
辣椒CaWRKY39基因克隆及其酵母单杂交pGADT7载体构建
2024-12-31 00:00:00夏玉琪郑贤涛武士波刘雅婷
山东农业科学 2024年7期


关键词:辣椒;CaWRKY39;生物信息学分析;酵母单杂交pGADT7载体构建
辣椒(Capsicum annuum L.)是茄科辣椒属植物,是我国重要的经济作物之一。根据FAO数据,2021年我国辣椒种植面积达82.7万公顷,约占全球辣椒种植面积的40%,年产值超过3500亿元。但番茄斑萎病毒(Tomato spotted wilt virus,TSWV)的入侵严重影响了辣椒的产量和品质。TSWV是布尼亚病毒目(Bunyavirales)番茄斑萎病毒科(Tospoviridae)正番茄斑萎病毒属(Orthotospo-vzrus)的一种,是世界十大农业病害之一,每年造成的经济损失超过10亿美元。TSWV侵染辣椒后会使其株形矮化、株幅变小、果实严重畸形、单果重降低、结果数量及单果种子数减少。在我国,TSWV已快速从最初发现的四川和云南扩散至辽宁、甘肃、山东等14个省份,可侵染多种蔬菜作物,已成为我国辣椒和番茄等主产区导致绝产的主要病毒,造成了巨大的经济损失。因此,如何有效防控该病毒是保护辣椒产业健康稳定发展的重要问题。
已有研究表明,WRKY转录因子在作物抗病性中发挥重要作用。如:水稻WRKY13、WRKY42、WRKY45-2形成转录调控级联,其中WRKY42可能通过抑制茉莉酸(JA)信号相关基因负向调控水稻对稻瘟病菌的应答,而WRKY45-2转录激活WRKY13,WRKY13的编码蛋白则反过来转录抑制WRKY42,从而调控水稻对稻瘟病菌的抗性:过表达苹果MdWRKY100后,其抗炭疽病菌能力增强,而RNAi基因沉默植株对炭疽病菌更敏感,表明MdWRKY100在苹果响应炭疽病中发挥正调控作用:CmWRKY15-J对菊花白锈病原菌掘氏菊柄锈菌具有类似正调控功能;桃PpWRKY45可以通过结合激活抗性基因PpCHI、PpGLU、PpPR-like等启动子中的W-box元件并激活其表达,增强对匍枝根霉菌的耐受性:OsWRKY114能直接结合几丁质酶的启动子激活其表达,从而降低水稻对水稻白叶枯病菌的敏感性。近年来,WRKY转录因子在辣椒中的研究也取得了一定的进展。报道显示,CaWRKY6、CaWRKY15、CaWRKY30等转录因子可能参与烟草花叶病毒、辣椒青枯病菌、辣椒疫霉菌和根结线虫等多种病原体侵染后的植物防御机制。以上研究表明,WRKY转录因子可以广泛参与植物的各种生物和非生物胁迫过程,并且功能具有多样性,但目前关于WRKY转录因子是否参与抗TSWV过程以及如何发挥作用尚无报道。
本课题组前期通过TSWV病毒侵染及转录组学分析发现,侵染后的辣椒叶片中特异性高表达,推测其可能在抗TSWV中发挥重要作用,但具体功能和作用机制尚不清楚。本研究从辣椒叶片中克隆基因,利用生物信息学方法对CaWRKY39蛋白的理化性质、结构和功能进行综合分析,并构建的酵母单杂交pGADT7表达载体,以期为进一步解析CaWRKY39抗TSWV的分子机理提供技术支撑和参考。
1材料与方法
1.1材料
本试验所用辣椒品种是‘萧新19’,种子购于安徽萧新种业有限公司。
载体、菌株及试剂:酵母单杂交表达载体pGADT7(Amp抗性)为云南省作物生产与智慧农业重点实验室自有;限制性内切酶购于纽英伦生物技术(北京)有限公司(NEB);质粒提取试剂盒购于艾科瑞生物工程(湖南)有限公司(AG);TRNzol Universal总RNA提取试剂购于天根生化科技(北京)有限公司( TIANGEN);胶回收试剂盒、PCR产物回收试剂盒、DL2000 DNAMarker、DL10000 DNA Marker、In-Fusion Snap As-sembly cloning kits购于宝生物工程(大连)有限公司(TaKaRa);pEASY-Blunt Zero Cloning Kit购于全式金生物技术股份(北京)有限公司(Trans-Gen);Hiscript Ⅱ1st Strand cDNA Synthesis Kit、2x Phanta Flash Master Mix购于诺唯赞生物科技(南京)有限公司(Vazyme);CaWRKY39基因克隆引物和载体构建引物合成及阳性菌落测序均由生工生物工程(上海)股份有限公司(Sangon)完成。
1.2方法
1.2.1植物材料的培育与处理
于2023年3月将辣椒种子播于穴盘中,待出苗后移栽到装有土壤和腐殖质混合物的塑料盆(10cm×10cm×10cm)中,置于光照培养箱,在(25+2)℃、相对湿度恒定为(65±5)%、光照周期为光照16h:黑暗8h条件下培养,每2d浇水一次,保证植株正常生长;待植株生长至四叶期,采集叶片,立即在液氮中冷冻后保存在-80℃冰箱中。
1.2.2CaWRKY39基因克隆
利用TRNzol Uni-versal总RNA提取试剂于2023年6月提取辣椒叶片的总RNA,经反转录试剂盒合成cDNA,并以该cDNA为模板进行PCR扩增。根据前期转录组分析得到的CaWRKY39编码序列利用PrimerPremier 6.0软件设计基因克隆引物(表1)。PCR反应体系:终延伸1min,4℃保存。利用胶回收试剂盒回收纯化CaWRKY39全长CDS序列片段,将纯化产物连接至克隆载体pEASY-Blunt Zero Cloning Vector上,转化至大肠杆菌感受态细胞Transl-T1,鉴定的含有CaWRKY39片段的阳性菌落送生工生物工程(上海)股份有限公司测序。
1.2.3CaWRKY39蛋白的生物信息学分析基于测序获得CaWRKY39基因的碱基序列,利用DNAMAN6.0推导CaWRKY39蛋白序列,利用ExPASy-ProtParam(https://web. expasy. org/prot-param/)分析CaWRKY39蛋白的理化性质,通过SOPMA软件(https://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma. html)预测分析其二级结构.通过SWISS-MODEL软件预测其三级结构。利用在线预测工具NetPhos 3.1(https://services.healthtech.dtu.dk/services/NetPhos-3.11)、TMHMM Server 2.0(https://services.healthtech.dtu.dk/services/TMHMM-2.01)、SignalP-4.1(http://www.cbs.dtu.dk/services/SignalP-4.1/),Cell-PLoc2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)等在线工具分析CaWRKY39蛋白的磷酸化位点、跨膜结构、信号肽以及蛋白的亚细胞定位。利用NCBI数据库搜索并下载其他物种的WRKY39蛋白序列,用MEGA11软件构建系统进化树(Neighbor-join-ing法,Bootstrap重复数为1000,线程数量为3)。
1.2.4 CaWRKY39基因表达载体构建和酶切鉴定
通过SnapGene软件设计带酶切位点NdeI、EcoRI插入片段的同源重组引物(表1),以pEASY-CaWRKY39质粒DNA为模板。PCR反应体系:质粒DNA1.0uL,上、下游引物各1.5uL,2xPhanta Flash Master Mix 25.0uL, ddH20 21.0uL,共50.0uL。PCR反应程序:98℃预变性30s;98℃变性10s,54℃退火5s,72℃延伸5s,34个循环;72℃终延伸1min,4℃保存。1%琼脂糖凝胶电泳检测并利用DNA胶回收试剂盒(TaKaRa,宝日医生物技术(北京)有限公司)回收目的基因片段。用限制性内切酶NdeI、EcoRI酶切载体pGADT7质粒5h后进行琼脂糖凝胶电泳并将酶切产物回收,按照无缝克隆试剂盒(TaKaRa,宝日医生物技术(北京)有限公司)说明书将酶切纯化后的pGADT7载体与回收的带同源臂CaWRKY39片段进行连接,连接产物转化大肠杆菌感受态细胞DH5a,利用含氨苄抗性的LB平板培养基过夜筛选,经PCR鉴定阳性的菌落送生工生物工程(上海)股份有限公司测序。构建的表达载体命名为pGADT7-CaWRKY39。挑选测序正确的菌落采用质粒提取试剂盒提取pGADT7-CaWRKY39质粒DNA,以该质粒DNA为模板进行PCR鉴定后,琼脂糖凝胶电泳检测。
2结果与分析
2.1CaWRKY39基因克隆
基于转录组数据得到的CaWRKY39基因CDS序列,利用DNAMAN6.0对其蛋白序列进行推测分析,结果(图1)显示,CaWRKY39的CDS序列全长为1035bp,共编码344个氨基酸,含有一个由WRKYGQK7个氨基酸组成的保守WRKY结构域,说明CaWRKY39蛋白属于WRKY成员。本实验以辣椒‘萧新19’叶片的cDNA为模板进行PCR扩增,琼脂糖凝胶电泳检测显示扩增产物与预期大小一致(图2),通过回收目的片段、转化大肠杆菌、单克隆PCR扩增和测序鉴定,结果表明成功克隆到CaWRKY39基因全长CDS序列。
2.2CaWRKY39蛋白基本理化性质分析
ExPASy-ProtParam在线分析结果显示,CaWRKY39蛋白的分子式为C1642 H2653 Nsis 0518 S21,总原子数为5349个,分子量为38570.53Da.等电点为9.59,带正电荷残基为30个,带负电荷残基为49个,总平均亲水性为-0.805,不稳定系数为59.71,脂肪族指数为63.17。表明CaWRKY39蛋白属于不稳定的碱性亲水蛋白。
2.3CaWRKY39蛋白二级结构、三级结构分析
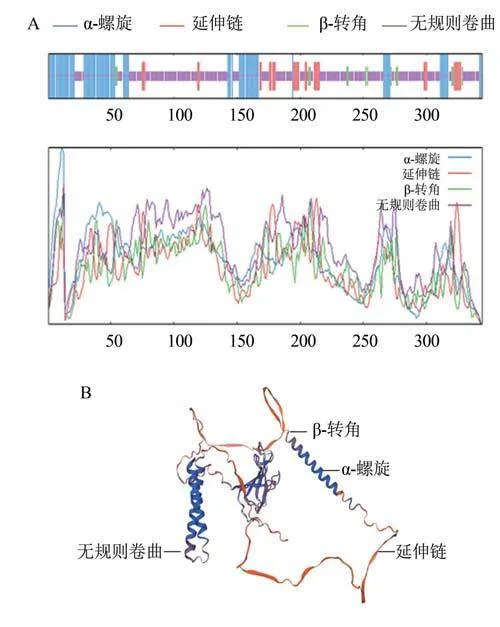
SOPMA分析结果显示,CaWRKY39蛋白的二级结构中无规则卷曲占60.76%,a-螺旋占25.58%,延伸链占9.59%,转角占4.07%(图3A)。进一步通过SWISS-MODEL网站预测CaWRKY39蛋白的三维结构,同样发现该蛋白无规则卷曲占比最大,同时还存在a-螺旋、延伸链和转角等结构(图3B)。表明CaWRKY39蛋白主要由无规则卷曲组成,其次为仪-螺旋,延伸链、转角占比较小,这可能与CaWRKY39蛋白的功能相关。
2.4CaWRKY39蛋白磷酸化位点、信号肽、跨膜结构域、亚细胞定位分析
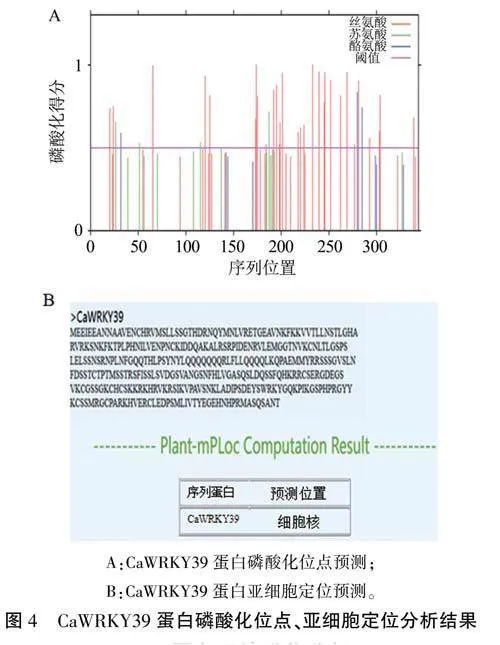
磷酸化对蛋白质功能的正常发挥起着重要的调节作用。经NetPhos 3.1分析发现,CaWRKY39蛋白由38个氨基酸磷酸化位点构成,包含30个丝氨酸(Serine)、5个苏氨酸(Threonine)和3个酪氨酸(Tyrosine)磷酸化位点(图4A)。根据CaWRKY39蛋白相应的磷酸化潜能,推测其可能被丝氨酸、苏氨酸和酪氨酸激酶磷酸化激活,从而调控基因的表达。
亚细胞定位分析结果显示CaWRKY39蛋白定位于细胞核(图4B),说明其为典型的核转录因子。但进一步分析发现其既没有跨膜区域,也不存在信号肽,说明CaWRKY39蛋白不是定位于生物膜上的膜蛋白,无跨膜运输,不属于分泌蛋白。
2.5CaWRKY39蛋白系统进化分析
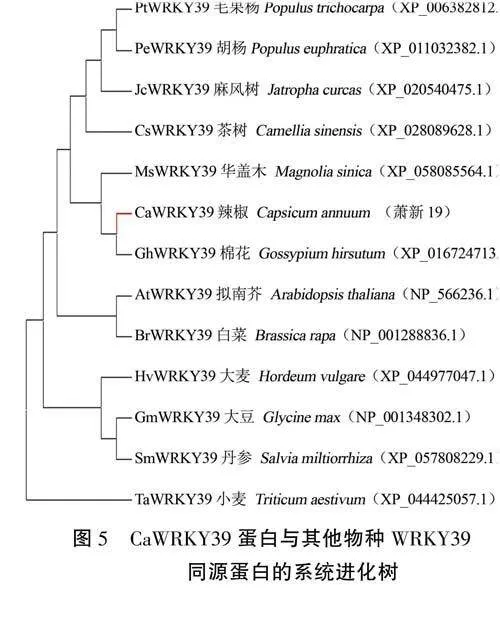
根据CaWRKY39蛋白的序列信息,在TAIR数据库下载AtWRKY39蛋白序列,在NCBI数据库搜索并下载其他11个物种的WRKY39蛋白序列,利用MEGA11软件,采用邻接法构建系统进化树。结果(图5)发现,辣椒CaWRKY39与棉花GhWRKY39的亲缘关系最近,与小麦TaWRKY39的亲缘关系较远,预示着CaWRKY39蛋白与Gh-WRKY39具有相似功能。
2.6表达载体的构建及验证
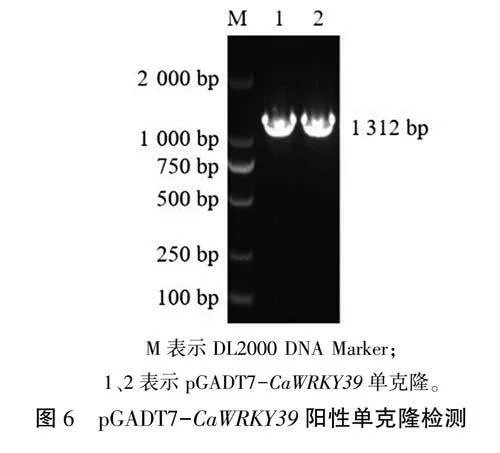
为进一步验证CaWRKY39对相关抗性基因的靶向调控作用,进行pGADT7-CaWRKY39酵母单杂交表达载体构建。以pEASY-Ca WRKY39质粒DNA为模板,用CaWRKY39-Nde I-F、CaWRKY39-EcoR I-R引物进行CaWRKY39基因带同源臂片段扩增,对扩增产物和NdeI、EcoRI双酶切后的线性pGADT7载体进行纯化回收,随后利用无缝克隆试剂盒将目的基因片段导人线性pGADT7载体中,将连接产物转入大肠杆菌感受态细胞,挑取单菌落进行PCR检测和测序鉴定。结果表明,目的片段大小为1312bp(图6),与预期相符,表明pGADT7-CaWRKY39酵母单杂交表达载体构建成功,可以用于后续实验。
3讨论
目前关于辣椒WRKY类转录因子的基因克隆及相关功能研究报道较为少见。本研究成功克隆了辣椒CaWRKY39全长CDS序列,全长1035bp,共编码344个氨基酸;CaWRKY39蛋白总平均亲水性为-0.805,不稳定系数为59.71,等电点为9.59.属于不稳定的亲水性碱性蛋白,这与已报道的CaWRKY30蛋白的预测结果类似。CaWRKY39蛋白结构中无规则卷曲占比60.76%,说明其结构不稳定。蛋白磷酸化会降低该蛋白的稳定性,CaWRKY39蛋白的磷酸化位点有38个,再次验证了该蛋白的不稳定性。
转录因子在细胞核内才能发挥调控功能。本研究结果显示CaWRKY39蛋白定位于细胞核,表明其可以发挥转录因子的调控功能。但该蛋白无跨膜区域和信号肽,是非分泌蛋白,形成后会在细胞内发挥作用并根据植物体需求调整数量和状态。WRKY转录因子最显著的功能是参与植物抗病调节过程。棉花GhWRKY39在本氏烟中过表达后,PR抗性基因的表达显著上调,青枯菌的侵染病症明显减弱,表明GhWRKY39具有抗青枯病的作用。从系统进化树可知,CaWRKY39与GhWRKY39亲缘关系最近,并且CaWRKY39在TSWV侵染下表达量显著提高,推测CaWRKY39具有抗TSWV功能。
本研究利用无缝克隆技术成功构建了酵母单杂交表达载体pGADT7-CaWRKY39,下一步将结合酵母单杂交诱饵载体构建和相关互作实验,进一步验证CaWRKY39对CaPR1等抗性基因的靶向调控作用,以阐明CaWRKY39转录因子参与辣椒抗TSWV病毒的分子机理,进而为抗TSWV病毒植物培育和绿色防控提供参考。
4结论
本研究对CaWRKY39全长CDS及其编码蛋白的理化性质、结构和系统进化等进行了生物信息学分析,并采用RT-PCR技术克隆得到CaWRKY39基因全长CDS序列,进一步通过无缝克隆的方法成功构建了其酵母单杂交pGADT7表达载体,可为全面深入研究CaWRKY39转录因子参与辣椒抗TSWV病毒分子机理提供技术支持,并为抗TSWV病毒植物培育和绿色长效防控提供理论依据。
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
小哥白尼(神奇星球)(2021年6期)2021-07-28 06:31:36
今日农业(2020年24期)2020-12-15 16:16:00
中外文摘(2020年9期)2020-06-01 13:47:56
天津医科大学学报(2019年6期)2019-08-13 07:04:42
兽医导刊(2016年12期)2016-05-17 03:51:50
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
