
高寒草甸主要毒杂草生态策略对高原鼠兔干扰的响应
2024-12-31 00:00:00程小云孙小丹王新源尚艺婕庞晓攀李捷
草地学报 2024年8期
关键词:干扰






摘要:生态策略模型主要包括C策略(Competitive strategy),S策略(Stress-tolerant strategy)和R策略(Ruderal strategy)。本研究以高原鼠兔为例,通过野外配对法建立监测样地,分析了小型草食动物干扰对毒杂草生态策略的影响,发现高原鼠兔干扰并没有改变高寒草甸毒杂草生态策略的分布特征,但导致S策略毒杂草平均得分由79.1%变为57.9%,C策略毒杂草得分由20.9%变为30.6%,R策略毒杂草得分则增加了2.9%~52.5%,毒杂草生态策略逐渐由耐受型向竞争型过渡。高原鼠兔打破了植物群落原有的种间和种内关系,弱化区域因子决定高寒草甸毒杂草生存策略中的地位,毒杂草生态策略的决定因子由海拔转变为土壤全氮和植物地上生物量。随高原鼠兔干扰强度的增加,S策略毒杂草分布逐渐减弱,R策略毒杂草则表现为逐渐增加的变化趋势。本研究对于揭示小型草食动物干扰下高寒草甸植物对环境梯度的生态适应,并通过管理高原鼠兔而维持高寒草甸植物多样性与草地质量具有重要意义。
关键词:高寒草甸;CSR策略;毒杂草;干扰;叶片特征
中图分类号:Q94""" 文献标识码:A"""" 文章编号:1007-0435(2024)08-2459-10
Response of Ecological Strategies of Noxious Weeds to Plateau Pika
(Ochotona Curzoniae) Disturbance in an Alpine Meadow
CHENG Xiao-yun2, SUN Xiao-dan1, WANG Xin-yuan2, SHANG Yi-jie2, PANG Xiao-pan1, LI Jie1*
(1. State Key Laboratory of Herbage Improvement and Grassland Agro-ecosystems, College of Pastoral Agriculture
Science and Technology, Lanzhou University, Lanzhou, Gansu Province 730020, China; 2. Gansu Provincial Ecological Resources
Monitoring Center, Gansu Forestry Survey and Planning Institute, Lanzhou, Gansu Province 730020, China)
Abstract:The primary ecological strategies include the Competitive strategy (C),Stress–tolerant strategy (S) and Ruderal strategy (R). This study took the plateau pika as an example,establishing monitoring plots through the field pairing method to analyze the impact of small herbivore disturbance on the ecological strategies of noxious weeds. The results showed that although the disturbance by plateau pikas did not change the distribution characteristics of the ecological strategies of noxious weeds in the alpine meadows,while it led to a decrease in the average score of S-strategy noxious weeds from 79.1% to 57.9%,an increase in C-strategy noxious weeds from 20.9% to 30.6%,and an increase in R-strategy noxious weeds from 2.9% to 52.5%. Consequently,the ecological strategy of noxious weeds gradually transitioned from tolerance to competition. Plateau pikas could disrupt the original interspecific and intraspecific relationships within plant communities,weakening the role of regional factors in determining the survival strategies of noxious weeds in alpine meadows and causing the determinants of noxious weed ecological strategies to shift from altitude to total soil nitrogen and above-ground plant biomass. With the intensification of plateau pika disturbance,the distribution of S-strategy noxious weeds gradually weakened,while R-strategy noxious weeds showed a gradually increasing trend. This study is significant for revealing the ecological adaptation of alpine meadow plants to environmental gradients under small herbivore disturbance and for maintaining plant diversity and grassland quality in alpine meadows through the management of plateau pikas.
Key words:Alpine meadows;CSR strategy;Noxious weeds;Disturbance;Leaf characteristics
植物生态策略模型(C-S-R Model of Plant Ecological Strategies)反映了植物对竞争、胁迫和干扰梯度的适应性[1-2],代表植物的三种基本生存和再生策略:竞争策略(C,Competitive),应激容忍策略(S,Stress-tolerant),以及扰动反应策略(R,Ruderal)[1,3]。具有C策略的植物在资源丰富的环境中最为成功,表现为可以在生态位稳定的环境中生存;具有S策略的植物适应在资源匮乏或条件恶劣的环境中生长;而R策略型植物则可以在受扰动的地方,如人类活动、野火或洪水后的环境中繁荣[2,4-5]。研究者可以通过叶片大小和功能性状来表示植物功能性状在不同尺度上的变化(例如物种入侵和气候变化)[6],进而证明环境如何影响植物CSR策略[6-7]。因此,植物生态策略模型已成为理解草地植物如何适应环境,以及响应不同的生态压力和扰动的科学手段。
草地毒杂草构成了草地植物多样性的绝大部分,是衡量草地质量的关键因素之一[8-9]。不同毒杂草的CSR策略受非生物扰动的影响[7]。已有研究表明,草地毒杂草的器官(根、茎和叶)的功能性状会因不同海拔差异分异,例如通过减小叶面积,增加叶片厚度来减少水分蒸发[6,10]。而当温度升高时,为了应对低温和辐射,毒杂草叶片气孔面积缩小幅度相比于禾草类植物更加明显[6]。随着非生物因素变化的强大适应性,允许草地毒杂草在不同环境下保障资源吸收,强调了草地毒杂草功能组之间不同水平的表观可塑性[8]。除海拔、温度等非生物因素外,生物干扰也是改变草地植物生长和生存策略的关键途径[11-12]。研究表明,大型动物放牧长期采食或踩踏,会导致部分杂类草的功能性状和生态策略发生转变[12],且这种变化与土壤养分的增加或降低存在权衡[13-14]。除了大型草食动物放牧干扰,草地上还存在着成千上万的小型哺乳类草食动物[15]。不同于大型放牧动物人为管理模式的日趋成熟,小型草食动物种群往往呈野生分布,种群密度可以在短时间内迅速爆发[15-16],其干扰活动对草地毒杂草生存策略的影响,也会导致植物群落结构和草地质量的变化。然而,目前关于草地毒杂草CSR策略如何响应小型草食动物干扰的研究还很匮乏。
青藏高原高寒草甸生态系统作为世界上海拔最高、覆盖范围最广的高寒区域[17],是泛北极植物区生物多样性和畜牧业生产的生态基础,也是我国重要的生态安全屏障,对气候调节、碳氮固持等生态系统功能具有重要作用[14]。然而,脆弱的高寒环境和较短的植被生长周期,致使高寒草地毒杂草生长和生存策略对外界干扰的响应十分敏感[7,11]。毒杂草生态策略对外界干扰的响应结果,往往引起高寒草甸群落结构和草地质量改变。其中,高原鼠兔(Ochotona curzoniae)作为青藏高原最广泛的生物干扰因子[15,18],往往通过掘洞、排泄、刈割等多种干扰方式[19-20],增加环境异质性,影响群落物种组成,增加杂类草生物量所占的比例,进而改变高寒草甸植物群落结构[16,20]。然而,高原鼠兔干扰对高寒草甸主要毒杂草的生态策略的影响及其驱动因素仍不清楚。
高原鼠兔在高寒草地中的扩散是一个渐进的过程[20],没有高原鼠兔的开放植被栖息地,被认为是高原鼠兔的潜在分布区[21]。高原鼠兔进入这些地区的缓慢扩散过程,为评估它们对高山草地毒杂草生态策略的影响提供了一个天然实验室。因此,本文以高原鼠兔为例,采用随机配对的方法建立高原鼠兔存在样地和不存在样地,旨在揭示小型草食动物干扰对毒杂草生态系策略的影响,并探讨草地毒杂草生态策略应对小型草食动物干扰的驱动因素。本研究将为小型草食动物干扰如何影响草地毒杂草生态策略提供理论依据,也为通过高原鼠兔管理而维持青藏高原高寒草地健康供科学依据。
1 材料与方法
1.1 研究地区概况
研究地区位于青藏高原东缘,隶属甘肃省甘南藏族自治州玛曲县境内(33°06′~34°30′N,100°40′~102°29′E),属典型高原大陆性气候,多风、寒冷且潮湿,年均温、月均温差异较大。研究区年平均降水量约为615.5 mm,主要集中在5—9月,年蒸发量为1000~1500 mm,全年日照时数约为2613.9 h,年内相对无霜期仅19 d,没有绝对无霜期。该地区海拔起伏大,介于3300~4806 m,土壤类型为亚高山草甸土,土壤营养物质多以有机态形式存在,表层草皮层约有7 cm厚,有机质含量可达到100~150 g·kg–1,土壤潜在肥力较高。草地类型为高寒草甸,以嵩草属(Kobresia)的高山嵩草(Carex parvula)为主要优势种,常见伴生种有垂穗披碱草(Elymus nutans)、小花草玉梅(Anemone rivularis)、美丽风毛菊(Saussurea pulchra)、钉柱委陵菜(Potentilla saundersiana)等。
1.2 试验设计与野外调查
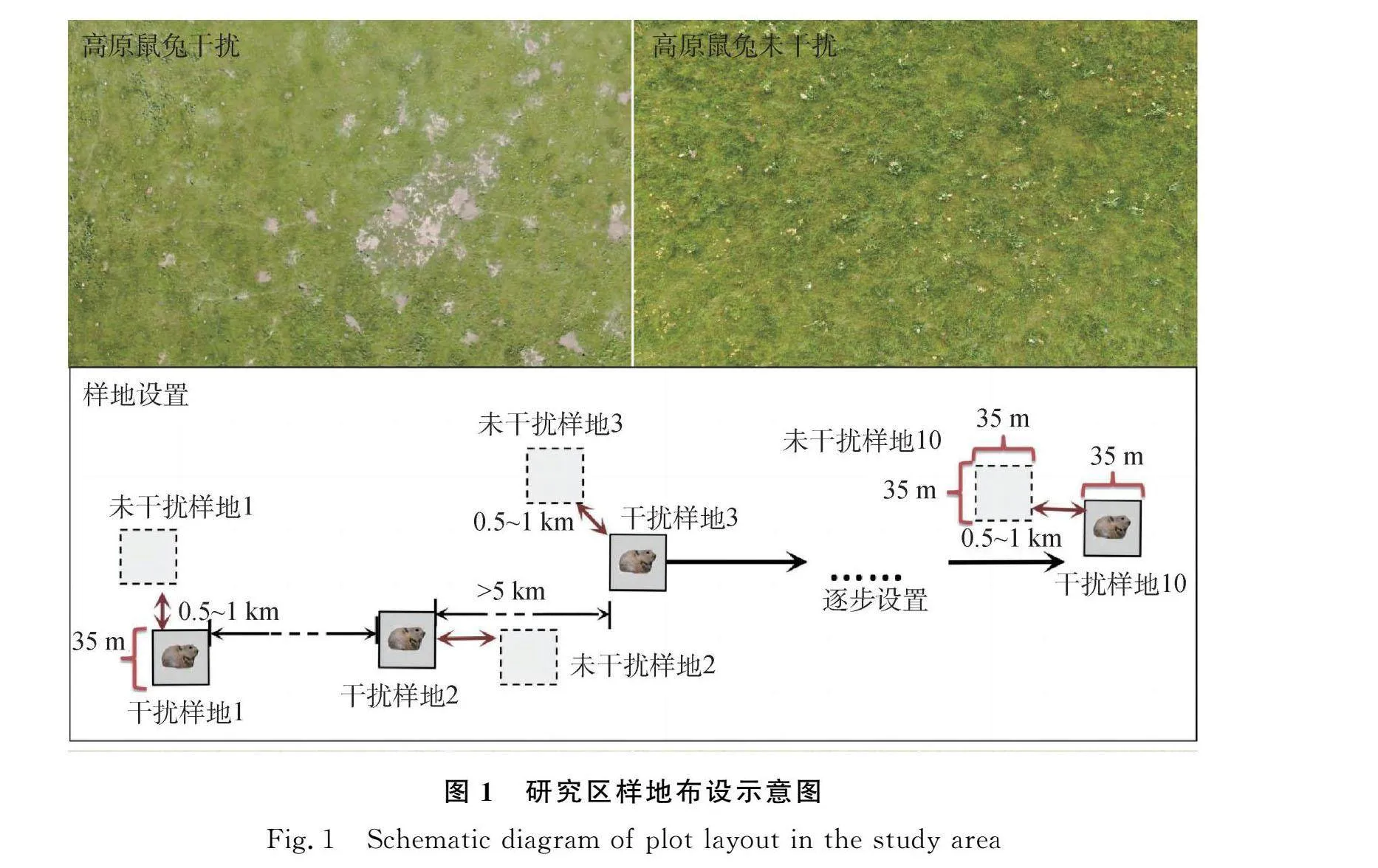
本研究于2021年采用随机配对设计并监测调查样地。首先,选择地势相对平坦、草地类型一致的高寒草甸区,确定是否有高原鼠兔干扰存在。高原鼠兔干扰以高寒草甸内目视有无高原鼠兔出没和有效洞口存在为依据[16]。由于高原鼠兔营家族生活,具有巢域特征,其平均面积大约是1262.5 m2[22],因此,本研究的样地面积大小设置为35 m×35 m(1225 m2),尽可能接近高原鼠兔的巢域面积。首先,选定第1个干扰样地,继续向任何一个方向前行,当距离超过10 km时,再次选择第2个干扰样地,以确保两个高原鼠兔家族生活范围不重复。以此类推,研究地点内累计设置12个干扰样地作为重复。当干扰样地确定后,针对每个干扰样地,配对选择一个未干扰样地。配对的未干扰样地位于干扰样地的任意方向,与干扰样地的距离大约为500 m到1000 m,距离过大则无法保证该配对干扰和未干扰样地的高寒草甸类型和地形地貌的一致性;距离过小则无法排除高原鼠兔移动对未干扰样地的影响(图1)。
研究地点内累计建立24个监测样地,包括12个高原鼠兔干扰样地和12个高原鼠兔未干扰样地。12个不同的高原鼠兔干扰样地作为重复,包含了高原鼠兔不同干扰区的生境异质性,以评估高原鼠兔干扰的影响。本研究样地均设置在冷季放牧的高寒草甸,并进行了标记和全年围封,因此从高寒草甸返青到植物枯黄期,均无大型放牧动物干扰。本研究采用“堵洞盗洞法”测定高原鼠兔有效洞口数,连续测定3 d衡量高原鼠兔干扰强度[16,23]。
高原鼠兔的繁殖期在4~5月份,其种群密度主要在7月中旬至8月底达到高峰,也是高寒草甸植物生长旺盛期,因此对高寒草甸产生的干扰效应最明显,因此本研究于2023年8月初,在甘肃省甘南藏族自治州玛曲县境内高寒草甸,沿着样带对样点进行了调查。首先,沿着草地样带收集了毒杂草植物叶片样品。本调查研究涉及的常见毒杂草包括具有毒性的草地植物以及无毒但对生态系统具有强烈侵占性,进而对原有草地生态平衡和植物群落演替造成威胁的杂类草。为了评估该地区常见毒杂草植物的生态策略,在每个样点选择常见毒杂草植物,样品采集时,选择同种不同株且大小相近的植物同一位置的叶片,并筛选出5片新鲜、健康、大小一致的叶片。在样地内,直接测量2种基本叶片功能性状的信息:叶面积和叶片鲜重。将叶片扫描后,使用ENVI 5.2计算毒杂草叶面积;用分析天平称量毒杂草植物叶片鲜重,取5片叶片的平均值作为最终的数据。测定结束后,将叶片样本装入信封袋,带回实验室测定毒杂草的叶干质量。
在每个调查样地内,通过“W”形布设植被调查样方,设立5个1平方米(1 m×1 m)的植物群落调查样方,保持样方间距约8米。如遇到大面积裸土,适当调整位置以避免其影响[24]。记录每个样方中的维管植物种类和数量,以此评估该生境的植物多样性和密度。物种鉴定完毕后,使用卷尺量取植物的高度。对于数量或丛数超过15的物种,随机选取15株作为代表进行高度测量;若少于15株,则对所有个体进行测量,并计算平均高度以代表该植物群落高度。植物群落分种盖度通过针刺法评估[21]。
随后,齐地刈割植物地上部分,装入信封带回实验室。样方调查结束后,采用环刀法测定每个调查样方的土壤容重。随后用内径为5 cm的土钻随机取0~10 cm和10~20 cm的土样3次。因为高寒草甸大多数植物根系分布于土壤表层10 cm以内,并且高原鼠兔的洞道系统一般不会深于20 cm[25]。本研究每个样地共取样15次,并按照0~10 cm和10~20 cm分层混合,共计取土样180份。
1.3 室内分析
实验室内,首先将叶片置于烘箱,105℃杀青1 h,65℃干燥48 h,由分析天平测定毒杂草的叶片干质量。其次,将样方植物地上部分在65℃烘箱干燥,并称重作为植物群落地上生物量。最后,土壤样品去除植物根系、石粒和凋落物等杂质后于室内自然风干,充分研磨过0.25 mm尼龙筛。土壤有机碳采用重铬酸钾外加热法测定,土壤全氮采用凯氏定氮法测定,土壤全磷采用钼锑抗比色法测定,测定方法参考中国土壤学会所编《土壤农业化学分析方法》[26]。
1.4 数据计算与统计
为了明确高原鼠兔干扰与未干扰下高寒草甸主要毒杂草分布特征,本研究首先在样方调查的基础上,计算了高寒草甸群落内主要毒杂草植物的重要值(Importance value,IV),计算方法采用IV=(HR+CR+DR)/3,其中IV为某一物种的重要值;HR(Relative height)为物种的相对高度;CR(Relative cover)为物种的相对盖度;DR(Relative density)为物种的相对密度。毒杂草重要值代表12个高原鼠兔干扰调查位点与12个高原鼠兔未干扰位点样地内毒杂草重要值的平均值。其次,计算高寒草甸植物群落内主要毒杂草生存策略,该过程采用电子表格计算工具“StrateFy”即可快速实现[2]。具体而言,在StrateFy表格计算工具内,分别导入叶面积、叶片鲜重以及叶干质量数据,计算出种毒杂草的叶片的肉质化指数(LSI)、叶片含水量(LWC)、叶片干物质含量(LDMC)和比叶面积(SLA)。然后,根据以上叶片功能性状数据,进一步通过表格计算工具StrateFy的内置公式,计算出每种毒杂草的C、S、R值[2]。
在本研究中使用了Origin 2018软件(OriginLab Corporation,2018)绘制毒杂草生存策略的CSR分布模式三角图,精确地根据毒杂草的C、S、R值计算结果,绘制出直观、清晰的分布模式图。CSR分布模式三角图是基于Grime的CSR理论,有效地展示了植物在竞争(C)、应激(S)和扰动(R)三个维度上的生存策略,帮助识别群落中各物种的生态位以及潜在的生存策略差异。此外,本研究利用R语言版本4.2.0,进行高原鼠兔干扰与未干扰区域内的毒杂草植物C,S,R值与主要环境因子(海拔,土壤pH,土壤容重,以及土壤氮、磷、钾含量)的相关性分析。这些环境因子被认为有可能影响植物的生存和生长状态。相关性分析采用Spearman相关性分析,由于该方法基于非参数检验,因此允许数据的非正态分布。为了直观展示毒杂草植物C、S、R值与主要环境因子之间的相关性,本研究采用了R语言中的“corrplot”包绘制相关矩阵图。“corrplot”通过颜色和图形大小展示变量间的相关系数。本研究通过配对样地t检验确定高原鼠兔干扰与未干扰区高寒草甸植物群落毒杂重要值的显著差异;采用一般线性回归模型,对各环境因子与C,S,R值进行拟合,回归分析通过R语言中的“lm”函数实现。
2 结果与分析
2.1 高原鼠兔干扰与未干扰区高寒草甸毒杂草特征
高原鼠兔未干扰区,累计出现毒杂草16种(表1)。研究区高寒草甸植物群落共出现毒杂草22种,占植被物种组成的38.8%(表2)。其中,蕨麻(Argentina anserina),高原毛茛(Ranunculus tanguticus),乳白香青(Anaphalis lactea)等重要值较高。而一些常见的毒杂草,如茵陈蒿(Artemisia capillaris),珠芽蓼(Bistorta vivipara),黄花棘豆(Oxytropis ochrocephala)等的物种重要值较低。高原鼠兔干扰下,高寒草甸植物群落毒杂草重要值相比于未干扰区显著增加(配对t检验,t=2.98,Plt;0.01)。其中,黄帚橐吾(Ligularia virgaurea)的重要值高达9.34,其次是披针叶野决明(Thermopsis lanceolata)重要值为5.22。随高原鼠兔干扰强度增加,植物群落累计物种数表现为缓慢的增加趋势,且这种变化主要来自于毒杂草物种丰富度的增加。
2.2 高原鼠兔干扰对植物生态策略分布特征的影响
高原鼠兔未干扰下,高寒草甸主要毒杂草植物主要表现出较高的S型策略,大多数毒杂草植物表现出S/CS策略,而较多毒杂草植物的R策略得分为0(图2A)。高原鼠兔干扰后,S策略毒杂草由58.7%~99.5%变为16.0%~99.8%,C策略毒杂草由0.5%~41.2%变为0.2%~60.9%,R策略毒杂草得分则增加了2.9%~52.5%。由于高原鼠兔的干扰,毒杂草的生长策略逐渐由耐受型向竞争型过渡(图2B)。
2.3 高原鼠兔干扰下植物生态策略的调节因子
高原鼠兔未干扰下,高寒草甸C策略植物和S策略植物与海拔高度显著相关(图3A)。高原鼠兔干扰下,高寒草甸S策略植物与R策略植物与植物群落地上生物量和土壤全氮含量显著相关(图3B)。
高原鼠兔未干扰下高寒草甸C策略植物和S策略植物的分布比例与海拔呈现显著线性相关关系。其中C策略植物的分布随海拔高度的增加表现为逐渐降低的变化趋势(Plt;0.01)(图4A),而S策略植物的分布表现为随海拔的增加而增加(Plt;0.05)(图4B)。高原鼠兔干扰下,S策略植物随植物群落地上生物量和土壤全氮的增加而降低(图4C,图4E),R策略植物的分布随群落地上生物量和土壤全氮的增加表现为逐渐增加的变化趋势(图5D,图5F)。
2.4 高原鼠兔不同干扰强度对植物生态策略的影响
高原鼠兔干扰下,高寒草甸毒杂草植物S策略和R策略与高原鼠兔的干扰强度呈现显著的线性相关关系。其中,高寒草甸S策略毒杂草植物随高原鼠兔干扰强度增加表现为逐渐降低的变化趋势(R2=0.44,Plt;0.05)(图5A),高寒草甸R策略毒杂草植物随高原鼠兔干扰强度增加表现为线性增加的变化趋势(R2=0.38,Plt;0.05)(图5B)。
3 讨论
本研究首次揭示了小型草食动物干扰对草地植物生态策略的影响。小型草食动物干扰对植物群落的影响不仅表现为短时间内毒杂草形态的改变[15,27],更关键的是其对植物功能性状会产生累积效应,进而导致毒杂草生态策略的转变。研究表明高原鼠兔未干扰区高寒草甸毒杂草主要表现为以S策略为优势,S/CS策略为主导的分布状态。事实上,植物CSR策略是生态位轴上的极端[1],植物可能存在于C,S,R策略之间的任何位置。即在某些环境中,毒杂草可能展现出混合策略[3,7]。高原鼠兔未干扰区的S/CS策略主导格局,主要是因为高寒草甸往往表现为低温、低养分供给以及较短的生长周期[14,28],会诱导高寒草甸毒杂草做出功能权衡,在单位叶面积上投资更多的碳[6,10],表现为资源保守性状。因此,趋向于S/CS型的生态策略的演变和进化,可以缓冲环境变化引起的代谢变化[11,13]。例如多数高寒草地植物物种具有较低的SLA和较大的LDMC[14,28],进而适应多变的生产力条件,产生广泛的储备组织(例如禾本科和莎草科植物交错发育的植被根系所形成的致密草毡层)。此外,没有高原鼠兔干扰的高寒草甸,已经形成了以黄花棘豆、蕨麻、美丽风毛菊等杂类草植物为伴生种的稳定植物群落结构。尽管杂类草植物占有较小的物种丰富度,却具有较宽的生态位。因此,面对资源抢占可以根据环境变化,将资源投入到相对快速和持续增长的部分个体中,保证了繁殖优势。因此,没有高原鼠兔干扰的高寒草甸主要表现为以S策略为优势,S/CS策略为主导的分布状态。
高原鼠兔干扰下,高寒草甸毒杂草物种组成和重要值发生转变,毒杂草生态策略逐渐向R策略过渡。总体而言,高原鼠兔干扰下竞争型植物生长受到了水分和养分压力的影响[24],致使毒杂草生长环境相对宽松,允许植被具有更强的恢复力稳定性,可以在高原鼠兔干扰后进行较快的恢复。这有利于R策略植物的发展,尤其是表型可塑性较高的毒杂草。此外,毒杂草可以在繁殖体上投入大量资源,种群可以通过繁殖体再生[9]。具体而言,首先高原鼠兔的刈割行为通过增加低矮毒杂草对光的可利用性,增加其生长潜势[19]。其次,高原鼠兔在栖息地生境的活动有利于种子传播[24,29],一些R策略杂草由于扩散能力弱,无法在特定的栖息地生存[7],高原鼠兔的存在则可以让它们更好地传播和定居。最后,高原鼠兔的掘土行为将深层的土壤重置于地表,导致植被表面和洞口附近形成裸露的土壤斑块。土壤斑块使光照资源重置[30-31],提高了土壤微生物活性和养分含量,为机会主义毒杂草种子提供适宜生长和定植的环境,增加了R策略植物的分布占比。
高原鼠兔干扰下,毒杂草生态策略的调节因素由海拔转变为土壤全氮和植物地上生物量。高原鼠兔未干扰区具有较高植被覆盖率[24],存在广泛的植物生态位重叠,因此不同海拔梯度下的温度和降雨等微气候资源的竞争更敏感,从而有利于更具竞争力的生态策略形成。高原鼠兔干扰下,随着氮和地上生物量的增加,S策略毒杂草在生态系统中的相对地位可能会下降,而更加善于在养分扰动条件下生长的R策略毒杂草可能会增加。这是由于S策略或C策略毒杂草通常在低养分环境中比较优势,因为它们对养分的利用效率很高[1,7],并且可以在养分有限的环境中存活和生长。首先,高原鼠兔干扰后有概率导致土壤氮富集,而地上生物量的增加往往伴随着土壤有机质的增加。以往的研究表明,叶片含水量和叶面积是影响植物CSR策略的主要因素,而土壤全氮含量与叶片含水量和叶面积密切相关[6]。因此土壤氮含量可以通过影响毒杂草的叶片含水量和叶面积,间接改变毒杂草的生存策略,S策略毒杂草生存优势随之降低。反之,增加的氮供应为R策略植物提供了一个更好的生长环境,从而促进它们的繁殖和扩散,在氮素充足的环境中快速生长并取得优势。其次,地上生物量的添加可能意味着生长季凋落物的积累,进而改变土壤的微生物群落结构和功能,影响土壤中的养分循环和供应,导致局部土壤或群落特征对扰动的反应更加敏感[32-33],如pH值的变化、土壤结构或土壤动物的活动变化[20],从而影响S策略毒杂草的生存。富含氮的土壤也会促进植物的快速生长[28,34-35],在养分充足的环境中,R策略植物更有能力与其他植物竞争资源,从而在生态系统中取得优势并迅速覆盖地面,影响光照,从而影响那些依赖于特定光照条件的S策略植物。第三,氮和地上生物量的增加可能会促进某些植物种类的生长,植物可能因此与S策略的毒杂草进行更激烈的竞争或者产生更强的化感作用[36-37],从而抑制S策略毒杂草的生长。最后,大部分豆科植物,可以通过与菌根共生来获取营养,从而增强其缓解环境压力的能力。例如,高原鼠兔在栖息地的排泄等行为,有机会在植被上形成肥岛效应[31],导致土壤氮的可利用性显著升高,增强黄花棘豆、披针叶野决明等固氮毒杂草植物的生长潜势。
随着高原鼠兔干扰强度的增加,R策略毒杂草所占的比例明显增加。高原鼠兔轻度或者中度干扰条件下,高寒草甸具有更强的恢复力[23],有利于植物R策略的发展,尤其是表型可塑性较高的杂草(即养分获取类植物),可以通过对养分有效性的灵活响应,将高生物量转化高原鼠兔干扰后的快速再生。已有研究表明,水和土壤养分可用性的改善将使群落的物种组成从纯S策略物种转变为具有较高C和R成分的抗逆性较低的物种[7,38]。随着高原鼠兔干扰强度的增加,挖掘行为的密集化不仅破坏了植物根系的稳定性,导致土壤营养和水分分布的不均匀[29,31]。随着高原鼠兔干扰强度的增加生境可能趋于旱化[20,32],而S策略植物通常生长缓慢,依赖于稳定的土壤环境和充足的营养供应,因此对生境的破坏特别敏感。竞争关系、生长速度和繁殖策略的变化,也可能导致S策略毒杂草所占的比例明显降低。例如S策略植物通常适应稳定环境,对资源有长期稳定的需求,高原鼠兔干扰强度的增加,逐渐减少了它们的竞争力,从而使R策略植物在资源竞争中占据优势。R策略植物通常具有更快的生长速度和更高的繁殖率,这使得它们能在经历干扰后迅速恢复和扩散。此外,高原鼠兔的取食活动等持续干扰行为,可能增强植被群落空间异质性[21,39],导致某些物种增多而使其他物种(如S策略植物)数量减少,从而间接促进了某些耐扰动的植物种类(如快速生长的R策略植物)的繁盛。然而也有研究表明,水和养分资源的稀缺会导致群落恢复困难,从而减缓了R策略物种(例如一年生植物)的生长,降低R策略植物的强度[4,11]。这暗示了在本文所研究的干扰程度下,有可能并未出现R策略降低的阈值,需要进一步扩大调查梯度,探究高原鼠兔干扰强度与CSR策略的关系。
4 结论-
天然高寒草甸毒杂草的CSR生态策略存在海拔格局,即海拔升高会显著降低C策略毒杂草的比例,而S策略毒杂草的分布随之变宽。高原鼠兔干扰会导致影响毒杂草生态策略的因素由区域尺度转变为局部群落,弱化了气候环境因子在决定高寒草甸毒杂草生存策略中的地位。随着高原鼠兔干扰强度的增加,毒杂草植物CS/S策略向R策略过渡。高原鼠兔的干扰下,R策略型毒杂草的生存更占优势。因此,基于不同的目标高寒草甸的管理存异。如果基于生态目标以保护较高的植物多样性,建议适当保护高原鼠兔栖息地生境,从而维持不同生态策略的毒杂草;基于天然草地的经济目标,应当控制高原鼠兔干扰密度在较低的水平,因为大部分毒杂草植物生态策略的转变会诱导毒杂草生物量的增加,从而降低草地可食牧草的比例。
参考文献
[1] GRIME J P. Evidence for the existence of three primary strategies in plants and its relevence to ecological and evolutionary theory[J]. American Naturalist,1977,111:1169-94
[2] PIERCE S,NEGREIROS D,CERABOLINI BEL,et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide[J]. Functional Ecology,2017,31:444-457
[3] CACCIANIGA M,LUZZARO A,PIERCE S,et al. The functional basis of a primary succession resolved by CSR classification[J]. Oikos,2006,112(1):10-20
[4] HOLMGREN M,POOTER L. Does a ruderal strategy dominate the endemic flora of the West African forests?[J]. Journal of Biogeography,2007,34:1100-1111
[5] LI Y,SHIPLEY B. An experimental test of CSR theory using a globally calibrated ordination method[J]. PLoS One,2017,12(4):e0175404
[6] 王常顺,汪诗平. 植物叶片性状对气候变化的响应研究进展[J]. 植物生态学报,2015,39(2):206-216
[7] 林马震,黄勇,李洋,等. 高寒草地植物生存策略地理分布特征及其影响因素[J]. 植物生态学报,2023,47(1):41-50
[8] 黄梅,尚占环. 青藏高原毒草型退化草地治理技术研究进展[J]. 草地学报,2019,27(5):1107-1116
[9] 张国娟,刘旻霞,李博文,等. 玛曲高寒草甸植物黄帚橐吾与莓叶委陵菜种群点格局分析[J]. 生态学杂志,2021,40(6):1660-1668
[10]王瑞丽,于贵瑞,何念鹏,等. 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例[J]. 生态学报,2016,36(8):2175-2184
[11]ZHOU T,HOU G,SUN J,et al. Degradation shifts plant communities from S-to R-strategy in an alpine meadow,Tibetan Plateau[J]. Science of The Total Environment,2021,800:149572
[12]WANG J,ZHANG C,YANG H,et al. Plant community ecological strategy assembly response to yak grazing in an alpine meadow on the eastern Tibetan Plateau[J]. Land Degradation amp; Development,2018,29:2920-2931
[13]CROSS E L,GREEN P T,MORGAN J W. A plant strategy approach to understand multidecadal change in community assembly processes in Australian grassy woodlands[J]. Journal of Ecology,2015,103(5):1300-1307
[14]LI Y Y,DONG S K,WEN L,et al. Soil carbon and nitrogen pools and their relationship to plant and soil dynamics of degraded and artificially restored grasslands of the Qinghai-Tibetan Plateau[J]. Geoderma,2014,213:178-184
[15]周雪荣,郭正刚,郭兴华. 高原鼠兔和高原鼢鼠在高寒草甸中的作用[J]. 草业科学,2010,27(5):38-44
[16]庞晓攀,贾婷婷,李倩倩,等. 高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响[J]. 生态学报,2015,35(3):873-884
[17]谢高地,鲁春霞,肖玉,等. 青藏高原高寒草地生态系统服务价值评估[J]. 山地学报,2003,21(1):1008-2786
[18]SMITH A T,FOGGIN J M. The plateau pika (Ochotona curzoniae) is a keystone species for biodiversity on the Tibetan plateau[J]. Animal Conservation,1999,2:235-240
[19]ZHANG W N,WANG Q,ZHANG J,et al. Clipping by plateau pikas and impacts to plant community[J]. Rangeland Ecology amp; Management,2020,73:368-374
[20]李捷,陈莹莹,乔福云,等. 高原鼠兔干扰对高寒草甸β多样性的影响[J]. 植物生态学报,2021,45(5):476-486
[21]LI J,QI H H,DUAN Y Y,et al. Effects of plateau pika disturbance on the spatial heterogeneity of vegetation in alpine meadows[J]. Frontiers in Plant Science,2021,12:771058
[22]FAN N,ZHOU W,WEI W,et al. Rodent pest management in the Qinghai-Tibet alpine meadow ecosystem[J]. In:Ecologically Based Rodent Management,Australian Centre International Agricultural Research,1999:285-304
[23]徐海鹏,于成,舒朝成,等. 高原鼠兔干扰对高寒草甸植物群落多样性和稳定性的影响[J]. 草业学报,2019,28(5):90-99
[24]PANG X P,GUO Z G. Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau,China[J]. Rangeland Journal,2017,39:133-144
[25]王倩,贾婷婷,庞晓攀,等. 高原鼠兔干扰对达乌里秦艽繁殖特征的影响[J]. 草业科学,2015,32(5):796-801
[26]鲁如坤. 土壤农业化学分析方法[M]. 北京:中国农业科技出版社,2000:228-264
[27]董克池,郝媛媛,董瑞,等. 高原鼢鼠鼠丘斑块空间分布格局及对植物群落特征影响[J]. 草地学报,2023,31(6):1853-1860
[28]孙建,刘国华. 青藏高原高寒草地:格局与过程[J]. 植物生态学报,2021,45:429-433
[29]周睿,花蕊,华铣泽,等. 高原鼠兔干扰下高寒草甸植物群落结构的时空动态特征[J]. 草原与草坪,2021,41(1):1-7
[30]才文代吉,谢先福,张静,等. 高原鼠兔鼠丘覆压对鼠丘上植被重建的影响研究[J]. 草地学报,2019,27(6):1615-1621
[31]邵梓桐,秦彧. 高原鼠兔干扰对高寒草地碳循环的影响研究进展[J]. 草地学报,2022,30(5):1086-1094
[32]SUN F D,CHEN W,LIU L,et al. Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan Plateau[J]. Grassland Science,2015,61(4):195-203
[33]宋文杰,张海兰,洛藏昂毛,等. 多年鼠类防控对高原鼠兔种群数量及植物群落结构的影响[J]. 草地学报,2023,31(9):2853-2859
[34]肖升木,谢学辉,薛凯,等. 氮沉积效应对植物生长及土壤微生物群落结构的影响[J]. 东华大学学报(自然科学版),2010,36(6):665-669
[35]ZHOU T C,SUN J,LIU M,et al. Coupling between plant nitrogen and phosphorus along water and heat gradients in alpine grassland[J]. Science of the Total Environment,2020,701:134660
[36]任元丁,尚占环,龙瑞军. 中国草地生态系统中的化感作用研究进展[J]. 草业科学,2014,31(5):993-1002
[37]LIAO H,LI D,ZHOU T,et al. The role of functional strategies in global plant distribution[J]. Ecography,2021,44(4):493-503
[38]姚忠,吴永明,游海林,等. 竞争-耐胁迫-杂草型植物对策理论及其应用研究进展[J]. 生态学报,2022,42(1):24-36
[39]LINDTNER P,SVITOK M,UJHZY K,et al. Disturbances by the European ground squirrel enhance diversity and spatial heterogeneity of plant communities in temperate grassland[J]. Biodiversity and Conservation,2020,29(3):853-867
(责任编辑 彭露茜)
猜你喜欢
中国新通信(2016年21期)2017-01-06 12:15:31
中国新通信(2016年21期)2017-01-06 10:31:29
中国科技纵横(2016年15期)2016-12-27 19:07:47
科技传播(2016年19期)2016-12-27 16:08:31
大众健康(2016年12期)2016-12-14 15:36:52
大众医学(2016年12期)2016-12-14 15:10:53
移动通信(2016年20期)2016-12-10 09:04:22
科学与财富(2016年26期)2016-12-01 21:05:22
科技视界(2016年14期)2016-06-08 12:55:41
