
林下生境对多花黄精光合和叶绿素荧光特征的影响
2024-12-31 00:00:00姜霞何佳丽袁丛军罗永飞丁访军
湖北农业科学 2024年12期



摘要:以林窗、林缘、林阴生境多花黄精(Polygonatum cyrtonema Hua)为研究对象,通过研究其光合和叶绿素荧光特征,揭示其在林下环境的适应机制。选择长势基本一致的3年生多花黄精,测定其光合参数净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、表观量子效率(AQY)、暗呼吸速率(Rd)、最大光合速率(Pnmax)、光补偿点(LCP)、光饱和点(LSP)和叶绿素荧光动力学参数初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、光化学淬灭系数(qP)、实际光化学量子效率(φPSⅡ)、非光化学淬灭系数(NPQ)、电子传递速率(ETR)、最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、PSⅡ有效光化学效率(Fv′/Fm′)。结果表明,不同生境多花黄精的Pn、Tr、Gs、Fo、Fm、Fv、Fv/Fm、Fv/Fo、Fv′/Fm′差异显著,以林窗最大;LSP、LCP、Pnmax均表现为林窗gt;林缘gt;林阴,林窗多花黄精的LSP、LCP、Pnmax分别为1 256、17、6.10 μmol/(m2·s);qP与φPSⅡ变化趋势一致,随着PAR的增大均呈下降趋势;NPQ随着PAR的增大呈增大趋势;ETR随着PAR的增大呈先增大后减小的趋势。多花黄精在林下生境生长均未受到抑制,但其光合和叶绿素荧光特征存在显著差异,表现为林窗更有利于多花黄精有机物质的积累。
关键词:多花黄精(Polygonatum cyrtonema Hua); 光合特征; 叶绿素荧光特征; 林下生境
中图分类号:S567.23+9" " " " "文献标识码:A
文章编号:0439-8114(2024)12-0105-05
DOI:10.14088/j.cnki.issn0439-8114.2024.12.019 开放科学(资源服务)标识码(OSID):
Effects of understory habitats on photosynthesis and chlorophyll fluorescence characteristics of Polygonatum cyrtonema Hua
JIANG Xia1,2, HE Jia-li1, YUAN Cong-jun1, LUO Yong-fei 3, DING Fang-jun1
(1.Guizhou Academy of Forestry,Guiyang" 550005,China;2.Guizhou Liping Rocky Desertification Ecosystem National Observation and Research Station,Liping" 556200,Guizhou,China;3.Chishui Changqi Town Forestry Station,Chishui" 564711,Guizhou, China)
Abstract: The photosynthetic and chlorophyll fluorescence characteristics of Polygonatum cyrtonema Hua in forest gap(FG), forest edge (FE) and forest canopy (FC) were studied to reveal its adaptation mechanism. The 3-year-old Polygonatum cyrtonema Hua with basically the same growth was selected. The photosynthetic parameters including the net photosynthetic rate (Pn), transpiration rate (Tr) and stomatal conductance(Gs), apparent quantum efficiency(AQY), dark respiration rate(Rd), maximum photosynthetic rate (Pnmax), light compensation point (LCP) and light saturation point (LSP), as well as chlorophyll fluorescence kinetic parameters including the initial fluorescence(Fo), the maximum fluorescence(Fm), the variable fluorescence(Fv), the photochemical quenching coefficient(qP), photochemical quantum efficiency(φPSⅡ), non-photochemical quenching coefficient(NPQ), electron transfer rate(ETR), the maximal photochemical efficiency(Fv/Fm), the potential activity of PSⅡ(Fv/Fo), and the effective quantum efficiency of PSⅡ(Fv′/Fm′), were measured by LI-6800. The results showed there were significant differences in Pn,Tr, Gs, Fo, Fm, Fv, Fv/Fm, Fv/Fo, Fv′/Fm′ of Polygonatum cyrtonema Hua in different habitats, their values of Polygonatum cyrtonema Hua in FG were the largest. The Pnmax, LCP and LSP of Polygonatum cyrtonema Hua in different habitats were different too, the order of values was FGgt;FEgt;FC. The LSP, LCP, Pnmax of Polygonatum cyrtonema Hua in FG were 1 256, 17, 6.10 μmol/(m2·s), respectively. The trend of qP and φPSⅡ was consistent, which decreased with the increase of PAR; NPQ increased with the increase of PAR. ETR increased first and then decreased with the increase of PAR. The growth of Polygonatum cyrtonema Hua in understory habitats light condition was not inhibited. However, there were significant differences in photosynthesis and chlorophyll fluorescence characteristics, which showed that Polygonatum cyrtonema Hua in FG was more conducive to the accumulation of organic matter.
Key words: Polygonatum cyrtonema Hua; photosynthetic characteristics; chlorophyll fluorescence characteristics; understory habitats
收稿日期:2024-02-22
基金项目:黔科合服企项目([2020]4010)
作者简介:姜 霞(1981-),女,重庆人,副研究员,硕士,主要从事森林生态、林下经济研究,(电话)18984146188(电子信箱)43644135@qq.com;通信作者,丁访军(1969-),男,研究员,博士,主要从事森林生态、林下经济研究,(电子信箱)842346405@qq.com。
多花黄精(Polygonatum cyrtonema Hua)是百合科黄精属的多年生草本植物,是中药黄精基原植物,具健脾、补气养阴、润肺、益肾等功效,其保健、药用价值高,属药食同源中药,其肥厚的根状茎富含黄精多糖、类固醇皂苷、黄酮苷等化合物[1,2]。林下经济指依托森林、林地及其生态环境,遵循可持续经营原则,以开展复合经营为主要特征的生态友好型经济,包括林下种植、林下养殖、相关产品采集加工、森林景观利用等[3]。随着贵州省林下经济的发展和多花黄精野生资源的开发利用,人工仿野生栽培受到重视。野生多花黄精常分布于林下、林缘、灌丛或山阴处,喜阴湿的环境[4],林下栽培多花黄精是林下栽培中药材的主要模式。光照是影响林下植物生长发育的主要因子,光合作用是反映植物生长对环境变化最敏感的指标之一[5]。叶绿素荧光参数是植物光合的内在探针,反映光合系统的内在特点[6]。叶绿素荧光参数在光、干旱、盐等逆境生理中广泛应用,可以比较逆境胁迫下PSⅡ的功能变化情况,包括电子供体和反应中心的变化[7,8]。桂琴等[9]研究了不同生境对大叶丹参叶表型及光合特性的影响。刘延惠等[5]研究了头花蓼光合特性及叶绿素荧光对林下光照生境变化的响应。这些研究有助于揭示植物对林下环境的适应机制。
目前对多花黄精的栽培技术[10]、化学成分[11]、生物学和经济价值[12]等方面研究较多,对多花黄精光合生理的研究相对较少,主要集中在人工控制光照对多花黄精生长、光合的影响[13],不同遮阴处理下多花黄精光响应特性[14,15],高温强光胁迫下水杨酸对多花黄精生理及光合特性的影响[16]等方面,对于自然条件下,不同生境多花黄精光合和叶绿素荧光特征鲜有报道。本研究以阔叶树林下3种生境(林窗、林缘、林阴)种植的3年生多花黄精为对象,研究其叶片光合特性和叶绿素荧光特征,揭示其在林下环境的适应机制,为多花黄精的仿野生栽培,充分利用林下资源,发展林下中药材种植及管理技术提供依据。
1 材料与方法
1.1 试验地概况
试验地处贵州省赤水市长期镇太平村,28.29°N、106.03°E,海拔791 m。该区域为半高山和高山地区,沟壑纵横,山高坡陡,地形地貌复杂;属亚热带季风气候,多年平均气温18.70 ℃,年日照百分率为28%~30%,极端最低气温-1.6 ℃,极端最高气温37.7 ℃。年平均无霜期350 d,年均降雨量1 300 mm,土壤以紫色土为主。林分类型为天然阔叶林,主要乔木有杜仲(Eucommia ulmoides Oliver)、黄皮树(Phellodendron sinii Y.C.Wu),灌木有油茶(Camellia oleifera Abel.)、穗序鹅掌柴[Schefflera delavayi (Franch.) Harms ex Diels.]。试验材料于2021年栽植,均为3年生,株行距为30 cm×30 cm。试验前选取林窗、林缘、林阴生境下生长健康、长势均匀的植物各3株。林窗、林缘、林阴光照强度分别为923.57、259.61、105.89 μmol/(m2·s)。
1.2 方法
1.2.1 光合参数的测定 选择2023年9月晴朗无云的天气,测定时间为9:00—12:00,采用Li-6800型便携式光合作用仪(美国LI-COR公司)自带红蓝光源完成各项光合生理指标测定。选取待测植株中上部完全成熟的叶片,每次测定3片样叶,每样叶3次重复。测定时设置光照强度为1 000 μmol/(m2·s),控制叶片温度为25 ℃、CO2浓度为400 μmol/mol和相对湿度为60%,测定其净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。光响应曲线的测定:设置光合有效辐射(PAR)梯度为2 000、 1 500、1 200、1 000、800、500、200、100、50、0 μmol/(m2·s),测量光合生理生态因子和电子传递速率(ETR)的变化,间隔时间3 min,获得一系列数据点。
1.2.2 叶绿素荧光的测定 采用Li-6800型自带荧光叶室测定植株的叶绿素荧光参数。用锡箔纸裹住待测叶片,待叶片暗适应12 h(21:00—9:00)后,在第2天9:00开始荧光参数的测定,首先用1束小于0.1 μmol/(m2·s)的测量光(频率为50 Hz)照射,得到初始荧光(Fo);后打开1个饱和脉冲光[8 000 μmol/(m2·s),100 Hz,1 000 ms],得到暗适应下的最大荧光(Fm),然后根据环境光强设定活化光,待F稳定后,此时的F即为稳态荧光Fs;然后再打开1个同样光照强度的饱和脉冲光,得到Fm′(光适应下最大荧光);同样打开远红光[25 μmol/(m2·s)]得到Fo′(光适应下初始荧光)。根据测定数据,可计算得出PSⅡ实际光化学量子效率(φPSⅡ)、PSⅡ潜在活性(Fv/Fo,Fv为可变荧光)、PSⅡ最大光化学量子产量(Fv/Fm)、PSⅡ有效光化学效率(Fv′/Fm′,Fv′为光适应下最大可变荧光)、非光化学淬灭系数(NPQ)和光化学淬灭系数(qP)[17]。
1.2.3 光响应曲线拟合模型 采用直角双曲线修正模型[18]对多花黄精的净光合速率进行拟合,计算出光补偿点、光饱和点、表观量子效率、最大净光合速率、暗呼吸速率([Rd])等光合参数。
直角双曲线修正模型表达式如下。
[Pn=α1-βI1+γII-Rd]" " " " " " " " " (1)
式中,I为光合有效辐射;Pn为净光合速率;α、β、γ为系数。
1.3 数据处理
采用Excel 2007、SPSS 18.0软件进行数据统计分析与制图。采用单因素方差分析法比较不同生境多花黄精叶片光合参数和叶绿素荧光参数的差异。
2 结果与分析
2.1 不同生境对多花黄精光合特征参数和荧光特征参数的影响
由表1可知,不同生境多花黄精的净光合速率、气孔导度、蒸腾速率均存在显著差异,胞间CO2浓度差异不显著。多花黄精的Pn、Tr、Gs均表现为林窗gt;林缘gt;林阴。林窗多花黄精的Pn、Tr、Gs分别比林阴显著增加了76.58%、210.87%、100.00%。
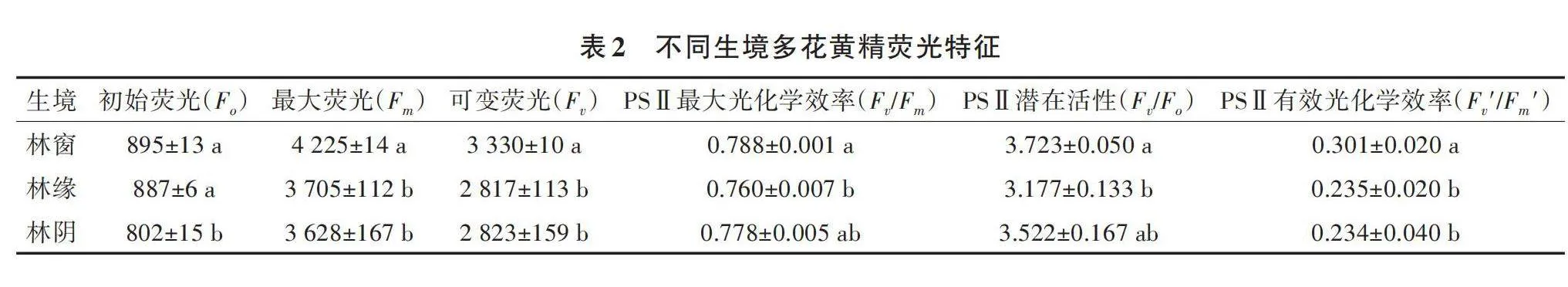
叶绿素荧光能够灵敏、快速地反映植物对光能的吸收、传递、耗散和分配等方面的状况。由表2可知,不同生境多花黄精的初始荧光、最大荧光、可变荧光、最大光化学效率、PSⅡ潜在活性、PSⅡ有效光化学效率均存在显著差异。多花黄精的Fo、Fm、Fv′/Fm′表现为林窗gt;林缘gt;林阴;Fv、Fv/Fm、Fv/Fo表现为林窗gt;林阴gt;林缘。可见,林窗多花黄精具有更大的光合作用能力和潜力。
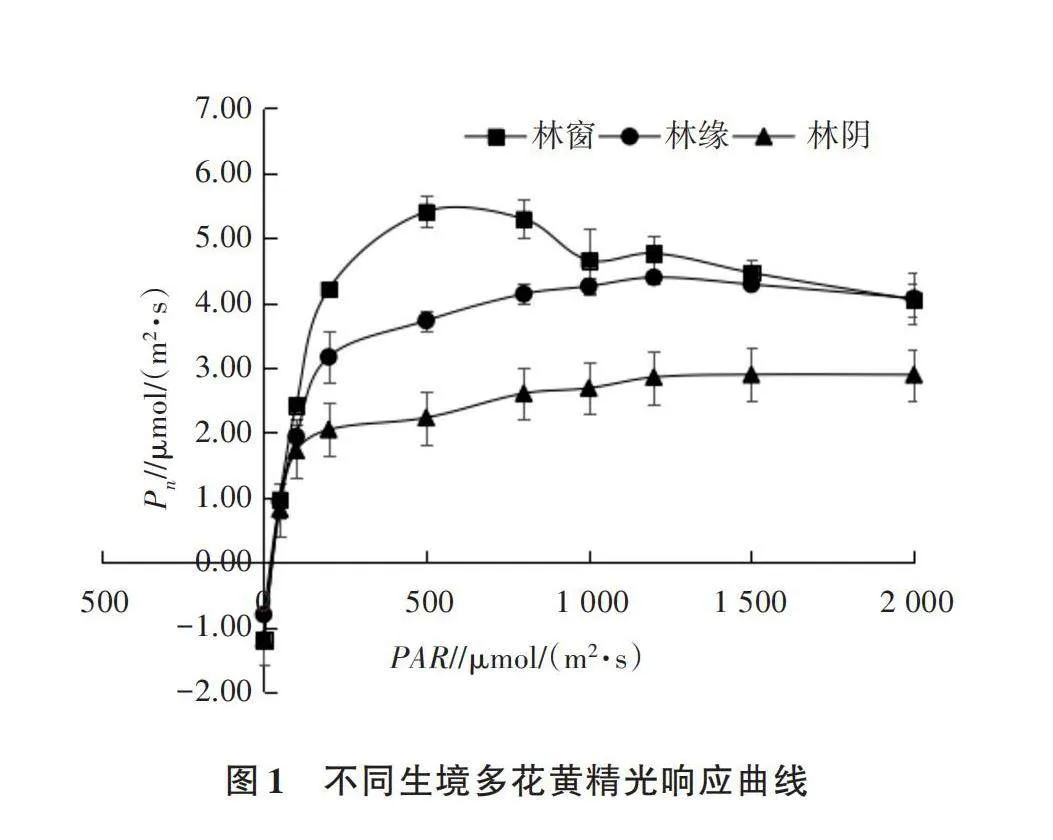
2.2 不同生境下多花黄精的光响应曲线
不同生境多花黄精的Pn对光合有效辐射变化的响应特征存在差异(图1)。光响应曲线反映了植物光合速率随光照强度增减的变化规律[19]。不同生境多花黄精的Pn随着PAR增加而上升,在达到峰值后下降。当PAR在弱光照[0~200 μmol/(m2·s)]时,Pn迅速上升,当PAR增加至500 μmol/(m2·s)时,各生境多花黄精的Pn出现分化,林窗多花黄精的Pn快速增加,超过林缘和林阴。
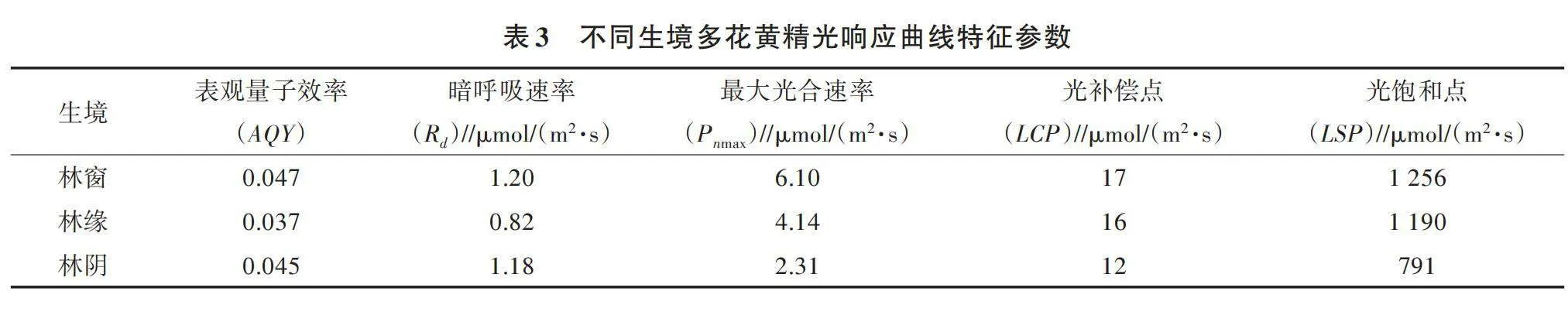
光饱和点(LSP)和最大光合速率(Pnmax)反映植物对高光强的利用能力,光补偿点(LCP)、暗呼吸速率、表观量子效率(AQY)反映植物的耐阴能力[20]。由表3可知,多花黄精的LSP、LCP、Pnmax均表现为林窗gt;林缘gt;林阴,林窗多花黄精的LSP、LCP、Pnmax分别为1 256、17、6.10 μmol/(m2·s),分别比林阴增加了58.79%、41.67%、164.07%。AQY、Rd均表现为林窗gt;林阴gt;林缘,林窗多花黄精AQY为0.047、Rd为1.20 μmol/(m2·s),分别比林缘增加了27.03%、46.34%。由此可见,生长在3种生境的多花黄精对光强的适应性不同,生长在林窗的多花黄精叶片光饱和点较高,光合能力高于林缘和林阴的叶片。
2.3 不同生境下多花黄精荧光特性对PAR变化响应
由图2可知,不同生境多花黄精的光化学淬灭系数和实际光化学量子效率差异显著(Plt;0.05)。随着PAR的增加,林窗、林缘、林阴多花黄精的qP和φPSⅡ均呈下降趋势,反映出PAR增加使PSⅡ反应中心的光能利用效率和开放程度逐渐减小。其中,随着光照强度的增加,林阴多花黄精的qP和φPSⅡ下降幅度大于林窗。不同生境多花黄精的非光化学淬灭系数和电子传递速率差异显著(Plt;0.05)。当PAR在0~800 μmol/(m2·s)时,不同生境多花黄精的NPQ随着PAR的增大而增大,当PAR超过800 μmol/(m2·s)时,NPQ基本趋于平缓;当PAR在0~500 μmol/(m2·s)时,不同生境多花黄精的ETR随着PAR的增大而增大,当PAR超过500 μmol/(m2·s)时,ETR呈下降趋势。
3 小结与讨论
光合作用是植物生长快慢的重要影响因素,主要受气孔因素和非气孔因素的限制,叶肉多孔性和叶表气孔是植物进行有效气体交换的前提条件[21]。而不同光照条件会影响植物叶片的气孔密度,进一步影响植物的净光合速率、蒸腾速率和气孔导度[22]。本研究中,林窗和林缘多花黄精的气孔导度显著高于林阴,且净光合速率、蒸腾速率比林阴大,这可能是由于气孔的变化改变了叶片气孔的特性,导致林窗多花黄精气孔导度变大,对CO2的固定能力增加,净光合速率和蒸腾速率显著提高,通过光合作用合成有机物的能力提高。
光饱和点和光补偿点是植物重要的光合生理指标,反映了植物对光照的适应和利用能力,是判断植物耐阴性的重要指标[23]。具有较高光饱和点、较低光补偿点的植物,适应光环境的能力较强[24]。有研究表明,光补偿点小于20 μmol/(m2·s)的植物为阴生植物[25]。本试验中,3种生境多花黄精光补偿点小于20 μmol/(m2·s),说明多花黄精是阴生植物,与李迎春等[13]关于多花黄精的研究结果一致。本试验中,多花黄精光饱和点、光补偿点、最大光合速率均表现为林窗gt;林缘gt;林阴,这可能是经过一段时间的适应,多花黄精能够适应弱光环境。这与王树凤等[26]对弗吉尼亚栎在遮阴条件下的光合变化研究结果一致。光合参数指标的分析表明,多花黄精利用强光和弱光的能力较强,对光环境的适应范围较宽,具有一定的耐阴性。
叶绿素荧光动力学参数是有效衡量植物的受害程度和光合潜能高低的指标[27]。本试验中,不同生境多花黄精的初始荧光、最大荧光、可变荧光、PSⅡ最大光化学量子产量、PSⅡ潜在活性、PSⅡ有效光化学效率差异显著,以林窗多花黄精的值最大,表明多花黄精在3种生境光系统PSⅡ反应中心的状态差异显著,从而导致多花黄精光合速率差异显著。有研究表明,光化学淬灭系数和电子传递速率等参数被认为是植物叶片光合作用效率的重要依据[23]。林窗的PSⅡ实际光化学量子产量、光化学淬灭系数和电子传递速率都高于林缘和林阴,说明林窗多花黄精的PSⅡ反应中心色素分子较多,能较快地将光能转化为电能,提高光合能力,从而使得光合速率较高。PSⅡ最大光化学量子产量是反映植物的潜在最大光合能力和光化学转换效率的良好指标[28],植物在不受胁迫条件下PSⅡ最大光化学量子产量的波动范围为0.75~0.85[29],本试验PSⅡ最大光化学量子产量的波动范围为0.760~0.788,表明3种生境多花黄精的PSⅡ系统光能转换效率正常,没有受到光抑制。植物光合器官的结构和功能受外因和内因的综合影响,对不同光照强度可能表现一定程度的适应[30]。进一步说明,多花黄精在3种生境下光合器官没有受到损伤,反映其对林下环境的适应,这与其野外自然分布环境条件一致。
综上所述,多花黄精具有耐阴性,在阔叶林下能很好地生长并适应林下生境。林窗多花黄精叶片净光合速率、光饱和点、PSⅡ实际光化学量子产量、电子传递速率较高,对光能利用率高,有利于其地下根茎生长。因此,在实际生产中,可以通过间伐、修剪等措施,改善林下光合有效辐射,营造有利于多花黄精生长的生境,为仿野生栽培多花黄精提供技术支撑。
参考文献:
[1] 倪天宇,罗晓朦,张春椿,等.不同生境下多花黄精化学成分比较[J].中成药,2020,42(11):2948-2953.
[2] 邱泽澜,陈 锦,张 卓,等.湖南省多花黄精炭疽病病原鉴定及其药剂筛选[J].西南农业学报,2023,36(1): 91-97.
[3] 王 妍,陈幸良.我国林下经济研究进展[J].南京林业大学学报(人文社会科学版),2022,22(4):80-87.
[4] 刘跃钧,曾岳明,叶征莺,等.多花黄精栽培技术研究进展[J].中国现代中药,2022,24(4):715-720.
[5] 刘延惠,姜 霞,丁访军,等.头花蓼光合特性及叶绿素荧光对林下光照生境变化的响应[J].水土保持学报, 2017,31(6):345-352.
[6] 周 婷,张往祥,范俊俊,等.不同栀子光合和荧光特性比较[J].林业科技开发,2015,29(5):18-23.
[7] 黄平升,刘世男,李 婷,等.外源硅对盐胁迫下黄果厚壳桂幼苗的光合荧光及抗氧化特性的影响[J].中国农学通报,2022," " " 38(23):32-38.
[8] KOJI Y,MICHIO K,MITSUTAKA T,et al. Correlation between chloroplast ultrastructure and chlorophyll fluorescence characteristics in the leaves of rice (Oryza sativa L.) grown under salinity[J].Plant production science,2008,11(1):139-145.
[9] 桂 琴,郭其强,袁刚毅,等.林下生境对大叶丹参叶表型及光合特性的影响[J].森林与环境学报,2023,43(2): 185-193.
[10] 龚雄夫,邹 辉,肖剑锋.不同光照强度对多花黄精生长的影响研究[J].中国果菜,2019,39(9):35-37.
[11] 李亚霖,周 芳,曾 婷,等.药用黄精化学成分与活性研究进展[J].中医药导报,2019,25(5):86-89.
[12] 吴媛媛,徐庆国.多花黄精的生物学和经济价值研究进展[J].安徽农业科学,2017,45(34):128-130.
[13] 李迎春,杨清平,陈双林,等.光照对多花黄精生长、光合和叶绿素荧光参数特征的影响[J].植物研究,2014,34(6):776-781,786.
[14] 童 龙,张 磊,刘小明,等.遮阴对多花黄精光合特性的影响[J].东北大学学报,2020,48(9):76-79.
[15] 梁永富,易家宁,王康才,等.遮阴对多花黄精生长及光合特性的影响[J].中国中药杂志,2019,44(1):59-67.
[16] 梁永富,王康才,薛 启,等.高温强光胁迫下水杨酸对多花黄精生理及光合特性的影响[J].南京农业大学学报,2018,41(5):839-847.
[17] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,19(4):444-448.
[18] 叶子飘,于 强.一个光合作用光响应新模型与传统模型的比较[J].沈阳农业大学学报,2007,38(6): 771-775.
[19] 姜 霞,苏春花,伍生磊,等.白云质砂石山地区翅荚香槐春季光合生理特性研究[J].广东农业科学,2017, 44(9):41-47.
[20] 刘佳哲,余小红,樊东函,等.广西4种珍贵树种在桉树林下间种的光合响应[J].中南林业科技大学学报, 2023,43(1):96-103.
[21] 张青青,周再知,王西洋,等.施肥对柚木光合生理和叶绿素荧光特性的影响[J].中南林业科技大学学报,2021,41(4):31-38.
[22] 李 威,杨德光,牟 尧,等.去遮荫后东北红豆杉幼苗和幼树光合特性对比[J].林业科学,2018,54(2): 179-185.
[23] 高贵宾,钟 浩,吴志庄,等.不同混生地被竹光合和荧光特征比较[J].福建农林大学学报(自然科学版), 2016,45(5):515-521.
[24] 李 威,赵雨森,周志强,等.不同光环境下东北红豆杉幼树叶光合和荧光特性比较[J].经济林研究,2012, 30(1): 51-55,94.
[25] 蒋高明.植物生理生态学[M].北京:高等教育出版社,2004.
[26] 王树凤,施 翔,陈益泰,等.遮荫对弗吉尼亚栎苗木生长及光合-荧光参数的影响[J].林业科学研究,2019, 32(5):99-106.
[27] BAGHERI V,SHAMSHIRI M H,ALAEI H,et al. The role of inoculum identity for growth, photosynthesis, and chlorophyll fluorescence of zinnia plants by arbuscular mycorrhizal fungi under varying water regimes[J]. Photosynthetica,2019,57(2):409-419.
[28] 凡莉莉,MUHAMMAD W K T,张洋洋,等.不同生物炭处理对福建柏光合荧光特性的影响[J].中南林业科技大学学报,2021,41(3):37-44.
[29] 朱志勇,陈晓东,张 韫,等.花龟竹与龟甲竹的光合荧光特性比较[J].甘肃农业大学学报,2019,54(4): 142-149,158.
[30] 赵 洋,陈 云,张晓晨,等.遮阴对少叶花楸幼苗光合和叶绿素荧光参数的影响[J].东北林业大学学报, 2019,47(1):30-34.
