
蓝紫色月季品种“紫霞仙子”花发育阶段转录组分析
2024-12-31 00:00:00孙骏飞孟宪水张晶莹任凤山李益
山东农业科学 2024年11期


关键词:“紫霞仙子”月季;蓝紫色花色;转录组分析;类黄酮;花青素;类胡萝卜素
中图分类号:S685.12 文献标识号:A 文章编号:1001-4942(2024)11-0013-08
月季(Rosa hybrid)品种多、花色丰富且适应性强,是世界切花最多的花卉种类。花色作为月季最重要也是最受人们关注的经济性状之一,主要受花瓣中的黄酮类和类胡萝卜素类色素的种类及含量影响。黄酮类色素多数属于水溶性色素,存在于液泡中,花朵的红、粉、绿等主要受其影响;类胡萝卜素类色素多数是脂溶性的,存在于质体中,对花朵的橙色至黄色影响比较大。花色的形成受多种基因的影响及控制,其中,转录因子复合体R2R3MYB、WD40和bHLH等花青素合成相关转录因子以及MADS-box、NAC和AP2/ERF等类胡萝卜素合成相关转录因子,通过调控查尔酮合成酶基因(CHS)、查尔酮异构酶基因(CHI)、花青素3-糖基转移酶基因(A3GT)、二氢黄酮醇还原酶基因(DFR)、类黄酮3-羟化酶基因(F3H)、类黄酮3′,5′-羟化酶基因(F3′5′H)、八氢番茄红素合成酶基因(PSY)、八氢番茄红素去饱和酶基因(PDS)、ζ-胡萝卜素去饱和酶基因(ZDS)、紫黄质脱环氧化酶基因(VDE)、玉米黄质环氧化酶基因(ZEP) 等结构基因的表达水平来调控花色。此外,色素合成相关结构基因还受到IncRNA和siRNA等转录后调控的影响。因此,花色调控是通过一个非常复杂的调控网络进行的。
蓝色是花卉中比较少的花色,蓝色花仅占花卉总量很小的比例,象征高贵,颇受人们青睐。蔷薇属缺少蓝色色素合成的关键结构基因F3′5′H,因此自然界缺少蓝色玫瑰与月季品种。为满足市场需求,近年来研究人员培育出“蓝色海洋”、“紫霞仙子”等接近蓝紫色的月季品种,具有较高观赏价值和经济价值,也具有重要的研究价值。本研究选用蓝紫色月季品种“紫霞仙子”(Rosa‘Nightingale’),在其花朵不同发育期采集花瓣样品,进行转录组测序及差异表达基因筛选和富集分析,并挖掘类胡萝卜素和类黄酮合成相关基因,筛选出引起其花色趋向蓝色的关键基因,为月季种质创新提供重要参考。
1材料与方法
1.1供试材料与设备
1.1.1植物材料 于5月中旬在山东省农业科学院济阳试验示范基地挑选生长状况良好、没有病虫害的三年生“紫霞仙子”月季植株。根据花的不同发育阶段,分别在花蕾期(N_S1)、初开期(N_S2)、半开期(N_S3)、盛开期(N_S4)和落花期(N_S5)5个时期采集花朵的第3~5轮花瓣,每个样本中至少包含取自10朵不同花的花瓣,每个时期3个重复,共得到15个转录组测序样本。样品采用液氮冷冻,送北京诺禾致源科技股份有限公司进行转录组测序。
1.1.2主要设备 冷冻高速离心机,-80℃超低温冰箱,核酸定量仪,全自动凝胶图像分析系统,水平电泳仪(美国Bio-Rad公司),PCR扩增仪(伯乐T100),Eppendorf移液器(德国Eppendorf公司),全自动毛细管电泳系统(Agilent 2100 bio-analyzer),分光光度计NanoDrop ND-1000(美国NanoDrop公司)。
1.2实验方法
1.2.1转录组测序 采用天根多糖多酚总RNA提取试剂盒从“紫霞仙子”月季花瓣样品中提取总RNA,并使用Agilent 2100 bioanalyzer严格检测RNA样品的完整性,以RNA总量gt;0.4μg、RIN5.6为合格标准。质检合格后,把不同文库按照有效浓度及目标下机数据量需求pooling后进行Illu-mina测序。
1.2.2差异基因筛选及富集分析 差异分析分为3个主要阶段:首先,对原始的readcount进行标准化,并对测序深度进行校正。其次,应用统计学模型计算假设检验中的概率值(P-value);进行多重假设检验校正后,计算得到FDR值(FDR指的是错误发现率,Padj是它的常见形式)。富集分析基于超几何分布原理,其中差异基因集为差异显著分析所得并注释到GO(Gene Ontology)或KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库的基因集,而将所有参与差异显著分析并注释到GO或KEGG数据库的差异基因集作为背景基因集。
1.2.3花色合成相关基因的挖掘 结合KEGG注释和Blast方法,对“紫霞仙子”月季类黄酮、花青素和类胡萝卜素合成通路上的功能基因进行识别。从Phytozome网站(https://phytozome. jgi.doe.gov/pz/portal.html)下载草莓、苹果中已经确定与类黄酮、花青素和类胡萝卜素合成相关的蛋白序列,用其比对月季全基因组,通过NCBI蛋白数据库中的保守序列鉴定筛选月季基因。
2结果与分析
2.1转录组分析
2.1.1转录组数据质量评估 对“紫霞仙子”月季花发育5个时期的15个样品进行Illumina测序,结果(表1)显示,去除低质量reads后,共获得1634066710条、99.68Gb的有效数据;每个样品的clean bases都在5.96Gb及以上,GC含量为45.55%~46.23%;Q30碱基所占百分比在92.75%及以上。以上数据证明本次转录组测序质量良好,结果可信。
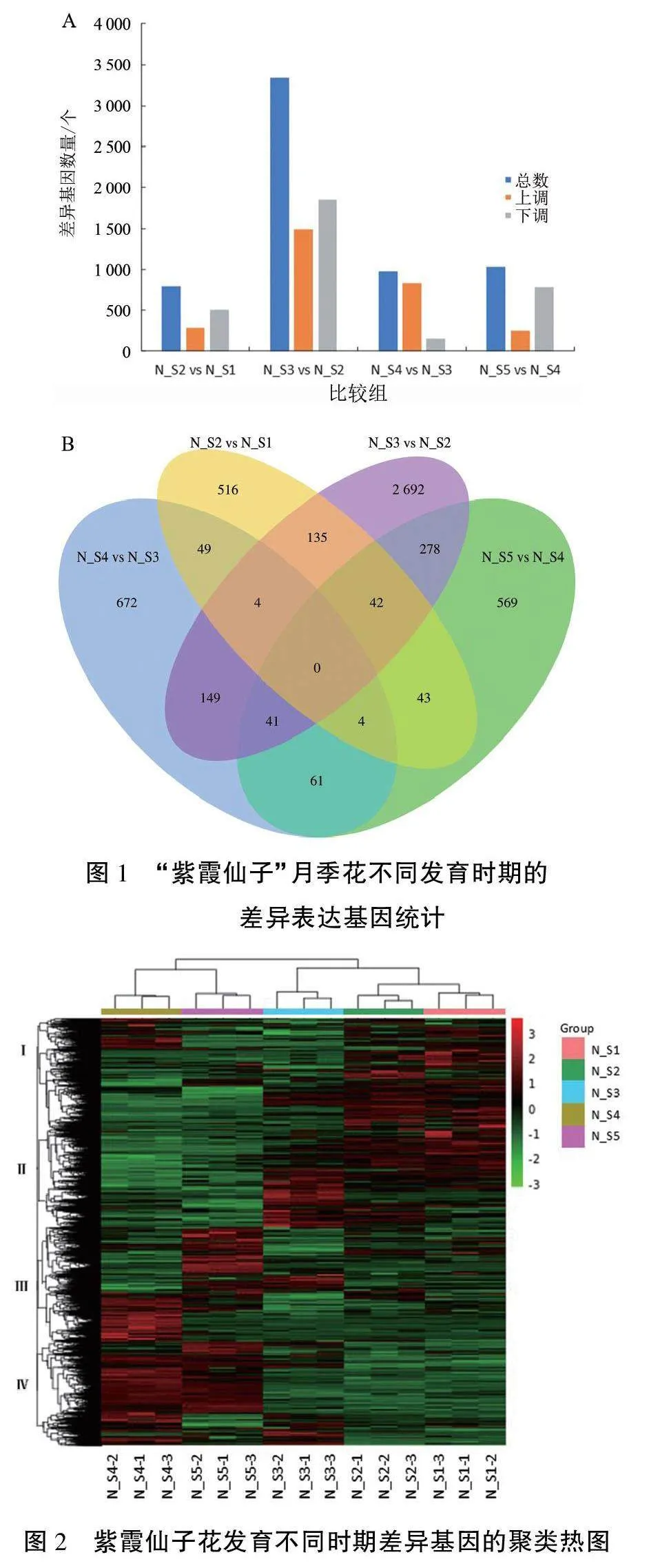
2.1.2不同发育时期差异表达基因分析 利用转录组测序数据对“紫霞仙子”月季花不同发育时期进行比较,在4个比较组中,N_S2 vs N_S1共找到793个差异基因,其中上调286个,下调507个;N_S3 vs N_S2差异基因最多,共找到3341个,其中上调1492个,下调1849个;N_S4 vs N_S3共找到980个差异基因,其中上调830个,下调150个;N_S5 vs N_S4共找到1038个差异基因,其中上调255个,下调783个(图1A)。
用韦恩图展示4组差异基因的统计结果,可以看出在N_S2 vs N_S1、N_S3 vs N_S2、N_S4 vs N_S3、N_S5 vs N_S4组合中分别有516、2692、672、569个特异的差异基因(图1B)。
聚类热图可以反映“紫霞仙子”月季各发育时期的差异基因表达情况。由图2可见,花发育不同时期的基因表达存在明显差异,但每个时期的生物学重复之间不存在显著差异。“紫霞仙子”月季花基因的表达可分为4簇,Ⅰ簇在N_Sl和N_S2时期表达量较高;Ⅱ簇在N_S3时期表达量最高,N_S4与N_S5时期表达量较低;Ⅲ簇在不同时期均有表达,其中N_S4与N_S5时期表达量最高;Ⅳ簇在N_S4与N_S5时期表达量较高,在N_S3时期少量表达。整体而言,Ⅰ簇在花发育初期表达量较高,随着花发育进入半开期则维持在比较低的水平;Ⅱ簇在花发育的前中期表达量较高,而在花的发育进入盛开期之后表达量急剧降低,在盛开期与落花期表达量极低;Ⅲ簇和Ⅳ簇主要表达于花发育后期,在花发育的前中期仅有少量表达。
2.2差异基因的GO富集分析
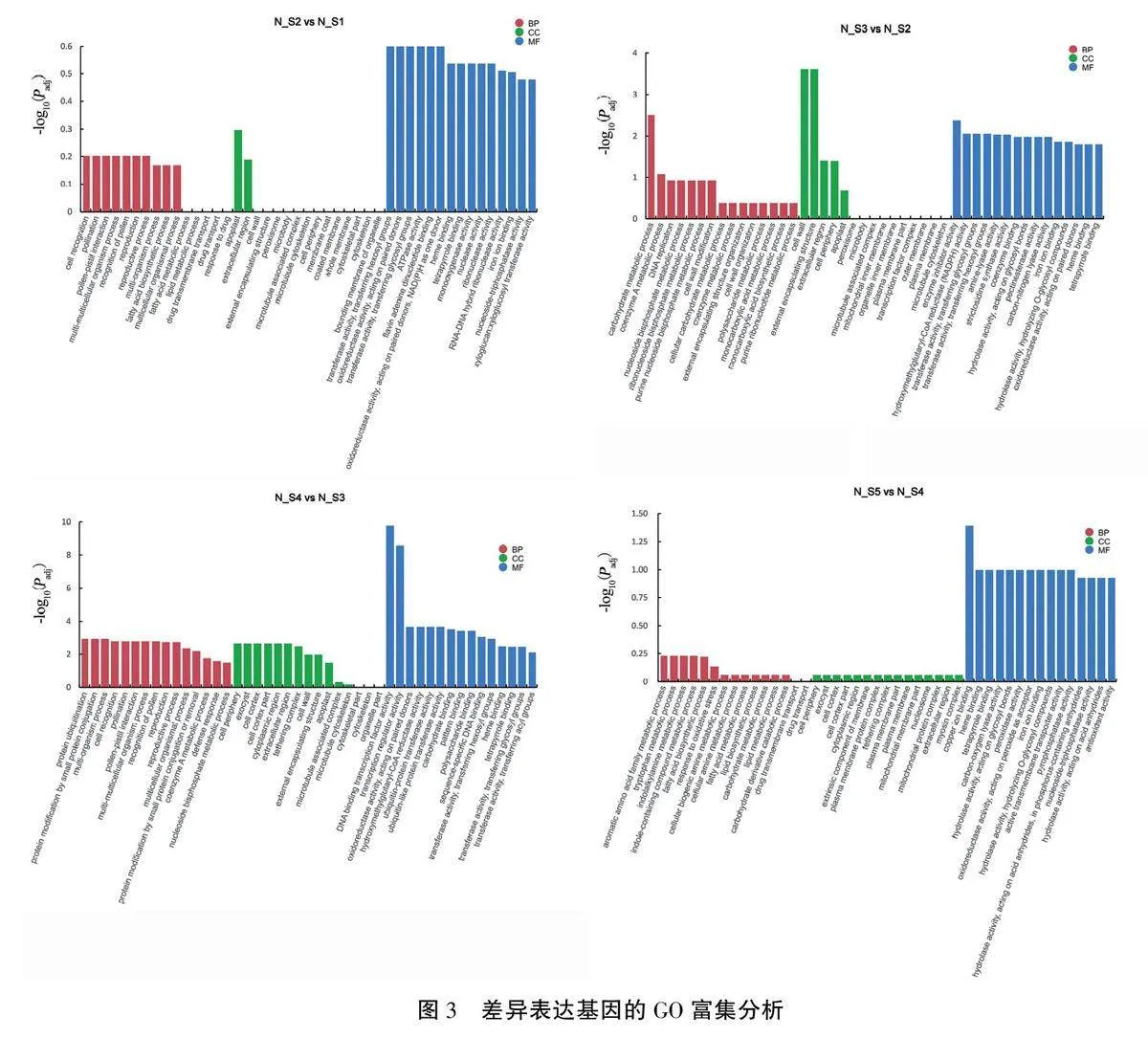
我们将在两组样品中的表达量差异达到两倍以上且Padjlt;0.05的基因确定为显著差异表达基因,对其进行GO富集分析,结果如图3所示。
从N_S1期到N_S2期,差异基因显著性较低,未富集到显著差异表达基因。
从N_S2期到N_S3期,共有750个差异基因可在GO数据库中得到注释,其中上调300个,下调450个。差异基因在生物过程(biological process,BP)、细胞组成(cellular component,CC)和分子功能(molecular function,MF)方面均有富集,BP方面参与最多的为碳水化合物代谢过程(carbohydrate metabolic process),共有88个差异基因;CC方面参与最多的为细胞壁(cell wall)、外部封装结构( external encapsulating structure)、细胞外周(cell periphery)和胞外区(extracellular re-gion),差异基因数量分别为18、18、21个和10个:MF方面差异基因最多的为葡萄糖基转移酶活性(transferase activity, transferring glycosyl groups)、己糖转移酶活性(transferase activity,transferring hexosyl groups)、血红素结合(heme binding)和四吡咯结合(tetrapyrrole binding),差异基因数量分别为71、61、57个和57个。
从N_S3期到N_S4期,共有641个差异基因可在GO数据库中得到注释,其中上调608个,下调33个。该时期基因表达差异最为显著,在BP、CC和MF方面均有富集。BP方面参与最多的为多有机体过程(multi-organism process)、细胞识别(cell recognition)、授粉(pollination)、花粉一雌蕊相互作用( pollen-pistil interaction)、花粉识别(rec-ognition of pollen)和多细胞生物过程(multicellularorganismal process),差异基因数量分别为20、18、18、18个和18个;CC方面参与最多的为细胞外周,有9个差异基因;MF方面参与最多的为DNA结合的转录因子活性(DNA binding transcriptionfactor activity)、转录调节因子活性(Transcription regulator activity)和作用于成对供体的氧化还原酶活性(oxidoreductase activit, acting on paired donors),差异基因数量分别为38、38个和29个。
从N_S4期到N_S5期,差异基因主要集中于MF方面,参与最多的为铜离子结合(copper ion binding),有10个差异基因。
2.3差异基因的KEGG通路富集分析
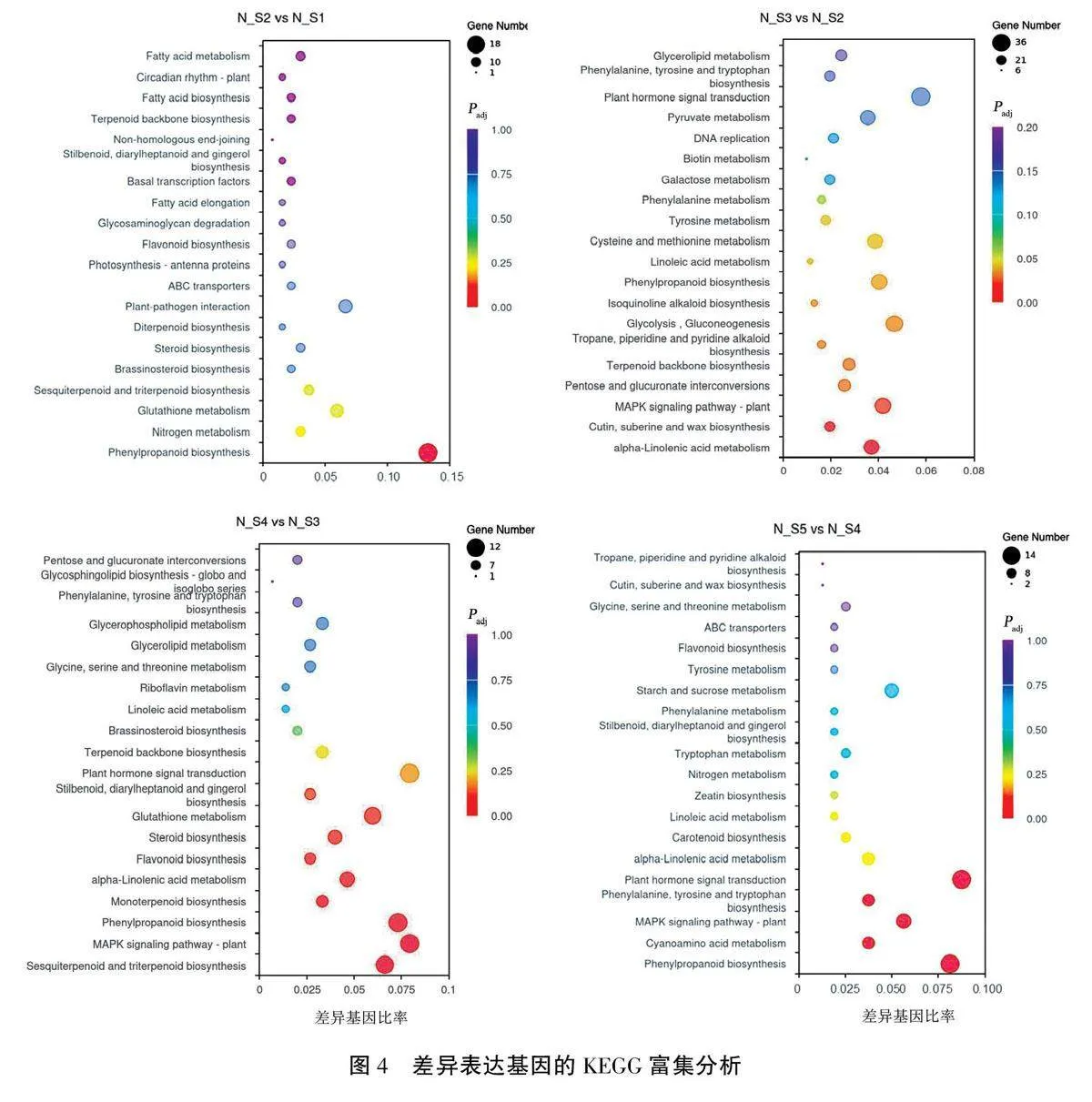
采用clusterProfile软件对4个比较组(N_S2vs N_S1、N_S3 vs N_S2、N_S4 vs N_S3、N_S5 vsN_S4)的差异基因进行KEGG通路富集分析,用Padjlt;0.05作为确定显著性富集的阈值,结果见图4。图中展示了每个对比组显著性最高的20个通路,横坐标表示注释到KEGG通路的差异基因数与总差异基因数的比率,纵坐标代表富集到的KEGG通路:点的大小表示注释到KEGG通路的基因数大小,颜色由红色到紫色代表了富集到的基因的显著性由大到小。
在N_S1期到N_S2期,有202个基因可在KEGG数据库中得到注释,共富集到81条通路,其中显著富集1条通路,即苯丙烷类生物合成通路(phenylpropanoid biosynthesis),共包含18个差异基因。
在N_S2期到N_S3期,共有1108个基因可在KEGG数据库中得到注释,共富集到115条通路,显著富集12条通路。其中富集差异基因较多的是糖酵解/糖异生(glycolysis,gluconeogenesis)、植物MAPK信号通路(MAPK signaling pathway-plant)、苯丙烷类的生物合成和a-亚麻酸代谢(alpha-linolenic acid metabolism),分别含有29、26、25、23个差异基因。
在N_S3期到N_S4期之间,共有227个基因在KEGG数据库中得到了注释,共富集到77条通路,有8条通路显著富集,其中植物MAPK信号通路、苯丙烷类的生物合成以及倍半萜和三萜的生物合成(sesquiterpenoid and triterpenoid biosyn-thesis)通路是富集差异基因多的通路,分别含有12、11、10个差异基因。
在N_S4期到N_S5期,共254个基因在KEGG数据库中有注释,共有88条通路被富集,5条通路显著富集。其中苯丙烷类生物合成和植物激素信号转导(plant hormone signal transduction)是富集差异基因最多的通路,分别包含14个和13个差异基因。
2.4类黄酮与花青素合成相关基因的挖掘与分析
根据转录组数据的分析发现,类黄酮与花青素合成的相关基因中有15个基因在“紫霞仙子”月季花的不同发育时期差异表达。其中,关键基因CHI、CHS、DFR、F3H、A3GT5随着花的发育表达量先上升后下降,CHI、CHS、F3H在半开期达到峰值,而DFR、A3GT5在盛开期达到峰值;MYB转录因子基因MYB6和MYB113与上述基因表达趋势相同,可能也对类黄酮及花青素的合成存在调控作用(图5)。
2.5类胡萝卜素合成相关基因的挖掘与分析
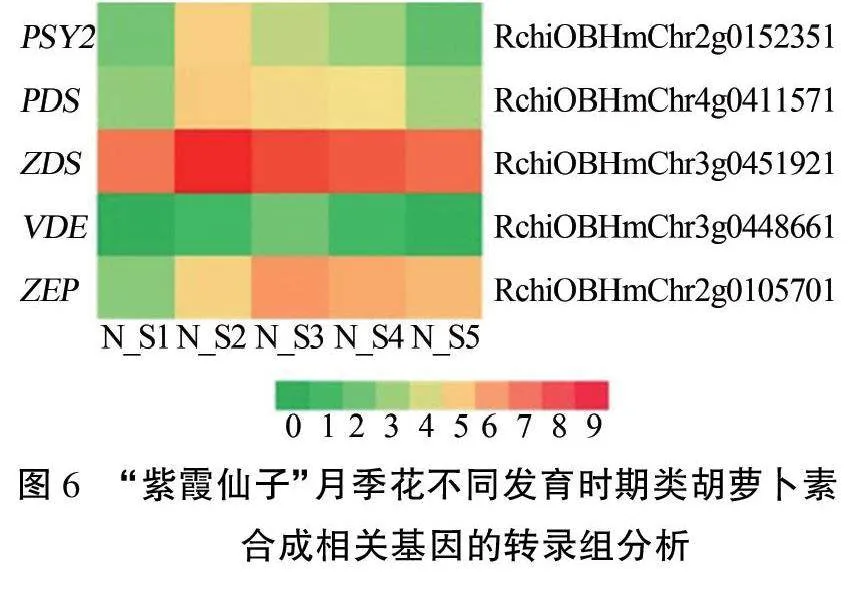
根据转录组数据的分析发现,类胡萝卜素合成的相关基因中有14个在“紫霞仙子”月季花的不同发育时期差异表达。其中,关键基因PSY2、PDS、ZDS、VDE和ZEP随着花的发育表达量先上升后下降,PSY2、PDS、ZDS在初开期表达量达到峰值,VDE和ZEP则在半开期达到峰值(图6)。
3讨论与结论
本研究通过转录组测序分析的方法,分析了“紫霞仙子”月季花不同发育时期影响花色变化的差异基因,结果表明,初开期到半开期的差异表达基因最多,共3341个:半开期到盛开期的差异基因最少,共980个。证明初开期到半开期是“紫霞仙子”月季花生理活动最为活跃的时期。整个花发育过程中均与苯丙烷类的合成密切相关。从花蕾期到半开期,均与己糖转移酶活性和葡萄糖基转移酶活性的KEGG通路密切相关,证明糖代谢参与到“紫霞仙子”月季花的开放过程中。有研究表明糖的转运与代谢可促进牡丹解除休眠和二次开花,这也可以佐证本研究在“紫霞仙子”月季中得到的结果。从初开期到盛开期,均与植物MAPK信号通路的KEGG通路密切相关。从盛开期到落花期,植物激素信号转导的KEGG通路都与花的发育阶段密切相关,证明在花发育后期植物激素发挥着重要作用。有研究表明茉莉酸、水杨酸和脱落酸可能协同调控杜鹃花的衰老过程,这也佐证了本研究结果。
在研究类黄酮与花青素合成通路的基因变化规律时,共找到15个基因在“紫霞仙子”月季花不同发育时期差异表达,其中,CHI、CHS、F3H基因表达在半开期达到顶峰,证明半开期是“紫霞仙子”月季大量积累二氢黄酮醇的时期:而DFR作为合成无色矢车菊素、无色天竺葵素和无色飞燕草素的重要基因,A3GT5作为对花青素进行糖基化修饰的糖基转移酶,二者的表达量均在盛开期达到顶峰,这一时期也是“紫霞仙子”月季花中花青素快速积累的时期。有研究表明MYB6与MYB113作为R2R3MYB转录因子,可以正向调控植物花中的花青素合成;本研究结果也表明MYB6与MYB113可能在“紫霞仙子”月季花发育过程中参与花青素合成的调控。
本研究在对类胡萝卜素合成相关基因的分析中找到了14个基因在“紫霞仙子”月季花不同发育时期差异表达。其中,PS Y2、PDS、ZDS基因表达量在初开期达到峰值,随后逐步下降,这可使植物积累的类胡萝卜素减少。在某些橙色和黄色花卉中,类胡萝卜素的表达量从花蕾期到落花期始终居高不下,如龙胆。“紫霞仙子”作为蓝紫色月季花,其类胡萝卜素合成基因从早期就开始下调,推测可能是通过减少类胡萝卜素在体内的积累降低黄色对蓝紫色花色的干扰。而VDE和ZEP基因的表达量在半开期达到峰值,随后逐渐下降,可能是由于这两个基因位于类胡萝卜素合成通路的下游,存在一定的响应延迟。
综上所述,本研究通过“紫霞仙子”月季花发育不同时期的转录组分析,找出了开花过程中类黄酮与花青素、类胡萝卜素合成通路中的差异基因,探讨了其花色形成机理,为日后月季的花色改造奠定了理论基础。
猜你喜欢
保健与生活(2021年13期)2021-07-14 00:50:04
中国果树(2020年2期)2020-07-25 02:14:12
天然产物研究与开发(2018年11期)2018-11-30 02:06:46
天然产物研究与开发(2018年11期)2018-11-30 02:06:28
中成药(2017年5期)2017-06-13 13:01:12
现代农业研究(2017年1期)2017-03-20 18:53:56
中成药(2016年8期)2016-05-17 06:08:41
中国科技博览(2016年5期)2016-04-23 10:56:35
食品界(2016年4期)2016-02-27 07:37:20
中国康复理论与实践(2015年10期)2015-12-24 05:42:43
