnt模型在绘制ROC曲线时,是利用预测出的适生区占总面积比例作为评估特异
性的代替措施。因此,当适生面积较大时,计算所得特异性就会较差,即模型的AUC值会偏低。本研究
中预测出瓜类细菌性果斑病适生面积较大,占我国总面积47.36%,模型AUC值均大于0.9,说明构建的模型优秀,准确性较好。同时近5
年来,我国江苏省宿迁市和南通市,浙江省杭州市、丽水市、温州市,山东省临沂市、菏泽市、日照市,吉林省长春市、松原市、白城市,辽宁省盘锦市、丹东市等地出现新的瓜类细菌性果斑病疫情,与本研究所得的适生区预测结果基本相符,进一步佐证该模型的准确性。
模型预测呈现的适生区遍布我国大部分省份,仅西藏、青海等极少数西南地区适生面积较小。中、高适生区分布区域与我国西甜瓜主要产区(山东、河南、河北、江苏、新疆、内蒙古、浙江、湖南等)[39]重合度极高,病害发生后若不及时采取相应措施,将会对我国西甜瓜产业造成巨大损失,潜在适生区做好病害的防控对西甜瓜产业健康发展有重要意义。
对所有气候因子采用贡献率和皮尔逊系数结合进行筛选,最后得到7个气候因子并构建模型。刀切法显示,为模型构建提供较高增益的气候因子是:最热月份最高温度、月平均昼夜温差、最干月份降水量和最干季平均温度。最热月份最高温度低于30℃或高于15℃,最干月份降水量低于60 mm,月平均昼夜温差大于8℃,最干季平均温度低于15℃都会降低病害存在概率。对气候因子响应曲线的分析不难发现,该病害喜好高温高湿,昼夜温差较小的气候条件。分析结果与病原菌适宜生长温度[40]以及病害的适宜发病条件[41]等是符合的,进一步证明了模型预测的科学性、准确性。虽然当前多数西甜瓜产地由露天种植逐渐改为设施栽培[42],但由于棚内植株密集,环境高温高湿,并且主要采用喷灌设施,十分有利于病害的发生和扩散,因此,即使在气候并非适宜发病的情况下,也需要警惕果斑病的发生。
相较于CMIP5, CMIP6增加了社会经济发展方面的考虑,综合考虑了典型浓度排放路径(representative concentration pathway, RCP)和共享社会经济路径(shared socioeconomic pathway, SSP)[4344]。SSP一共有5种情景:SSP1~SSP5分别描述了5种社会经济发展趋势对未来气候的影响[45];RCP2.6、RCP4.5、RCP6.0、RCP8.5分别表示了不同的温室气体排放情景,其中RCP8.5导致的温度上升幅度最大,其次是RCP6.0和RCP4.5, RCP2.6对全球变暖的影响最小[46]。在未来气候变化情景下,无论是低环境强迫(SSP126)还是高环境强迫(SSP585),总适生区的面积都呈现增加的趋势,这也预示着随着气候的变化,果斑病在我国发生的潜在风险也会不断增加。中、高适生区覆盖的地区总体变化不大,所以即使在气候变化的情况下,疫情发生风险较高的地区在当下和未来都需要警惕病害的发生和蔓延。
研究采用的WorldClim 2.1数据库中过去气候仅包含1970年-2000年的数据,缺少最近20年的气候数据,且病害的分布不仅受气候影响,还有海拔、植被、人类活动等其他因素的影响。因此,本研究用于构建模型的数据还不够充分,预测结果也可能会有一定的偏差,后续应该综合考虑多种因素,进一步调整优化模型。瓜类细菌性果斑病适生范围极广,并且可以在西甜瓜的整个生育期发生,带来严重的经济损失,本研究的预测结果对相关部门制定相应的监测预警、防控策略,作物的生产管理规划等提供一定的参考,并为应对气候变化导致的适生区的变化提供数据支撑。
参考文献
[1]" SCHAAD N W, POSTNIKOVA E, SECHLER A, et al. Reclassification of subspecies of Acidovorax avenae as A.avenae (Manns 1905) emend., A.cattleyae (Pavarino, 1911) comb. nov., A.citrulli Schaad et al., 1978) comb. nov., and proposal of A.oryzae sp. nov. [J]. Systematic and Applied Microbiology, 2008, 31(6/7/8): 434446.
[2]" WEBB R E, GOTH R W. A seedborne bacterium isolated from watermelon [J]. Plant Disease Reporter, 1965, 49: 818821.
[3]" 胡俊, 黄俊霞, 刘双平, 等. 内蒙古哈密瓜细菌性果斑病的发生特点与防治技术[J]. 中国植保导刊, 2006(12): 1920.
[4]" 赵廷昌, 赵洪海, 王怀松. 山东省西瓜、甜瓜发生瓜类细菌性果斑病[J]. 植物保护, 2009, 35(5): 170171.
[5]" 蔡学清, 黄月英, 杨建珍, 等. 福建省西瓜细菌性果斑病的病原鉴定[J]. 福建农林大学学报, 2005, 34(4): 434437.
[6]" 任小平, 李小妮, 王琳, 等. 广东西瓜果斑病的病原鉴定[J]. 华南农业大学学报, 2010, 31(4): 4043.
[7]" 金岩, 张俊杰, 吴燕华, 等. 西瓜细菌性果斑病的发生与病原菌鉴定[J]. 吉林农业大学学报, 2004, 26(3): 263266.
[8]" TIAN Yanli, ZHAO Yuqiang, ZHOU Jiaju, et al. Prevalence of Acidovorax citrulli in commercial cucurbit seedlots during 2010-2018 in China [J]. Plant Disease, 2020, 140(1): 255259.
[9]" ELITH J, LEATHWICK J R. Species distribution models: ecological explanation and prediction across space and time [J]. Annual Review of Ecology Evolution and Systematics, 2009, 40(1): 677697.
[10]朱耿平, 刘国卿, 卜文俊, 等. 生态位模型的基本原理及其在生物多样性保护中的应用[J]. 生物多样性, 2013, 21(1): 9098.
[11]PETERSON A T. Ecological niche conservatism: a time-structured review of evidence [J]. Journal of Biogeography, 2011, 38(5): 817827.
[12]PHILLIPS S J, DUDIK M, SCHAPIRE R E. A maximum entropy approach to species distribution modeling [C]∥Proceedings of the 21th international conference on machine learning, 2004.
[13]PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions [J]. Ecological Modelling, 2006, 190(3/4): 231259.
[14]URBANI F, DALESSANDRO P, FRASCA R, et al. Maximum entropy modeling of geographic distributions of the flea beetle species endemic in Italy (Coleoptera: Chrysomelidae: Galerucinae: Alticini) [J]. Zoologischer Anzeiger, 2015, 258: 99109.
[15]赵宇, 王佳冠, 陈超, 等. 基于MaxEnt生态位模型柑橘木虱的云南潜在适生区研究[J].云南农业大学学报(自然科学), 2022, 37(1): 6168.
[16]杨鹄溶, 李恩, 徐玉梅, 等. 腐烂茎线虫在山西省的适生区预测[J]. 福建农林大学学报(自然科学版), 2022, 51(1): 3945.
[17]魏鹏, 张源, 何佳遥, 等. 基于MaxEnt模型分析气候变化下玉米褪绿斑驳病毒的潜在地理分布[J]. 植物保护学报, 2022, 49(5): 13671376.
[18]张秀芸, 伍文慧, 梁英梅. 落叶松枯梢病在中国的适生性分析[J]. 生态学报, 2024, 44(7): 30273037.
[19]吴卓瑾, 梁特, 石娟. 基于MaxEnt模型预测梨火疫病菌的潜在地理分布[J]. 植物保护学报, 2023, 50(6): 15181527.
[20]TIAN Yanli, ZHAO Yuqiang, WALCOTT R R, et al. Reliable and sensitive detection of Acidovorax citrulli in cucurbit seed using a padlock-probe-based assay [J]. Plant Disease, 2013, 97(7): 961966.
[21]赵子婧, 芦钰, 田文, 等. 应用微滴数字PCR同时检测瓜类种子携带果斑病菌和角斑病菌[J]. 植物保护, 2021, 47(2): 156163.
[22]杨玉文, 乔培, 刘德华, 等. 基于多重PCR技术的西瓜噬酸菌分组检测方法及其应用[J]. 植物保护, 2022, 48(4): 211219.
[23]TIAN Yanli, ZHAO Yuqiang, WU Xinrong, et al. The type VI protein secretion system contributes to biofilm formation and seed-to-seedling transmission of Acidovorax citrulli on melon [J]. Molecular Plant Pathology, 2015, 16(1): 3847.
[24]WANG Jianan, LIU Jun, ZHAO Yuqiang, et al. OxyR contributes to virulence of Acidovorax citrulli by regulating anti-oxidative stress and expression of flagellin FliC and type Ⅳ pili PilA [J/OL]. Frontiers in Microbiology, 2022, 13: 977281. DOI: 10.3389/FMICB.2022.977281.
[25]YANG Linlin, ZHAO Mei, ZHANG Xiaoxiao, et al. Acidovorax citrulli type Ⅲ effector AopU interferes with plant immune responses and interacts with a watermelon E3 ubiquitin ligase [J/OL]. Frontiers in Microbiology, 2023, 14: 1275032. DOI: 10.3389/FMICB.2023.1275032.
[26]LIU Dehua, ZHAO Mei, QIAO Pei, et al. ntrC contributes to nitrogen utilization, stress tolerance, and virulence in Acidovorax citrulli [J/OL]. Microorganisms, 2023, 11(3): 767. DOI: 10.3390/MICROORGANISMS11030767.
[27]林涛, 马国斌, 姜守阳, 等. 防治甜瓜细菌性果斑病种传病原菌的杀菌剂筛选[J]. 上海农业学报, 2023, 39(3): 5560.
[28]刘宝玉, 孙福庆, 杨玉文, 等. 化学药剂防控厚皮甜瓜细菌性果斑病的研究与应用[J]. 中国瓜菜, 2020, 33(9): 7478.
[29]白庆荣, 陈悦, 孙于淼, 等. 28个农药产品推荐剂量对西瓜噬酸菌的抑制活性[J]. 中国瓜菜, 2019, 32(2): 3235.
[30]辛晓歌, 吴统文, 张洁, 等. BCC模式及其开展的CMIP6试验介绍[J]. 气候变化研究进展, 2019, 15(5): 533539.
[31]刘婷, 曹家豪, 齐瑞, 等. 基于GIS和MaxEnt模型分析气候变化背景下紫果云杉的潜在分布区[J]. 西北植物学报, 2022, 42(3): 481491.
[32]RINNHOFER L J, ROURA-PASCUAL N, ARTHOFER W, et al. Iterative species distribution modelling and ground validation in endemism research: an alpine jumping bristletail example [J]. Biodiversity and Conservation, 2012, 21(11): 28452863.
[33]PHILLIPS S J, ANDERSON R P, DUDK M, et al. Opening the black box: an open-source release of MaxEnt [J]. Ecography, 2017, 40(7): 887893.
[34]HANLEY J A, MCNEIL B J. The meaning and use of the area under a receiver operating characteristic (ROC) curve [J]. Radiology, 1982,143(1): 2936.
[35]SWETS J A. Measuring the accuracy of diagnostic systems [J]. Science, 1988, 240(4857): 12851293.
[36]林伟, 徐淼锋, 权永兵, 等. 基于MaxEnt模型的草地贪夜蛾适生性分析[J]. 植物检疫, 2019, 33(4): 6973.
[37]ESCHEN R, BRITTON K, BROCKERHOFF E, et al. International variation in phytosanitary legislation and regulations governing importation of plants for planting [J]. Environmental Science and Policy, 2015, 51: 228237.
[38]汤思琦, 武扬, 梁定东, 等. 未来气候变化下栎树猝死病菌在中国的适生性分析[J]. 生态学报, 2023, 43(1): 388397.
[39]王娟娟, 李莉, 尚怀国. 我国西瓜甜瓜产业现状与对策建议[J]. 中国瓜菜, 2020, 33(5): 6973.
[40]WILLEMS A, GOOR M, THIELEMANS S, et al. Transfer of several phytopathogenic Pseudomonas species to Acidovorax as Acidovorax avenae subsp. avenae subsp. nov., comb. nov., Acidovorax avenae subsp. citrulli, Acidovorax avenae subsp. cattleyae, and Acidovorax konjaci [J]. International Journal of Systematic Bacteriology, 1992, 42(1): 107119.
[41]赵廷昌, 孙福在, 王兵万. 西瓜细菌性果斑病研究进展[J]. 植保技术与推广, 2001(3): 3738.
[42]吴敬学, 赵姜, 张琳. 中国西甜瓜优势产区布局及发展对策[J]. 中国蔬菜, 2013(17): 15.
[43]RIAHI K, VAN VUUREN D P, KRIEGLER E, et al. The shared socioeconomic pathways and their energy, land use, and greenhouse gas emissions implications: An overview [J]. Global Environmental Change, 2017, 42: 153168.
[44]GEORGE C H, LOUISE C, RITVIK S, et al. Harmonization of global land use change and management for the period 8502100 (LUH2) for CMIP6 [J]. Geoscientific Model Development, 2020, 13(11): 54255464.
[45]ALEXANDER P, KATHERINE C, SHINICHIRO F, et al. Land-use futures in the shared socio-economic pathways [J]. Global Environmental Change, 2017, 42: 331345.
[46]HURTT G C, CHINI L P, FROLKING S, et al. Harmonization of land-use scenarios for the period 1500-2100: 600 years of global gridded annual land-use transitions, wood harvest, and resulting secondary lands [J]. Climatic Change, 2011, 109(1/2):117161.
(责任编辑:杨明丽)
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
今日农业(2022年15期)2022-09-20 06:55:22
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
竹子学报(2018年2期)2018-10-25 02:02:12
吉林蔬菜(2018年3期)2018-03-12 05:35:17
吉林蔬菜(2017年10期)2017-11-01 07:47:04
吉林蔬菜(2017年10期)2017-11-01 07:47:04
吉林蔬菜(2017年7期)2017-07-18 12:09:54
现代园艺(2017年11期)2017-06-28 11:32:18
