
作物多样性(轮作)下砖红壤溶解性有机质含量及光谱特征
2025-02-16 00:00:00陈心怡刘钰钦陈淼王文峰
农业资源与环境学报 2025年1期
关键词:轮作

摘要:本研究依托海南儋州热带农业生态系统国家野外科学观测研究站长期定位试验,设置玉米连作(单作物)、辣椒+玉米轮作(双作物)、辣椒+玉米/绿肥套种(三作物)和辣椒/玉米套种+豆角/绿肥套种(四作物)4种处理,利用紫外吸收光谱、三维荧光光谱结合平行因子法考察作物多样性(轮作)对研究区砖红壤溶解性有机质(DOM)含量及光谱特征的影响。结果显示,相比于单作物连作,增加作物多样性的轮作处理土壤有机质含量和持水率增加,但DOM含量降低。不同处理土壤DOM由可见光区类富里酸(Ex/Em=355 nm/432 nm,C1)、类腐植酸(Ex/Em=425 nm/471 nm,C2)和类蛋白(Ex/Em=325 nm/336 nm,C3)3种荧光组分组成,组分C1对DOM荧光组分贡献最高(47%~76%)。相比于单作物连作,增加作物多样性的轮作处理土壤DOM的C1组分相对丰度增加,而C2和C3相对丰度降低。光谱特征参数结果显示,单作物连作处理土壤DOM的芳香度(UV254、SUVA254)、腐殖化程度(HIX)及分子量(E2/E3、光谱斜率比SR)高于作物多样性轮作处理土壤。豆科作物的引入降低了三作物和四作物轮作处理土壤DOM的自生源特征。HIX与C1组分呈显著正相关(R2=0.96,Plt;0.05),表明土壤腐殖化程度主要由类富里酸物质贡献。研究结果可为热带地区土壤农业种植模式的构建和优化提供理论依据。
关键词:作物多样性;轮作;砖红壤;溶解性有机质;光谱分析
中图分类号:S153.6;S344 文献标志码:A 文章编号:2095-6819(2025)01-0149-10 doi: 10.13254/j.jare.2023.0627
尽管现代集约化农业为满足粮食安全作出了巨大贡献,但其所带来的资源环境代价与粮食安全及可持续发展之间的矛盾日益突出。因此,在确保国家粮食安全的同时,迫切需要构建可持续的作物生产系统,实现我国农业的绿色转型发展[1]。作物轮作是指在同一块田地上,有顺序地在季节间或年间轮换种植不同的作物或复种组合的一种种植方式[2]。而多样化轮作制度作为现代农业可持续发展的重要农业技术措施,能够在提高土壤肥力、增加农田生态系统稳定性的同时,实现地力保育、丰产稳产、资源高效利用等多目标协同,目前已成为世界范围内实现农业健康绿色可持续发展的重要途径[3]。
已有大量研究表明,作物轮作可增加农田土壤有机质含量,且土壤有机质含量的增加与作物多样性呈正相关[4]。土壤溶解性有机质(Dissolved OrganicMatter,DOM)作为土壤有机质中最重要、最活跃的组成部分,是土壤微生物可直接利用的有机质源,在土壤一系列生物化学过程中发挥着重要作用,对作物类型及耕作栽培方式等农艺措施具有敏感反应[5]。然而,目前关于作物多样性轮作对土壤碳库的影响研究主要集中在土壤总有机质含量,而对土壤DOM含量、组成分布及结构特征的影响关注较少。DOM 的组成、结构特征、芳香性和腐殖化程度会影响相关环境中污染物的迁移转化,进而影响其相关的地球化学循环[6-7]。因此,明确作物多样性轮作对土壤DOM含量、组成分布及结构特征的影响,对理解作物多样性轮作对土壤DOM动态变化及其相关生态环境效益的影响具有重要意义。
鉴于此,本研究依托海南儋州热带农业生态系统国家野外科学观测研究站(简称“儋州站”)长期定位试验,设置玉米连作(单作物)、辣椒+玉米轮作(双作物)、辣椒+玉米/绿肥套种(三作物)和辣椒/玉米套种+豆角/绿肥套种(四作物)四种处理(施肥等管理措施均保持一致),考察作物多样性(轮作)对热带地区砖红壤DOM含量的影响,并结合紫外-可见光谱、三维荧光光谱和平行因子分析法,系统研究长期作物多样性轮作下土壤DOM结构组成、来源及腐殖化特征,以期为热带地区土壤农业种植模式的构建和优化提供理论依据。
1 材料与方法
1.1 试验地概况
本研究中的试验田位于海南省儋州市那大镇宝岛新村(19°31′ N,109°34′ E),其所在地区属于热带季风气候,年平均温度23.5 ℃,年降雨量1 815.0 mm,土壤为花岗岩母质风化所形成的砖红壤,质地为砂壤土。2017年开展作物多样性轮作时土壤pH为5.9;氧化还原电位为112.25 mV;全氮含量为0.93 g·kg-1;全磷含量为0.56 g·kg-1;全钾含量为46.46 g·kg-1。
1.2 试验设计
本研究选取辣椒(丰椒十八)、玉米(丰都八号)、豆角(新优丰特长豆角)、绿肥(硬皮豆)4种热带地区典型作物,四种轮作处理均进行底肥条施、覆膜的种植管理措施。试验田均施复合肥(N、P2O2、K2O 比例为15∶15∶15),施肥量分别为N 345 kg·hm-2、P2O2 273kg·hm-2、K2O 417 kg·hm-2,每个试验田(7.5 m×8 m)有6垄。每个处理设置三个平行,为了更好地进行后续统计学分析(PARAFAC 分析要求样本量不少于20,且样本量越大越好)[8],每块样地采用五点取样法采集耕层土壤(0~20 cm)样品3个,最终共采集36个土壤样品,所有土壤样品混匀后立即带回实验室,预处理后置于冰箱4 ℃储存备用。
1.3 测定项目与方法
1.3.1 土壤理化性质
将预处理后的土壤样品,分别采用电位法和电导率法测定土壤酸碱度(pH)和电导率(EC),重铬酸钾容量法(外加热法)测定土壤有机碳含量[9]。
1.3.2 土壤DOM提取
以水土比5∶1混合并采用振荡法提取土壤DOM溶液,用总有机碳/氮分析仪(Multi N/C 3100/1,耶拿分析仪器公司,德国)测定土壤DOM中的溶解性有机碳(DOC)含量。
1.3.3 土壤DOM的光谱分析
DOM 样品的紫外-可见吸收光谱通过紫外分光光度计(UV2600,岛津公司,日本)扫描获得。三维荧光光谱测定利用F97PRO 荧光分光光度计进行。三维荧光光谱的扫描结果经空白扣除、内滤效应校正及拉曼散射校正后进行数据分析。
1.4 平行因子分析
平行因子分析(Parallel Factor Analysis,PARAFAC)是在三线性理论的基础上用交替最小二乘法实现的统计分析方法,用来表征DOM中组分的荧光特征并提供溯源信息[10]。本研究利用DOMFlour 工具箱在MATLAB R2021a 中对DOM 样品的三维荧光光谱数据进行PARAFAC分析,并通过了拆半分析检验。
1.5 数据处理与分析
本研究中所有试验数据分析及制图采用的软件为Excel、Origin 2021、JMP Pro 14。
2 结果与分析
2.1 不同轮作模式下土壤理化性质特征
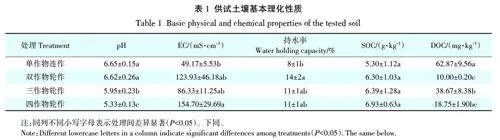
由表1可知,在不同轮作模式下土壤理化性质存在显著差异。与单作物连作处理土壤(pH=6.65)相比,多作物轮作降低了土壤pH(5.33~6.62),且作物种类越多,土壤pH值越低。轮作作物种类的增加可显著增加土壤EC和土壤持水率。
通过轮作增加作物多样性可以增加SOC 含量。双作物、三作物和四作物轮作处理的SOC含量相比单作物轮作分别提高18.9%、20.6% 和30.8%。而作为反映微生物活性最强的有机碳库,DOC 的含量变化趋势与SOC 不同。单作物连作土壤中DOC 含量最高,而通过轮作增加作物多样性的土壤DOC含量均显著下降。
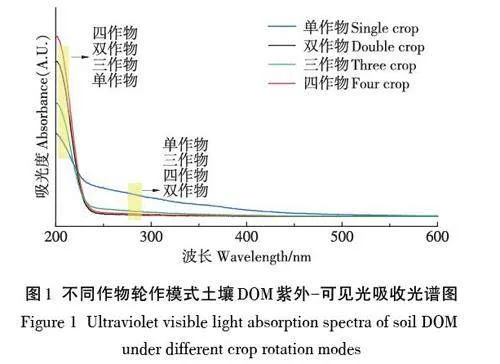
2.2 紫外光谱分析
紫外-可见光谱具有灵敏度高、所需样品量较少等优点,其光谱特征及相关参数通常用来表征土壤DOM 腐殖化程度、组成结构和来源[11-12]。如图1 所示,四种不同处理土壤DOM样品的吸收曲线均表现为吸光度随波长增加在200~240 nm之间迅速下降,随后逐渐降低并趋于0,但不同DOM的吸收曲线特征也存在一定差异。在200~225 nm波段上,不同DOM吸收曲线吸光强度大小顺序为四作物轮作gt;双作物轮作gt;三作物轮作gt;单作物连作,而在225 nm之后的波段上,不同DOM吸收曲线吸光强度大小顺序变为单作物连作gt;三作物轮作gt;四作物轮作gt;双作物轮作。
UV254 的数值变化可表征芳香性结构和DOM 的浓度[13]。由表2可知,单作物连作模式下土壤DOM浓度及其芳香性相对于多作物轮作模式更高,DOM 的芳香性结构或芳香性物质更多,这与紫外吸收光谱特征的结果吻合。
SUVA254为UV254和DOC浓度的比值,通常用来表征DOM 的芳香性结构和腐殖化程度[14-15]。由表2可知,不同处理土壤DOM的SUVA254大小顺序依次为单作物连作gt;双作物轮作gt;三作物轮作=四作物轮作。与UV254分析结果相似,单作物连作土壤DOM芳香性结构更多且腐殖化程度更高。
E2/E3 为波长254 nm 和365 nm 处吸光度比值,其大小与DOM分子量呈负相关关系[16]。由表2可知,不同处理土壤DOM的E2/E3值大小顺序为三作物轮作gt;双作物轮作gt;四作物轮作gt;单作物连作,表明单作物连作土壤DOM分子量最大,三作物轮作土壤DOM分子量最小。
E3/E4 为波长300 nm 和400 nm 处吸光度比值,通常用来表征DOM来源,其值gt;3.5表示DOM以富里酸为主,lt;3.5以胡敏酸为主[13]。由表2可知,三作物轮作处理土壤DOM的E3/E4值最高且大于3.5,其DOM以富里酸为主。而单作物连作、双作物轮作和四作物轮作处理土壤DOM的E3/E4值均小于3.5,以胡敏酸为主[17]。
光谱斜率比SR 为波长275~295 nm 与波长350~400 nm的积分面积比,该值与DOM浓度无关,通常用来表征DOM结构变化,与分子量呈反比[18]。由表2可知,SR值变化趋势与E2/E3相似,单作物连作处理土壤DOM分子量最大。
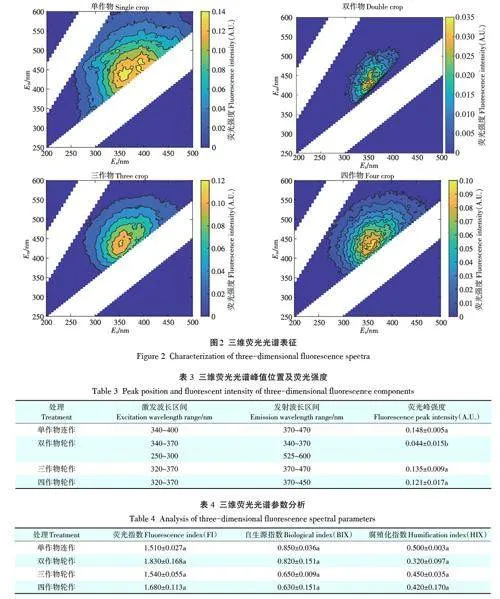
2.3 三维荧光光谱分析
不同轮作模式处理下土壤样品DOM的三维荧光光谱如图2所示,光谱峰位置如表3所示。轮作作物多样性的增加导致DOM荧光强度发生了明显变化。三维荧光光谱结果表明,所有处理土壤样品DOM主要出现一个类腐植酸峰[19],该峰在不同轮作处理下峰强度存在显著差异:单作物连作gt;三作物轮作gt;四作物轮作gt;双作物轮作,该变化趋势与紫外-可见光谱220 nm 后的吸收曲线变化一致,然而与200~225 nm之间的吸收曲线变化趋势不一致,这可能是由于DOM样本中不同硝酸盐含量影响所致[20]。
荧光指数(FI)可以反映环境中DOM 来源情况,FIgt;1.9为微生物内源,FIlt;1.4为陆地外源输入[21]。由表4 可知,本研究所有处理样品DOM 的FI 值均在1.4~1.9之间,表明土壤DOM为内源微生物与外源输入共同贡献。与其他处理土壤相比,双作物轮作处理土壤更接近1.9,内源特征明显。自生源指数(BIX)通常用来表征环境中DOM的自身来源贡献情况,BIXlt;0.8表示生物源特征不显著,0.8随着作物多样性的增加,土壤DOM腐殖化程度逐渐降低,其中双作物轮作处理土壤腐殖化程度降低最为明显。
2.4 平行因子分析
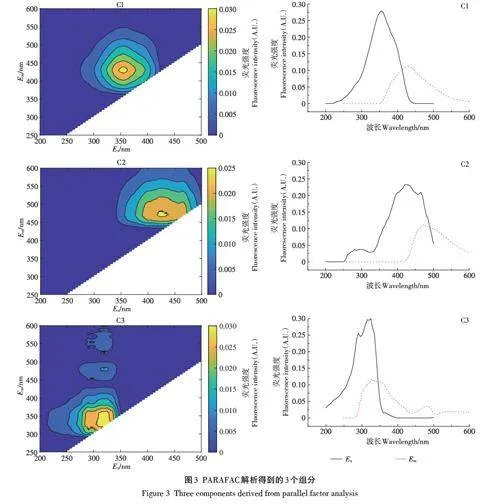
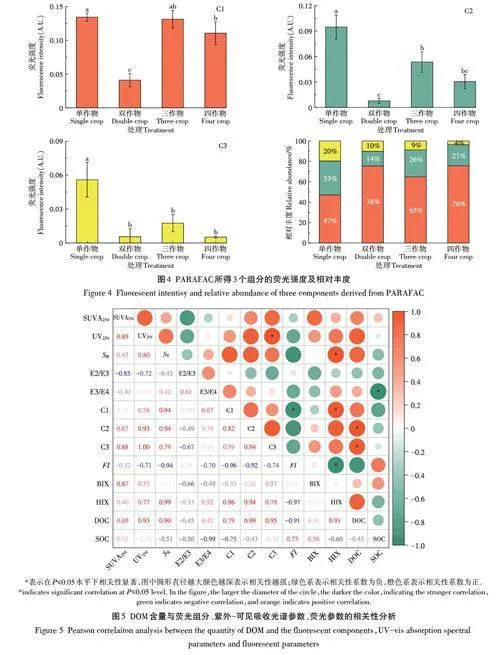
对所有种植模式下土壤DOM的三维荧光光谱数据进行平行因子(PARAFAC)分析,解析出C1、C2和C3 3个组分(图3)[22-23]:C1荧光峰位于Ex/Em=355 nm/432 nm,属于可见光区类富里酸峰;C2荧光峰位于Ex/Em=425 nm/471 nm,属于陆源类腐植酸峰;C3荧光峰主要位于Ex/Em=325 nm/336 nm,属于含有类色氨酸和类络氨酸的类蛋白峰。不同轮作处理土壤DOM样品平行因子组分荧光强度及相对含量变化如图4所示。紫外可见光类富里酸组分C1的荧光强度最大,其次为类腐植酸组分C2,类蛋白组分C3含量最低。作物多样性的增加,使3个荧光组分均呈下降趋势,但不同处理间下降趋势存在显著差异。随着C2和C3组分荧光强度的显著降低,其相对丰度也降低,而C1组分相对丰度显著增加。
2.5 土壤DOM与其光谱特征参数指标的相关性
对不同处理土壤的DOM 光谱特征参数进行Pearson相关性分析,结果如图5所示。在4种不同的轮作模式土壤中,类蛋白组分C3与UV254呈显著正相关(R2=1.00,P≤0.05);腐殖化系数HIX与SR(R2=0.99,P≤0.05)和可见光区类富里酸组分C1(R2=0.96,P≤0.05)均呈显著正相关,而与荧光指数FI呈显著负相关(R2=-0.97,P≤0.05);土壤DOC与陆源类腐植酸组分C2(R2=0.99,P≤0.05)和类蛋白组分C3(R2=0.95,P≤0.05)呈显著正相关;可见光区类富里酸组分C1与荧光指数FI呈负相关性(R2=-0.96,P≤0.05);荧光指数FI与腐殖化系数HIX呈负相关性(R2=-0.97,P≤0.05);E3/E4与土壤SOC呈显著负相关(R2=-0.99,P≤0.05)。
3 讨论
土壤SOC在促进土壤质量和功能、缓解土壤退化和减缓气候变化等方面发挥着重要作用,其含量极易受种植制度、轮作模式、施肥等农艺措施的影响。采用合理的轮作模式和规范的农田管理措施可以提高农田土壤固碳水平[24]。本试验结果表明,相比于单作物连作,增加作物多样性的轮作可显著增加土壤SOC的固持,该结果与前人研究结果相似[25]。更高的作物多样性轮作通过增加光合强度提高了初级生产力,促进地上及地上—地下相互作用,进而加强土壤功能和生物元素网络(植物输入和土壤食物网功能)和非生物因素(土壤基质和微环境)之间的相互作用,最终促进SOC积累[25]。作物多样性种植,尤其是豆科作物的引入,可增加土壤作物衰老根系和结节,提升总氮的储量,强化微生物碳的限制,进而刺激碳获取酶活性的增加,以催化降解多样化种植模式下的作物残茬,进而提升土壤碳储量[26]。此外,豆科作物残留物较低的碳氮比更容易被微生物利用,进一步降低了多样化种植模式下的SOC分解。豆科绿肥填闲作物的引入可增强土壤微生物活性和微生物衍生的结合剂(例如球囊霉素)的产生,进一步增强土壤团聚体稳定性,减少微生物对有机碳的接触和矿化,进而增加有机碳储量[27-28]。土壤团聚体稳定性的增加,提高了水分渗透和保留能力(表1),尤其是玉米+辣椒双作物轮作系统,这与前人研究结果一致[29]。这主要是由于双作物轮作条件下,粗根植物玉米与浅根植物辣椒的根系空间上合理搭配,增加了土壤的孔隙度,进而增加了土壤持水性能。土壤孔隙度及持水性能的增加,可进一步改善土壤中空气、水和溶质的空间和时间分布,从而影响根系和微生物呼吸以及幼苗出苗,对提高土壤质量至关重要。
相比于SOC,DOM 虽然在土壤有机碳库中占比很小,但其为土壤有机碳最活跃的组分,参与了土壤有机质腐殖化、迁移、微生物代谢等一系列生物地球化学过程,能够快速而灵敏地反映土壤有机质的动态,对全球碳循环和物质能量循环具有重要作用[30-31]。DOM的含量及其化学组成是决定其在土壤生态系统中功能和行为的重要因素。本研究结果显示,相比于玉米连作土壤,引入辣椒、豆角、绿肥等作物的轮作处理使土壤DOM的含量呈下降趋势,其中双作物和四作物轮作处理土壤DOM含量下降趋势更为明显。祖艳群等[32]的研究表明,玉米/辣椒系统有效促进了作物对氮、磷和钾等营养元素的吸收,进而促进了作物对土壤中易被利用碳源DOM的消耗。吴炯等[29]研究还发现,玉米/辣椒系统增加了土壤孔隙度,使得淋溶水与土壤颗粒的接触面积增加,进一步促进了土壤DOM的下渗淋失。尽管多样化的作物轮作增加了新的碳输入,但耕作、施肥等可以限制有机碳的积累。碳输入可能会通过以下机制刺激土壤DOM 的分解[25]:①增加R策略者的活性,土壤酶活性增加会导致DOM分解;②增加适应K策略者对底物的分解;③更多的根系分泌物(如草酸)破坏有机矿物的稳定性;④碳输入的化学计量学变化产生正激发效应,进而加速DOM分解。总而言之,作物多样性对土壤DOM的影响归因于各种因素,包括生物量生产、初始土壤碳水平、作物多样性轮作时间、土壤质地、气候等[33],这些影响因素的综合作用可能导致不同地区作物多样性轮作下土壤DOM变化趋势不一致。
由于作物多样性改变了植物营养物质的输入、植物凋落物、根系分泌物等数量和种类,进而改变DOM的来源,影响了土壤有机碳的固定过程及微生物活性,使得DOM的组成和性质在不同轮作处理下存在显著差异[34]。本研究结果显示,HIX与C1组分呈显著正相关,表明土壤DOM腐殖化程度主要由C1组分贡献(图5)。与玉米单作相比,轮作土壤DOM 的芳香性、腐殖化程度及分子量降低。张洪勇[35]的研究表明,与玉米单作相比,玉米/辣椒轮作系统可显著提升辣椒及玉米根际微生物对聚合物类、酚酸类、羧酸类等大分子碳源的利用,而显著降低对糖类、氨基酸、胺类等小分子碳源的利用。而随着豆科作物的引入,由于其根质量较低,产生的大量凋落物碳氮比低,分解速度快[36],进而降低了土壤DOM的芳香性、腐殖化程度及分子量,这也使得三作物和四作物轮作处理土壤DOM的自生源特征不显著。但是相比于玉米/辣椒轮作处理土壤,引入豆科绿肥作物的三作物和四作物轮作土壤腐殖化程度增加(表4),这可能是豆科植物根系分泌的腐植酸及其产生的凋落物腐解所致[36]。
4 结论
(1)增加作物多样性的轮作模式可以提升土壤SOC含量,且土壤SOC含量与作物多样性呈正相关。然而,作为土壤系统中生物活性及反应性最强的有机碳库,轮作处理下土壤DOM的含量较单作物连作处理土壤降低,且不同轮作模式下土壤DOM含量降低趋势不同。
(2)运用平行因子分析方法解析了不同轮作处理模式下土壤样品DOM的3种组分,分别为类富里酸、类腐植酸和类蛋白,组分类富里酸占比最高,类腐植酸次之,类蛋白占比最小。相比于单作物连作处理,轮作处理增加了土壤中类富里酸组分相对丰度,降低了类腐植酸和类蛋白组分相对丰度。
(3)光谱特征参数分析结果表明,相比于单作物连作处理土壤,轮作处理土壤DOM的芳香性、腐殖化程度及分子量降低。随着豆科作物的引入,土壤DOM 的自生源特征在双作物轮作处理下显著降低,在三作物和四作物轮作处理下不显著,而腐殖化程度增加。组分类富里酸与腐殖化程度呈显著正相关,表明其对DOM的腐殖化程度贡献较大。
参考文献:
[1] 吕送军, 雷云飞. 中国农业的现代化与可持续发展[J]. 湖南农机,
2008, 35(7):51-52. LÜ S J, LEI Y F. On modernization and the
sustainable development of Chinese agriculture[J]. Hunan Agricultural
Machinery, 2008, 35(7):51-52.
[2] 李银水, 余常兵, 廖星, 等. 湖北省不同油菜轮作模式下作物施肥现
状调查[J]. 中国农学通报, 2012, 28(36):205-211. LI Y S, YU C
B, LIAO X, et al. Investigation of present fertilization on crops by
different rapeseed rotation systems in Hubei Province[J]. Chinese
Agricultural Science Bulletin, 2012, 28(36):205-211.
[3] ZHAO J, CHEN J, BEILLOUIN D, et al. Global systematic review with
meta-analysis reveals yield advantage of legume-based rotations and
its drivers[J]. Nature Communications, 2022, 13(1):4926.
[4] LIU X T, TAN S W, SONG X J, et al. Response of soil organic carbon
content to crop rotation and its controls:a global synthesis[J].
Agriculture, Ecosystems amp; Environment, 2022, 335:108017.
[5] D’ANDRILLI J, SILVERMAN V, BUCKLEY S, et al. Inferring
ecosystem function from dissolved organic matter optical properties:a
critical review[J]. Environmental Science amp; Technology, 2022, 56(16):
11146-11161.
[6] CHEN M L, HUR J. Pre-treatments, characteristics, and
biogeochemical dynamics of dissolved organic matter in sediments:a
review[J]. Water Research, 2015, 79:10-25.
[7] LEYVA D, USMAN T M, JAFFÉ R, et al. Description of dissolved
organic matter transformational networks at the molecular level[J].
Environmental Science amp; Technology, 2023, 57(6):2672-2681.
[8] STEDMON C A, BRO R. Characterizing dissolved organic matter
fluorescence with parallel factor analysis:a tutorial[J]. Limnology and
Oceanography:Methods, 2008, 6(11):572-579.
[9] 鲍士旦. 土壤农化分析[M]. 北京:中国农业出版社, 2000. BAO S
D. Soil agrochemical analysis[M]. Beijing:China Agriculture Press,
2000.
[10] 梁以豪, 倪才英, 刘星星, 等. 不同稻虾种养模式土壤溶解性有机
质光谱特征[J]. 中国生态农业学报(中英文), 2023, 31(4):543-
557. LIANG Y H, NI C Y, LIU X X, et al. Spectral characteristics of
soil dissolved organic matter in different rice - crayfish cultivation
modes[J]. Chinese Journal of Eco-Agriculture. 2023, 31(4):543-557.
[11] ZHANG S Q, YIN Y G, YANG P J, et al. Using the end-member
mixing model to evaluate biogeochemical reactivities of dissolved
organic matter(DOM):autochthonous versus allochthonous origins[J].
Water Research, 2023, 232:119644.
[12] 田翔, 何天容, 尹德良, 等. 土壤溶解性有机质结构和组成对秸秆、
牛粪及其堆肥产品输入的响应特征[J]. 农业资源与环境学报,
2022, 39(3):556 - 566. TIAN X, HE T R, YIN D L, et al.
Responses of soil dissolved organic matter properties to the
amendment of straw, cow manure, or their composting products[J].
Journal of Agricultural Resources and Environment, 2022, 39(3):
556-566.
[13] 申慧彦, 王嘉富, 曹承泽, 等. 巢湖湖滨带不同土地类型土壤DOM
光谱研究[J]. 长江流域资源与环境, 2023, 32(4):729 -738.
SHEN H Y, WANG J F, CAO C Z, et al. Study on DOM spectra of
different soil types in Chaohu lakeside zone[J]. Resources and
Environment in the Yangtze Basin, 2023, 32(4):729-738.
[14] WEISHAAR J L, AIKEN G R, BERGAMASCHI B A, et al.
Evaluation of specific ultraviolet absorbance as an indicator of the
chemical composition and reactivity of dissolved organic carbon[J].
Environmental Science amp; Technology, 2003, 37, 20:4702-4708.
[15] 赵雄威, 吴东明, 李勤奋, 等. 基于紫外-可见光光谱法研究长期不
同施肥对砖红壤溶解性有机质化学性质的影响[J]. 光谱学与光谱分
析, 2022, 42(10):3210-3216. ZHAO X W, WU D M, LI Q F, et al.
Response of dissolved organic matter chemical properties to long -
term different fertilization in latosol:insight from ultraviolet-visible
spectroscopy[J]. Spectroscopy and Spectral Analysis, 2022, 42(10):
3210-3216.
[16] DE HAAN H, DE BOER T. Applicability of light absorbance and
fluorescence as measures of concentration and molecular size of
dissolved organic carbon in humic Lake Tjeukemeer[J]. Water
Research, 1987, 21(6):731-734.
[17] 周江敏, 代静玉, 潘根兴. 土壤中水溶性有机质的结构特征及其与
富里酸、胡敏酸的比较[J]. 土壤, 2004, 36(1):46-50. ZHOU J M,
DAI J Y, PAN G X. Structural characteristics of dissolved organic
matter:fulvic and humic acids in soil[J]. Soils, 2004, 36(1):46-50.
[18] HELMS J R, STUBBINS A, RITCHIE J D, et al. Absorption spectral
slopes and slope ratios as indicators of molecular weight, source, and
photobleaching of chromophoric dissolved organic matter[J].
Limnology and Oceanography, 2008, 53(3):955-969.
[19] 朱大伟, 武道吉, 孙翠珍, 等. 三维荧光光谱(3D-EEM)技术在溶
解性有机质(DOM)分析中的应用[J]. 净水技术, 2015, 34(1):14-
17. ZHU D W, WU D J, SUN C Z, et al. Application of threedimensional
excitation - emission matrix(3D - EEM) fluorescence
spectrum technology in analysis of dissolved organic matter(DOM)
[J]. Water Purification Technology. 2015, 34(1):14-17.
[20] 陈颖, 何磊, 崔行宁, 等. 基于紫外光谱的水体硝酸盐氮浊度补偿
研究[J]. 光谱学与光谱分析, 2020, 40(2):472-477. CHEN Y,
HE L, CUI X N, et al. Study on turbidity compensation of nitrate
nitrogen in water based on ultraviolet spectrum[J]. Spectroscopy and
Spectral Analysis, 2020, 40(2):472-477.
[21] LIN H, BARTLETT S L, GUO L D. Distinct variations in fluorescent
DOM components along a trophic gradient in the lower Fox River-
Green Bay as characterized using one-sample PARAFAC approach
[J]. Science of the Total Environment, 2023, 902:165891.
[22] CUI H Y, WEN X, WU Z H, et al. Insight into complexation of Cd
(Ⅱ) and Cu(Ⅱ) to fulvic acid based on feature recognition of
PARAFAC combined with 2DCOS[J]. Journal of Hazardous Materials,
2022, 440:129758.
[23] SCISCENKO I, ARQUES A, MICÓ P, et al. Emerging applications of
EEM-PARAFAC for water treatment:a concise review[J]. Chemical
Engineering Journal Advances, 2022, 10:100286.
[24] 黄凯文, 马珍, 苫君月, 等. 喀斯特山区土壤溶解性有机质光谱特
征及来源解析[J]. 环境工程, 2023, 41(5):115-124. HUANG K
W, MA Z, SHAN J Y, et al. Spectral characteristics and source
analysis of soil dissolved organic matter in Karst mountainous area[J].
Environmental Engineering, 2023, 41(5):115-124.
[25] ZHANG K L, MALTAIS-LANDRY G, LIAO H L. How soil biota
regulate C cycling and soil C pools in diversified crop rotations[J].
Soil Biology and Biochemistry, 2021, 156:108219.
[26] YAN Z J, ZHOU J, LIU C Y, et al. Legume-based crop diversification
reinforces soil health and carbon storage driven by microbial biomass
and aggregates[J]. Soil and Tillage Research, 2023, 234:105848.
[27] JIAN J S, DU X, REITER M S, et al. A meta-analysis of global
cropland soil carbon changes due to cover cropping[J]. Soil Biology
and Biochemistry, 2020, 143:107735.
[28] ZHANG D B, YAO Z Y, CHEN J, et al. Improving soil aggregation,
aggregate-associated C and N, and enzyme activities by green manure
crops in the Loess Plateau of China[J]. European Journal of Soil
Science, 2019, 70(6):1267-1279.
[29] 吴炯, 施翠仙, 祖艳群, 等. 辣椒/玉米间作条件下土壤持水能力和
作物水分利用特征研究[J]. 云南农业大学学报(自然科学), 2016,
31(1):153-159. WU J, SHI C X, ZU Y Q, et al. Research on soil
moisture retention capacity and crop water use efficiency in chili
pepper and maize intercropping system[J]. Journal of Yunnan
Agricultural University(Natural Science), 2016, 31(1):153-159.
[30] KALBITZ K, KAISER K. Ecological aspects of dissolved organic
matter in soils[J]. Geoderma, 2003, 113(3/4):177-178.
[31] LI X M, SUN G X, CHEN S C, et al. Molecular chemodiversity of
dissolved organic matter in paddy soils[J]. Environmental Science amp;
Technology, 2018, 52(3):963-971.
[32] 祖艳群, 胡文友, 吴伯志, 等. 辣椒//玉米间作条件下作物对氮、磷
和钾的吸收利用特征研究[J]. 中国农学通报, 2009, 25(12):234-
239. ZU Y Q, HU W Y, WU B Z, et al. Research on uptake and
utilization characteristics of N, P and K in chili pepper//maize
intercropping system[J]. Chinese Agricultural Science Bulletin, 2009,
25(12):234-239.
[33] BLANCO-CANQUI H. Do cover crops impact labile C more than
total C? Data synthesis[J]. Soil Use and Management, 2023, 39(3):
989-1005.
[34] 周悦, 褚克坚, 苏良湖, 等. 农艺措施对土壤可溶性有机质的影响
研究进展[J]. 土壤, 2022, 54(3):437-445. ZHOU Y, CHU K J, SU
L H, et al. Effects of agronomic measures on soil dissolved organic
matter:a review[J]. Soils, 2022, 54(3):437-445.
[35] 张洪勇. 辣椒/玉米间作对根际土壤微生态的影响[D]. 泰安:山东
农业大学, 2018. ZHANG H Y. Effects of pepper / maize
intercropping on rhizosphere soil microecology[D]. Tai′ an:Shandong
Agricultural University, 2018.
[36] YANG Y, TILMAN D, FUREY G, et al. Soil carbon sequestration
accelerated by restoration of grassland biodiversity[J]. Nature
Communications, 2019, 10(1):718.
猜你喜欢
今日农业(2021年11期)2021-11-27 10:47:17
中国化肥信息(2020年3期)2021-01-07 02:12:58
今日农业(2019年12期)2019-08-15 00:56:32
中国化肥信息(2018年12期)2018-03-01 03:12:54
新农业(2017年24期)2018-01-30 08:37:26
现代农业(2015年5期)2015-02-28 18:40:52
湖南农业(2015年5期)2015-02-26 07:32:28
长江蔬菜(2014年1期)2014-03-11 15:09:55
植物营养与肥料学报(2012年1期)2012-10-26 02:50:06
中国烟草学报(2012年6期)2012-04-09 07:07:00
