
菘蓝叶绿体基因组密码子使用偏好性分析
2024-12-31 00:00:00余志伟陈垣郭凤霞董鹏斌王红燕
草地学报 2024年8期
关键词:选择









摘要:为探究菘蓝(Isatis indigotica Fort.)叶绿体基因组密码子使用偏好性及其影响因素,本研究使用Codon W 1.4.2软件和在线软件CUSP对筛选到的52个蛋白质编码序列进行密码子使用模式分析。结果表明:菘蓝叶绿体基因组密码子总GC含量(Total GC content of codon,GCall)为37.59%,密码子第1位碱基GC含量(GC content of the first base of codon,GC1)>密码子第2位碱基GC含量(GC content of the second base of codon,GC2)>密码子第3位碱基GC含量(GC content of the third base of codon,GC3),且均小于50%,说明密码子第3位碱基拥有较低的GC含量且密码子偏好使用A或U。有效密码子数(Effective number of codon,ENC)值为35.92~55.32,密码子适应指数(Codon adaptation index,CAI)值为0.10~0.31,密码子偏好指数(Codon bias index,CBI)值为-0.24~0.20,最优密码子使用频率(Frequency of optimal codons,FOP)值为0.24~0.54,说明密码子使用偏好性较弱;通过中性绘图分析、PR2-plot分析、ENC-plot分析和对应性分析,发现菘蓝叶绿体基因组密码子使用偏好性受选择压力和突变等因素的共同影响,主要因素为自然选择。最优密码子分析共筛选出14个最优密码子,且均以A或U结尾。以上研究可为菘蓝的系统进化、环境适应性和种质改良提供理论基础。
关键词:菘蓝;叶绿体基因组;选择;密码子使用偏好性;最优密码子
中图分类号:S567.23+9""" 文献标识码:A"""" 文章编号:1007-0435(2024)08-2374-12
Codon Usage Bias Analysis of the Chloroplast Genome of Isatis Indigotica
YU Zhi-wei1,2, CHEN Yuan1*, GUO Feng-xia1, DONG Peng-bin1, WANG Hong-yan1
(1. College of Agronomy, College of Life Science and Technology, Gansu Agricultural University, State Key Laboratory of Aridland
Crop Science, Lanzhou, Gansu Province 730070, China; 2. Lanzhou Agro-technical Research and Popularization Center,
Lanzhou, Gansu Province 730030, China)
Abstract:To investigate the codon usage bias and its influencing factors in the chloroplast genome of Isatis indigotica Fort.,this study analyzed the codon usage patterns of 52 protein-coding sequences using Codon W 1.4.2 software and CUSP online software. The results revealed that the total GC content of codons in the chloroplast genome of I. indigotica was 37.59%,with GC1 gt; GC2 gt; GC3,all being less than 50%. This indicated a low GC content for the third base of codons,suggesting a preference for A or U base. Furthermore,the analysis of codon usage indices such as ENC,CAI,CBI and FOP showed ranges of 35.92 to 55.32,0.10 to 0.31,-0.24 to 0.20 and 0.24 to 0.54 respectively,pointing towards a relatively weak codon usage bias. Through various techniques such as neutral plot analysis,PR2-plot analysis,ENC-plot analysis,and correspondence analysis,we identified that selection pressure and mutation influenced the codon usage bias in I. indigotica chloroplast genome. However,natural selection emerged as the main factor affecting it. Additionally,optimal codon analysis identified a total of fourteen optimal codons with a preference for ending with A or U base. In conclusion,our findings can provide a theoretical foundation for valuable insights into phylogenetic evolution,environmental adaptability,and germplasm enhancement of I. indigotica.
Key words:Isatis indigotica;Chloroplast genome;Selection;Codon usage bias;Optimal codon
叶绿体是光合生物的细胞器,主要进行光合作用和生物合成,为其生命活动提供有机物和能量[1]。叶绿体是半自主性的细胞器[2],具有闭合双链环状结构以及独立于核基因组的叶绿体基因组;同时其具有结构稳定、序列保守和间隔区变异位点丰富以及分子进化速率较慢等特点,这些特征使叶绿体基因组广泛应用于亲缘关系、系统进化关系及遗传多样性研究[3-5]。伴随高通量测序技术的发展进步,叶绿体基因组数据库也得到了极大的丰富。其在解析植物类群系统发育关系研究中得到广泛应用,目前,已有超过一千多种植物完成了叶绿体基因组的测序,包括肉苁蓉(Cistanche deserticola Y.C.Ma)[6]、桔梗(Platycodon grandiflorus (Jacq.)A.DC.)[7]、远志(Polygala tenuifolia Willd.)[8]、单叶铁线莲(Clematis henryi Oliv)[9]等,这些测序结果在植物系统发育地位、物种鉴定和进化分析等方面发挥着重要作用[9]。密码子是DNA和蛋白质连接的重要桥梁,是生命遗传信息传递的基本单位,在蛋白质翻译过程中承担遗传信息的传递作用[10]。编码相同氨基酸的不同密码子称为同义密码子[11]。在蛋白质翻译过程中,基因倾向于使用特定的同义密码子,这被称为密码子使用偏好性(Codon usage bias,CUB)[12],密码子的选择具有物种和基因的特异性[13],这种偏好性是生物为适应环境而表现出的进化行为[14],突变和选择压力是这种偏好性存在的主要原因[15]。研究表明,密码子使用偏好性存在于自然界各种生物中,且受到诸多因素的影响[16-17]。密码子使用偏好性研究在基因结构和功能分析以及动植物、微生物和病毒的蛋白质表达等方面得到了广泛应用[17]。深入研究植物叶绿体基因组密码子的使用模式及其影响因素,能够为基因工程的载体选择和基因的表达提供理论依据,并且有助于判断未知基因的表达情况或预测某些未知功能基因,对于物种进化和遗传研究具有重要意义[18-19]。
菘蓝(Isatis indigotica Fort.)属十字花科菘蓝属,二年生草本植物,在我国广泛分布且栽培历史悠久[20]。菘蓝地下部分为板蓝根(Isatidis Radix),具有清热解毒、利咽凉血等功效[21],常用于治疗感冒、流感等病毒性疾病[22]。现代医学研究发现,板蓝根还具有抗炎、解热、提高免疫力以及预防和治疗SARS-CoV-2等作用[23-25]。其叶为大青叶(Isatidis Folium),具有清热解毒,凉血消斑的功效[21]。菘蓝作为我国传统中药历史悠久,栽培历史距今约2000余年,早在《尔雅·释草》中就有记载,《新修本草》中记载“本经所用乃是蓼蓝实也”[26]。作为传统的大宗药材,菘蓝在当前全球疫情形势下使用极为普遍,对防治流行性传染疾病有十分重要的意义。目前有关菘蓝的研究主要集中在其药用部位的有效成分、治病机理等方面。然而在菘蓝优良种质培育方面的研究则相对较少,因此如何提高其产量并稳定其质量十分重要。研究植物叶绿体基因组的密码子使用模式,可为提高基因表达载体构建效率、探讨物种进化关系、理解生物与环境适应的分子机制以及作物种质改良等方面提供数据支持。药用植物菘蓝的叶绿体基因组虽已报道[27],但关于菘蓝叶绿体基因组密码子使用偏好性的研究却非常有限。本研究对菘蓝叶绿体基因组的密码子使用偏好性进行了分析,并探究其密码子使用偏好性的影响因素,筛选出最优密码子,以期为后续进行菘蓝叶绿体基因组相关研究提供参考,为菘蓝基因功能和蛋白表达的深入研究以及菘蓝种质改良提供科学依据。
1 材料与方法
1.1 试验材料
菘蓝新鲜叶片采自甘肃省张掖市民乐县六坝镇,由甘肃农业大学陈垣教授鉴定为十字花科菘蓝属植物菘蓝,凭证标本存放于甘肃农业大学标本馆,凭证标本号为GAUML-20210625。样品经硅胶干燥处理后,取已干燥的叶片,采用CTAB法提取菘蓝的叶绿体DNA,利用1%琼脂糖凝胶电泳分析DNA的纯度和完整性,Qubit DNA试剂盒测定DNA浓度。随后使用Agilent 2100对文库的插入片段进行检测,待插入片段大小符合预期,使用Q-PCR方法对文库的有效浓度进行准确定量。质检后利用Illumina高通量测序平台Nova Seq 6000对基因组进行测序,PE读长为150 bp,最终得到2.29 Gbp的原始数据,上述过程由北京诺禾致源科技股份有限公司完成。测序得到的Clean data以菘蓝(NC_028415.1)为参考序列,在Linux系统下运用Get Organelle v 1.7.5.2软件对菘蓝叶绿体基因组进行组装,输出序列以FASTA格式存储。组装结果利用CPGAVAS2在线软件对序列进行注释,注释好的序列在Geneious软件中对存在问题的密码子位点进行手动校正。测序结果经组装、注释,上传至NCBI数据库,Genbank登录号为:0R936036.1(https://www.ncbi.nlm.nih.gov/nuccore/0R936036.1)。从菘蓝叶绿体基因组中共获取到87条蛋白编码区(Coding DNA sequence,CDS)序列,去除重复、长度小于300 bp、内部存在终止密码子或终止密码子异常的序列、保留ATG作为起始密码子的序列,得到用于分析的52条CDS序列。
1.2 试验方法
1.2.1 密码子相关参数计算 使用在线软件(CUSP(http://emboss.toulouse.inra.fr/cgi-bin/emboss/cusp)计算GC1,GC2,GC3,GCall以及密码子第3位碱基的A,T(U下同),C,G含量(A3,T3,C3,G3)。使用Codon W 1.4.2软件对每个基因的有效密码子数(ENC)、密码子适应指数(CAI)、密码子偏好指数(CBI)、最优密码子使用频率(FOP)以及相对同义密码子使用度(Relative synonymous codon usage,RSCU)进行计算。SPSS 20.0和Excel软件对结果进行统计分析。
有效密码子数(ENC)是评价同义密码子使用偏好性的指标[17],其取值为20~61,偏好性强弱与ENC值大小成反比,一般以ENC值35为评价密码子使用偏好性的强弱标准[17]。同义密码子相对使用度(RSCU)是密码子实际使用频率与无偏好性理论频率的比值[17],密码子使用偏好性强弱与RSCU值的大小成正比,RSCU值为1则代表无偏好性[28]。密码子适应指数(CAI)是指对于某一个基因,编码该蛋白的所有密码子相对于这条基因都使用最优密码子的情况下的适应系数,可预估未知基因密码子使用偏好性程度,取值为0~1,数值越大则偏好性越强[29];密码子偏好指数(CBI)表示特定基因对最优密码子子集的使用程度,取值为0~1,当最优密码子使用频次小于平均使用频次,CBI为负值[12]。最优密码子使用频率(FOP)表示基因中最优密码子占同义密码子数量的比值,取值为0~1,1表示只有最优密码子被使用,0则表示没有最优密码子被使用到[12]。
1.2.2 中性绘图分析 中性绘图常用于分析密码子使用模式和偏好性的影响因素,以GC3值为横坐标、密码子第1,2位碱基GC含量的均值(Mean of GC content of the first and second bases of codon,GC12)为纵坐标绘图,2者的相关程度表示密码子3个位置碱基的变异程度,通过对2者相关性进行分析可判断密码子偏好性的影响因素;若GC12与GC3相关性不显著,则密码子3个位置的碱基构成有明显的不同,即偏好性主要来自于选择压力[30];反之,若GC12和GC3显著相关,回归系数(对角线斜率)较大且接近1,说明影响密码子使用偏好性的主要因素为突变[17]。
1.2.3 PR2-plot分析 PR2-plot分析用于揭示核苷酸组成的影响因素,分别以G3/(G3+C3),A3/(A3+T3)为横坐标和纵坐标进行绘图,中心点表示A=T且C=G,即密码子使用无偏好性,其他各点与中心点的矢量距离则代表其偏好性强弱和方向[31]。如果基因在两条中心线上或接近的位置,这表示该基因四种碱基的使用频率大致相同,表明密码子使用偏好性只受到突变的影响;反之,四种碱基的使用频率有显著差异,密码子使用偏好性则同时受到选择压力和突变的影响[32]。
1.2.4 ENC-plot分析 分别以ENC,GC3为纵坐标和横坐标进行绘图,以公式ENC=2+GC3+29/[GC32+(1-GC3)2][33]为标准曲线,每点代表一个基因。基因与标准曲线的距离为评价指标,若基因位于标准曲线下方,且距离较远,此时的ENC值较低,表明密码子使用偏好性较强且受选择压力影响[34]。反之,若基因与标准曲线的距离越近,则密码子偏好性主要由碱基组成决定,受突变的影响较大。当选择出现在密码子第 3位碱基上时,为了分析选择压力和中性突变对密码子偏好性的影响,我们需结合ENC比值频数分布,以-0.05~0.05的范围为界对差异进行量化分析[34]。(ENC比值=(ENCact-ENCexp)/ENCact)。
1.2.5 对应性分析 基于RSCU值使用Codon W 1.4.2软件进行对应性分析[35],将各基因分布在多维向量空间中,根据基因在多维空间中的同义密码子使用情况、基因间值的变异及向量的情况,分析其主要变异来源和密码子使用偏好性的影响因素,同时将所有基因按照基因功能不同分为5类。以第1轴(Asix1)为横坐标、第2轴(Asix2)为纵坐标作图,散点代表基因,散点的分布可以用于分析密码子的使用模式,密码子使用变化的趋势以图中各基因的分散程度进行分析判断[36]。第1轴(Asix1)表示对密码子使用变化差异的最大影响,其余各轴的影响则依次降低[36]。
1.2.6 最优密码子的确定 将筛选后得到基因的ENC值从高到低的排列,选取排列在前后各10%的基因构建高、低表达数据库。综合高频密码子与高表达密码子的特点,若RSCU值>1则为高频密码子,高低表达数据库的差值(RSCU different between high and low expression groups,ΔRSCU)≥0.08则为高表达密码子;利用Codon W 1.4.2 计算RSCU值和ΔRSCU,同时满足高频密码子和高表达密码子的则为最优密码子[37]。
2 结果与分析
2.1 叶绿体基因组密码子使用偏好性分析
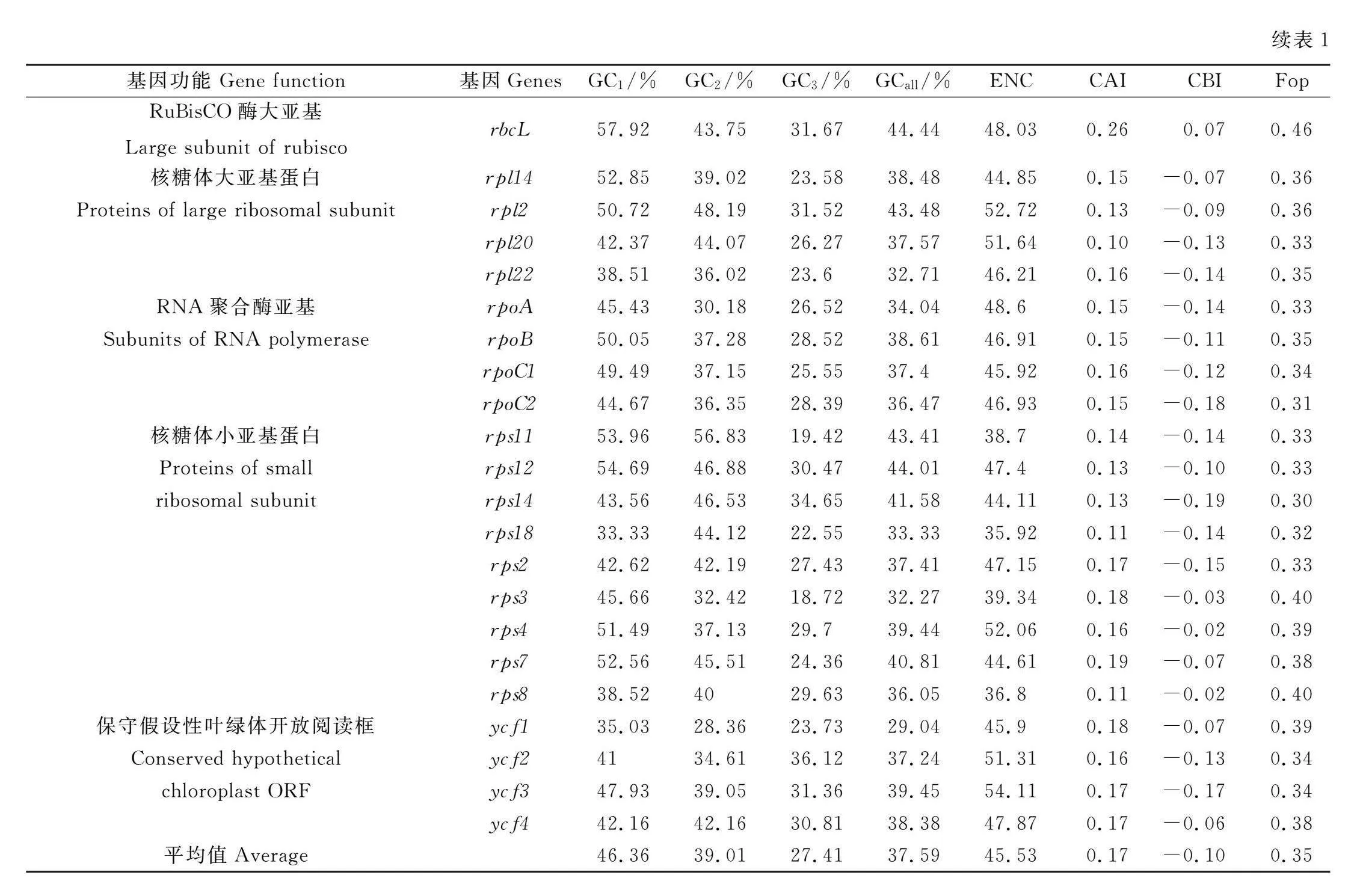
2.1.1 碱基组成及表达特征分析 如表1所示,菘蓝叶绿体基因组密码子GC1,GC2,GC3,GCall的均值分别为46.36%,39.01%,27.41%和37.59%,各基因密码子不同位置碱基的GC含量存在较大差异,GC1,GC2值普遍大于GC3值;密码子各位置碱基的GC含量均小于50%,这说明菘蓝叶绿体基因组密码子使用偏好以A或U结尾,此特性在第3位碱基上更为明显,A或U的占比更高。菘蓝叶绿体基因组ENC取值为35.92~55.32,均值为45.53,52个基因的ENC值均大于35,这表明菘蓝叶绿体基因组密码子偏好性相较弱;CAI取值为0.10~0.31,均值为0.17,则未知基因密码子使用偏好性较弱;CBI取值为-0.24~0.20,均值为-0.10,则最优密码子使用次数小于密码子平均使用次数;FOP取值为0.24~0.54,均值为0.35,则最优密码子占同义密码子数量的比值较小。以上结果均表明菘蓝叶绿体基因组密码子使用偏好性较弱。
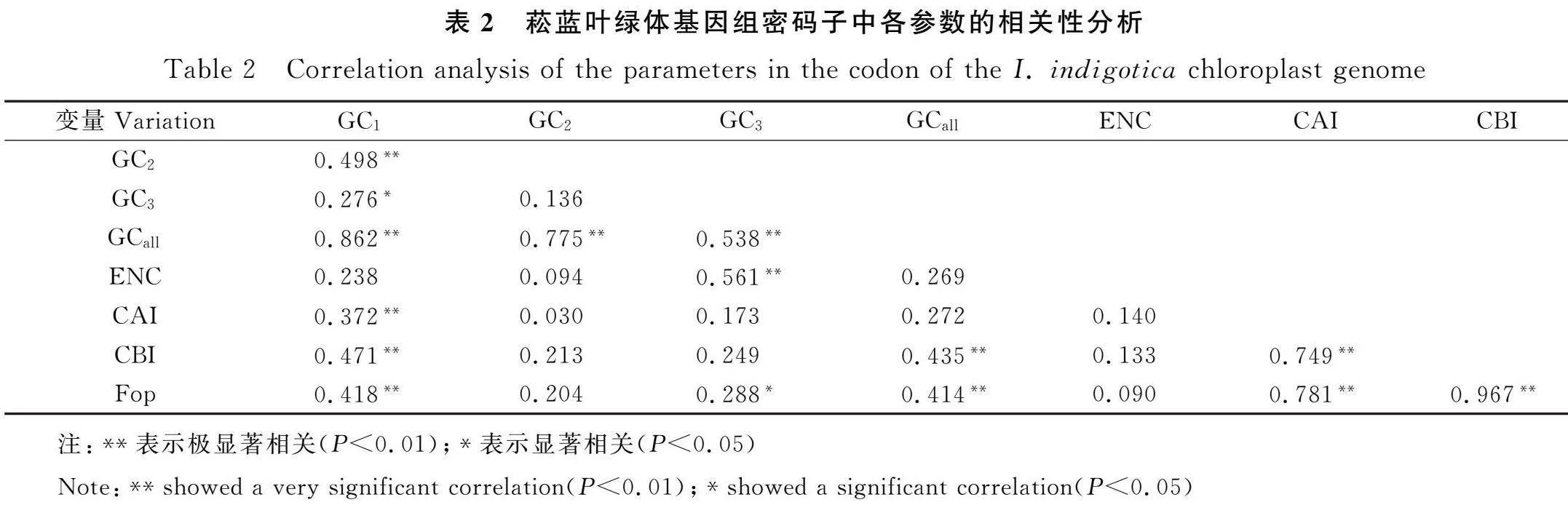
如表2所示,菘蓝叶绿体基因组密码子GC1值与GC2值极显著相关,GCall值与CC1,CG2,CG3值极显著相关,而GC3值与GC2值无显著相关,说明菘蓝叶绿体基因组密码子第1,2位碱基构成无显著差异,而第3位碱基构成差异显著。ENC值仅与GC3值极显著相关,ENC值与GC1,GC2值无相关性,说明密码子第3位碱基构成对密码子使用偏好性有显著影响;此外,GCall值与CBI和FOP值极显著相关,GC1值与CAI,CBI和FOP值极显著相关。说明密码子碱基构成对使用偏好性影响较大,特别对第3位碱基的构成影响较大。
2.1.2 相对同义密码子使用度(RSCU)分析 RSCU分析结果显示(图1):RSCU值>1的密码子有30个,表明这30个密码子的使用偏好性较强,为高频密码子,占密码子总数的46.88%。除UUG外,其余密码子均以A(13个)或U(16个)结尾,占比97%,具有明显的A或U偏向性。
2.2 中性绘图分析
如图2所示,菘蓝叶绿体基因组密码子GC12取值为31.70%~55.40%,GC3取值为18.72%~36.12%,所有基因均分布于对角线上方。atpB基因离对角线最近,其GC12与GC3比值为1.02,表明该基因密码子偏好性主要受突变影响。相关性分析结果显示,GC12与GC3无显著相关,回归系数为0.326,表明密码子前2位碱基构成与第3位碱基构成存在差异,基因组的GC含量较为保守,第3位碱基的GC含量较低。以上结果表明菘蓝叶绿体基因组密码子使用偏好性更多地受选择压力的影响,而非突变[31]。
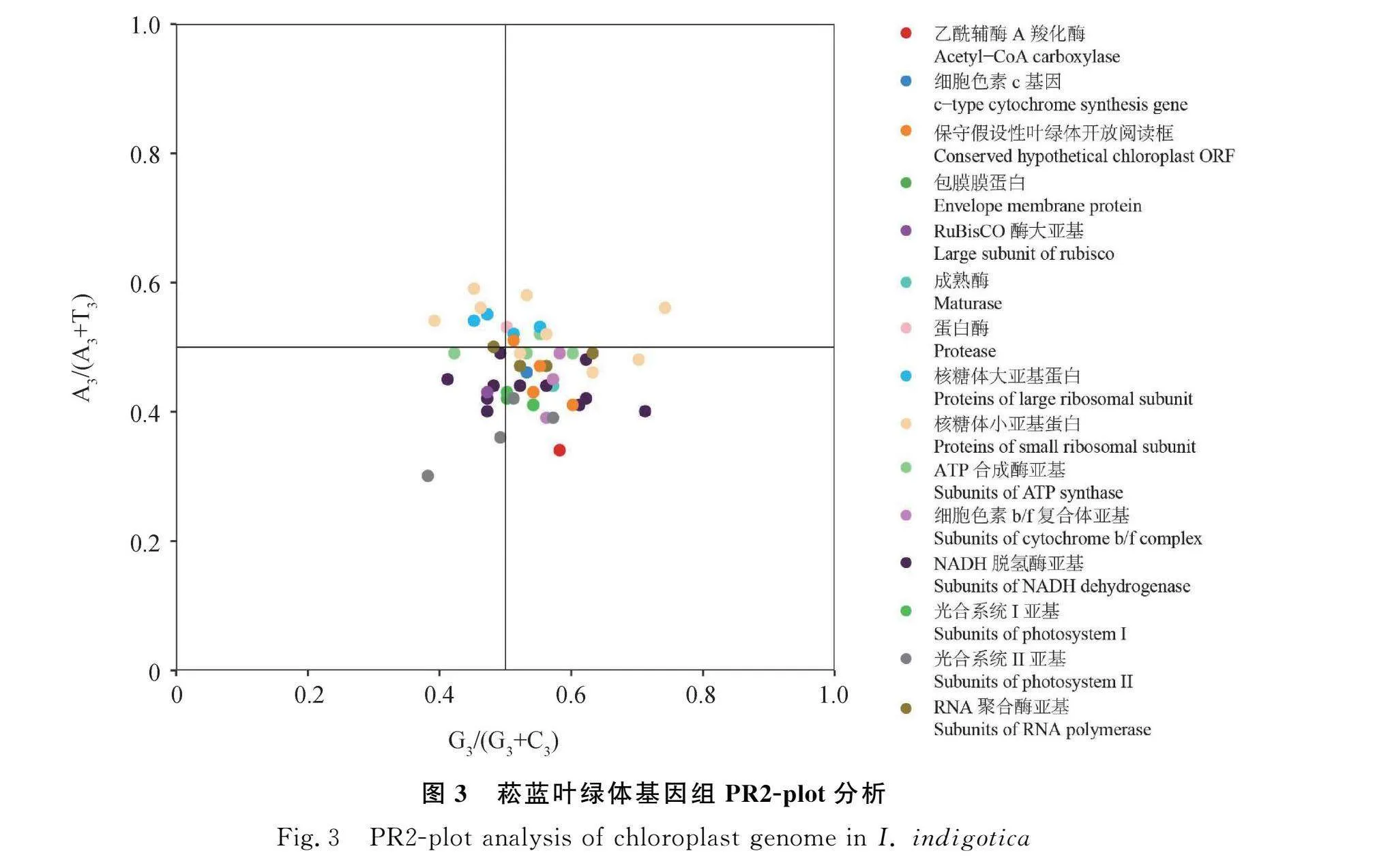
2.3 PR2-plot分析
PR2-plot分析结果显示(图3),菘蓝叶绿体基因在图中4个区域呈分散分布,仅psbA和ndhB基因距中心点最近,说明这2个基因的密码子偏好性较低,其使用偏好性主要受突变影响。菘蓝叶绿体基因组有26个基因分布在右下角区域,涉及:乙酰辅酶A羧化酶、光合系统I亚基、细胞色素c基因、成熟酶、保守假设性叶绿体开放阅读框、核糖体小亚基蛋白、ATP合成酶亚基、细胞色素b/f复合体亚基、NADH脱氢酶亚基、光合系统II亚基、RNA聚合酶亚基这些基因的部分或全部基因,说明密码子第3位碱基的使用频率T(U下同)>A、G>C,使用频率上4种碱基差异显著,因此,密码子第3位碱基的使用存在偏好性,主要受自然选择影响。反之,若基因均匀分布在两条中心线附近,则4种碱基的使用频率基本一致,密码子使用仅受突变影响。因此,充分说明菘蓝叶绿体基因组密码子使用偏好性并非仅受选择压力或突变的单一影响,而受2种因素共同影响,且受选择压力影响更大。
2.4 ENC-plot分析
如图4所示,菘蓝叶绿体基因组中大多数基因集中分布在标准曲线下方,仅3个基因(rpl20,ndhE,ycf3)其位置在标准曲线上方,表明菘蓝叶绿体基因组密码子使用偏好性受选择压力的影响较大。分析ENC比值分布(表3)发现,有14个基因位于(-0.05,0.05)区间,靠近标准曲线,表明这些基因的密码子使用偏好性受突变影响更大。有38个基因位于(0.05,0.35)区间,与标准曲线的距离较远,说明这些基因的密码子使用偏好性受选择压力影响更大。以上结果表明,相较于突变影响,菘蓝密码子使用偏好性受选择压力影响更大。
2.5 对应性分析
如图5所示,第1和第2轴的解释变化分别为11.05%和8.51%,其余各轴的解释变化均低于第1,2轴。第1轴对密码子使用偏好性的贡献率最大,表明菘蓝叶绿体基因组中密码子使用偏性存在部分变异。按照基因功能将所有基因分为5类,分别为遗传系统基因(4个)、核糖体蛋白基因(13个)、光合系统基因(26个)、其他蛋白基因(5个)和保守开放阅读框(4个)。其中,遗传系统基因集中分布在右下角,而其他各类功能基因的分布更为分散,说明这些基因密码子的使用模式差异显著,密码子使用偏好性受多种因素的影响。由表4可知,第一轴与CAI,CBI呈显著或极显著正相关,而与GC3和ENC则呈显著或极显著负相关。说明CAI,CBI,ENC和GC3对菘蓝叶绿体密码子偏好性的影响较大。以上分析说明菘蓝叶绿体的密码子使用偏好性受多种因素影响。
2.6 最优密码子
由表5可知,有19个密码子的ΔRSCU≥0.08,即高表达密码子。有30个密码子的RSCU>1,即高频密码子。满足上述两个标准的密码子为最优密码子,共14个,分别为:GCA,GCU,CGA,UGU,CAA,GGU,AUU,UUA,AAA,CCA,CCU,UCA,UAU,GUA,其中8个密码子以A结尾,6个密码子以U结尾。说明菘蓝叶绿体基因组密码子偏好以A或U结尾,这与密码子碱基构成以及相对同义密码子使用度分析的结果一致。
3 讨论
3.1 菘蓝叶绿体基因组密码子的特征
密码子使用偏好性是光合生物为应对基因组环境变化以及自然进化压力而进行的适应性选择[36],受多种因素影响[16-17]。其中,碱基构成对密码子的影响最为普遍[36],尤其密码子第3位碱基构成(GC3)。因此,GC3值被作为分析密码子使用偏好性的重要依据[38]。本研究发现,菘蓝叶绿体基因组密码子GC含量GC1>GC2>GC3,且均小于50%,表明菘蓝叶绿体基因组密码子第3位碱基拥有较低的GC含量,且密码子偏好使用A或U,这与葫芦巴(Trigonella foenum-graecum L.)[39]、珊瑚菜(Glehnia littoralis F. Schmidt ex Miq.)[40]、甜高粱(Sorghum bicolor ‘Dochna’)[41]等植物叶绿体基因组的特征相符,也符合“高等植物密码子偏好以A或T结尾”的这一假设[42]。参照Jiang等人[43]的研究,密码子使用偏好性强弱区分以ENC值35为界,菘蓝叶绿体基因组ENC的均值为45.53,有52个基因的ENC值大于35,这表明菘蓝叶绿体基因组密码子的偏好性较弱。密码子使用偏好性能够影响基因的表达水平[41],一般来说,密码子使用偏好性越强,基因表达水平越高,反之,密码子使用偏好性越弱,则基因表达水平越低[44]。本研究中,CAI,CBI和FOP的均值分别为0.17,-0.10,0.35,均表明菘蓝叶绿体基因组密码子使用偏好性较弱,因此,菘蓝叶绿体基因的表达水平相对较低。相关性分析表明,密码子碱基构成对密码子使用偏好性影响显著,尤其是第3位碱基的构成,因此,菘蓝叶绿体基因组密码子碱基构成对菘蓝叶绿体基因的表达也有很大影响。密码子使用偏好性能够调控基因的表达水平,同时最优密码子的使用能够提升翻译的效率和精确度[45]。因此,本研究涉及的含有最优密码子的基因可能呈现出较高的表达水平。
3.2 菘蓝叶绿体基因密码子偏好性的主要影响因素
根据中性绘图分析,密码子使用偏好性主要受选择压力的影响,同时也受突变影响。PR2-plot分析表明,菘蓝叶绿体密码子第3位碱基的使用频率T和G分别高于A和C,这表明选择压力在更大程度上影响了菘蓝叶绿体密码子使用偏好性。这与高守舆等[46]对白羊草(Bothriochloa ischaemum L. Keng)叶绿体基因组的研究结果相吻合。而Zhang等[47]研究小麦发现,小麦密码子第3位碱基使用频率上G显著高于C,而T和A的无显著差异。这表明叶绿体基因组的碱基构成和密码子偏好具有物种特异性。根据ENC-plot的分析结果,菘蓝叶绿体基因组密码子使用偏好性受突变的影响较小。对应性分析进一步说明,菘蓝密码子使用偏好性受到多种因素影响,其中CAI,CBI,ENC和GC3影响较大。综上所述,影响菘蓝密码子使用偏好性的主要因素是选择压力。
3.3 菘蓝叶绿体基因密码子偏好性的模式
本研究综合高频密码子和高表达密码子共得到14个最优密码子[36],且均以A或U结尾。通过前人对人参(Panax ginseng C.A.Mey.)[48]、苦荞(Fagopyrum tataricum L. Gaertn.)[37]、葫芦巴(T. foenum-graecum)[39]等植物叶绿体基因的密码子偏好性研究发现,这些植物同样表现出偏好A或U结尾的特征,说明菘蓝与其他双子叶植物一样偏好使用以A或U结尾的密码子[35],同时也说明密码子偏好性存在物种的相似性[39]。
4 结论
本研究首次对甘肃板蓝根道地产区的菘蓝叶绿体基因组密码子偏好性进行了分析,确定了14个最优密码子,密码子偏好使用A或U,且密码子使用偏好性和基因表达水平较弱;密码子碱基构成存在位置差异,密码子碱基第3位GC含量最低;密码子使用偏好性受突变和选择压力的共同作用,且主要的受选择压力的影响。以上研究可为菘蓝物种进化、基因工程研究、遗传研究、外源表达载体构建和种质改良等提供科学依据和理论支撑。
参考文献
[1] WANG X M,ZHOU T,BAI G Q,et al. Complete chloroplast genome sequence of Fagopyrum dibotrys:Genome features,comparative analysis and phylogenetic relationships[J]. Scientific Reports,2018,8(1):12379
[2] 姜汶君,郭梦月,庞晓慧. 叶绿体基因组在药用植物鉴定及系统进化研究中的应用[J]. 世界中医药,2020,15(5):702-708
[3] LI C J,LU J M,LI Y,et al. Comparative morphology of the leaf epidermis in Lobelia (Lobelioideae) from China[J]. Microscopy Research and Technique,2017,80(7):763-778
[4] 丁怡宁,毕光耀,胡赛文,等. 红花不同花色变异类型叶绿体基因组特征与系统进化分析[J]. 中草药,2023,54(1):262-271
[5] YU B,SUN Y B,LIU X F,et al. Complete chloroplast genome sequence and phylogenetic analysis of Camellia fraterna[J]. Mitochondrial DNA.Part B,Resources,2020,5(4):3840-3842
[6] 宋菊,龙月红,林丽梅,等. 五加科植物叶绿体基因组结构与进化分析[J]. 中草药,2017,48(24):5070-5075
[7] 张瑜,杜晨晖,詹海仙,等. 桔梗叶绿体比较基因组学分析及系统发育研究[J]. 中草药,2023,54(15):4981-4991
[8] 王星璐,李慧娟,赵伟,等. 远志叶绿体基因组序列特征与系统发育分析[J]. 中草药,2023,54(11):3655-3665
[9] 韦海忠,潘丽芹,田盛野,等. 单叶铁线莲叶绿体基因组的序列特征与系统发育分析[J]. 中草药,2022,53(12):3766-3773
[10]滕腾,赵懿琛,赵德刚. 鸟王茶基因密码子偏好性分析[J]. 基因组学与应用生物学,2021,40(2):795-801
[11]李秀璋,宋辉,张宗豪,等. 甘肃内生真菌基因组密码子使用的偏好性分析[J]. 草业学报,2020,29(5):67-77
[12]TANG D,WEI F,CAI Z,et al. Analysis of codon usage bias and evolution in the chloroplast genome of Mesona chinensis Benth[J]. Development Genes and Evolution,2021,231(1):1-9
[13]ZHANG R,ZHANG L,WANG W,et al. Differences in codon usage bias between photosynthesis-related genes and genetic system-related genes of chloroplast genomes in cultivated and wild solanum species[J]. International Journal of Molecular Sciences,2018,19(10):3142
[14]BOTZMAN M,MARGALIT H. Variation in global codon usage bias among prokaryotic organisms is associated with their lifestyles[J]. Genome Biology,2011,12(10):R109
[15]DURET L. Evolution of synonymous codon usage in metazoans[J]. Current Opinion in Genetics and Development,2002,12(6):640-649
[16]娄丽,袁丛军,陈锐,等. 安龙油果樟叶绿体基因组密码子偏好性分析[J/OL]. 分子植物育种:1-17[2024-02-05]
[17]原晓龙,李云琴,张劲峰,等. 降香黄檀叶绿体基因组密码子偏好性分析[J]. 广西植物,2021,41(4):622-630
[18]梁皓辉,符虹宇,黎钊坪,等. 小球藻叶绿体基因组密码子偏好性分析[J]. 分子植物育种,2020,18(17):5665-5673
[19]沈宗芳,陆添权,张志荣,等. 槲蕨属叶绿体基因组密码子偏好性分析[J]. 广西植物,2021,41(2):266-273
[20]邵玉娇,康雷,李再云. 药用植物菘蓝的遗传及远缘杂交研究进展[J]. 植物遗传资源学报,2023,24(6):1527-1533
[21]国家药典委员会. 中国药典(一部)[M]. 北京:中国医药科技出版社,2015:21-22,205-206
[22]CAO Y,XU X,ZHANG P Y. Advances in the traditional Chinese medicine-based management of viral myocarditis[J]. Cell Biochemistry and Biophysics,2015,73(1):237-243
[23]DAVID S N,GUILLERMO M R,ARDYTHE L M. Human milk glycans protect infants against enteric pathogens[J]. Annual Review of Nutrition,2005,25:37-64
[24]DENG W,XU Y,KONG Q,et al. Therapeutic efficacy of Pudilan Xiaoyan Oral Liquid (PDL) for COVID-19 in vitro and in vivo[J]. Signal Transduct Target Ther,2020,5(1):66
[25]杨春望. 板蓝根药理研究进展[J]. 中国现代药物应用,2016,10(9):282-283
[26]付照羽,冯治朋,韩颜超,等. 板蓝根研究进展[J]. 现代农业科技,2023(14):41-45
[27]YANG S,WANG Z. The complete chloroplast genome sequence of the medicinal and economic plant woad Isatis indigotica (Brassicaceae)[J]. Mitochondrial DNA.Part B,Resources,2017,2(2):514-515
[28]LIU H B,LU Y Z,LAN B L,et al. Codon usage by chloroplast gene is bias in Hemiptelea davidii[J]. Journal of Genetics,2020,99:8
[29]LIU S S,QIAO Z Q,WANG X M,et al. Analysis of codon usage patterns in“Lonicerae Flos”(Lonicera macranthoides Hand.-Mazz) based on transcriptome data[J]. Gene,2019,705:127-132
[30]MCLEAN M J,WOLFE K H,DEVINE K M. Base compositions kews,replication orientation,and gene orientation in 12 prokaryote genomes[J]. Journal of Molecular Evolution,1998,47:691-696
[31]SUEOKA N. Two aspects of DNA base composition:G+C content and translation-coupled Deviation from intra-strandruie of A=T and G=C[J]. Journal of Molecular Evolution,1999,49(1):49
[32]杨国锋,苏昆龙,赵怡然,等. 蒺藜苜蓿叶绿体密码子偏好性分析[J]. 草业学报,2015,24(12):171-179
[33]NOVEMBRE J A. Accounting for background nucleotide composition when measuring codon usage bias[J]. Molecular Biology and Evolution,2002,19(8):1390-1394
[34]NIE X J,DENG P C,FENG K W,et al. Comparative analysis of codon usage patterns in chloroplast genomes of the Asteraceae family[J]. Plant Molecular Biology Reporter,2014,32(4):828-840
[35]MCINERNEY J O. Replicational and transcriptional selecation on codon usage in Borrelia burgdorferi[J]. Proceedings of the National Academy of Sciences of the United States of America,1998,95(18):10698-10703
[36]牛元,徐琼,王嵛德,等. 大花香水月季叶绿体基因组密码子使用偏性分析[J]. 西北林学院学报,2018,33(3):123-130
[37]胡莎莎,罗洪,吴琦,等. 苦荞叶绿体基因组密码子偏爱性分析[J]. 分子植物育种,2016,14(2):309-317
[38]WAN X F,XU D,KLEINHOFS A,et al. Quantitative relationship between synonymous codon usage bias and GC composition across unicellular genomes[J]. BMC Evolutionary Biology,2004,4:19
[39]杜明川,王伟,鲍海娟,等. 葫芦巴叶绿体基因组密码子偏好性分析[J]. 草地学报,2024,32(2):409-418
[40]孙月琪,李密密,周义峰. 珊瑚菜叶绿体基因组密码子使用偏性分析[J]. 植物资源与环境学报,2023,32(6):1-10
[41]洪森荣,林顺来,李盈萍,等. 甜高粱叶绿体基因组特征及密码子偏好性分析[J]. 草地学报,2023,31(12):3636-3650
[42]CAMPBLL W H,GOWRI G. Codon usage in higher plants,green algae,and cyanobacteria[J]. Plant Physiology,1990,92(1):1-11
[43]JIANG Y,DENG F,WANG H L,et al. An extensive analysis on the global codon usage pattern of baculoviruses[J]. Archives Virology,2008,153:2273-2282
[44]ZHOU Z P,DANG Y K,ZHOU M,et al. Codon usage is an important determinant of gene expression levels largely through its effects on transcription[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(41):6117-6125
[45]SHARP P M,MATASSI G. Codon usage and genome evolution[J]. Current Opinion in Genetics and Development,1994,4:851-860
[46]高守舆,李钰莹,杨志青,等. 白羊草叶绿体基因组密码子使用偏好性分析[J]. 草业学报,2023,32(7):85-95
[47]ZHANG W J,ZHOU J,LI Z F,et al. Comparative analysis of codon usage patterns among mitochondrion,chloroplast and nuclear genes in Triticum aestivum L[J]. Journal of Integrative Plant Biology,2007,49(2):246-254
[48]石岩硕,赵永星. 人参属植物叶绿体基因组密码子偏好性分析[J]. 分子植物育种,2022,20(19):6350-6361
(责任编辑 刘婷婷)
猜你喜欢
教学月刊·小学综合(2016年11期)2016-12-05 14:19:42
商情(2016年40期)2016-11-28 10:26:18
启迪与智慧·教育版(2016年9期)2016-11-26 09:06:57
职业(2016年11期)2016-11-23 20:30:37
俪人·教师版(2016年15期)2016-11-22 04:29:04
中小企业管理与科技·下旬刊(2016年10期)2016-11-18 20:02:15
南北桥(2016年10期)2016-11-10 16:43:01
农机使用与维修(2016年10期)2016-11-10 10:01:58
中国科技博览(2016年22期)2016-11-01 14:03:21
新闻前哨(2016年10期)2016-10-31 17:41:16
