
阔叶红松林下幼苗、幼树定植生境散射辐射特征
2024-12-12 00:00:00杜昕董雪谷会岩陈祥伟
南京林业大学学报(自然科学版) 2024年6期
关键词:光照强度




摘要:【目的】为探明阔叶红松林下常见乔木树种更新幼苗、幼树的光强需求,分析了各树种更新对林下光强的响应差异,进而为阔叶红松林及相关生态系统的科学经营提供支持。【方法】以凉水国家级自然保护区内4块阔叶红松林调查样地林下红松(Pinus koraiensis)、红皮云杉(Picea koraiensis)、臭冷杉(Abies nephrolepis)、春榆(Ulmus davidiana)、色木槭(Acer pictum)、青楷槭(Acer tegmentosum)、花楷槭(Acer ukurunduense)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)9种常见乔木更新的幼苗、幼树为研究对象,在个体水平上计算各株更新幼苗、幼树定植生境的加权进界邻体散射荫蔽度(WINDSD),计算各树种幼苗、幼树定植生境的WINDSD平均值、标准差及分布倾向度。以Mann-Whitney U检验分析各树种幼树、幼苗定植生境的WINDSD与阔叶红松林下均匀布设样点间的差异显著性及同种树种幼苗与幼树定植生境WINDSD间的差异显著性;同时,以蒙特卡罗法检验各类乔木更新幼苗、幼树种群定植生境与阔叶红松林下同等规模的随机分布种群定植生境的WINDSD分布倾向度间的差异显著性。【结果】阔叶红松林下均匀布设样点的WINDSD均值与标准差分别为0.538及0.213,分布倾向度为1.069。红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、花楷槭、紫椴、水曲柳9种乔木树种的更新幼树、幼苗中,仅有春榆幼树及水曲柳幼树定植生境的WINDSD平均值与分布倾向度小于林下均匀布设样点。基于均值与分布倾向度共同判断,红松、红皮云杉、臭冷杉及紫椴更新幼树定植生境中的散射辐射强度弱于更新幼苗定植生境;春榆、色木槭、青楷槭、花楷槭、水曲柳更新幼树定植生境的散射辐射强度强于更新幼苗定植生境。蒙特卡罗检验与Mann-Whitney U检验共同显示,红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、水曲柳更新幼苗定植生境的WINDSD显著大于阔叶红松林下均匀布设样点,花楷槭、紫椴更新幼苗定植生境的WINDSD与林下均匀布设样点无显著差异;红松、红皮云杉、臭冷杉更新幼树定植生境的WINDSD显著大于阔叶红松林下均匀布设样点,而其余树种更新幼树定植生境的WINDSD与林下均匀布设样点无显著差异。Mann-Whitney U检验显示,9种乔木树种中春榆、色木槭幼树定植生境WINDSD与对应树种更新幼苗定植生境WINDSD间的差异达到显著水平(Plt;0.05)。【结论】阔叶红松林主要乔木更新幼苗及幼树倾向分布于林下光强较弱的区域内。相较于更新幼苗,更新幼树定植的生境内光强总体上相对更强。不同树种更新对林下光强差异的响应有所不同,红松、红皮云杉、臭冷杉幼苗与幼树定植生境的光强差异不明显,而春榆、色木槭、青楷槭幼树定植生境的光照强度明显强于对应树种幼苗的定植生境。
关键词:阔叶红松林;森林天然更新;光照强度;加权进界邻体散射荫蔽度;蒙特卡罗检验
中图分类号:S711;S754""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)06-0145-12
Diffuse radiation environment of regeneration seedlings and saplings under a broadleaved-Korean pine forest
DU Xin1,DONG Xue1,GU Huiyan1,2,CHEN Xiangwei1,2*
(1. Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, Key Laboratory of National Forestry and Grassland Administration on Northeastern Forest Silviculture, College of Forestry, Northeast Forestry University, Harbin 150040, China; 2. Liangshui National Key Field Observation and Research Station for Forest Ecosystem, Yichun 153000, China)
Abstract: 【Objective】 This study aims to determine the light intensity requirements for the regeneration of seedlings and saplings in mixed broadleaved-Korean pine (Pinus koraiensis) forests. Additionally, it seeks to investigate how the regeneration of different tree species responds to variations in light intensity. This research guides the scientific management and conservation efforts of broadleaved-Korean pine forests and their ecosystems.【Method】The research focused on nine common tree species: Pinus koraiensis, Picea koraiensis, Abies nephrolepis, Ulmus davidiana, Acer pictum, Acer tegmentosum, Acer ukurunduense, Tilia amurensis, and Fraxinus mandshurica, across four plots in the Liangshui National Nature Reserve. The weighted inside-boundary neighbor diffuse shading degree (WINDSD) was computed for the planting habitats of each seedling and sapling. The mean, standard deviation, and distribution tendency degree of WINDSD were calculated for the planting habitats of each tree species’ seedlings and saplings. The Mann-Whitney U test was employed to identify significant differences in WINDSD among the planting habitats of each tree species and uniformly distribute sample points beneath the broadleaved-Korean pine forest canopy. The analysis is extended to compare the differences in WINDSD among the planting habitats of seedlings and saplings for each tree species. Additionally, the Monte Carlo method was utilized to ascertain significant differences in the distribution tendency degree of WINDSD between the planting habitats of seedlings and saplings for various tree species populations and a random distribution population of equivalent size for each tree species.【Result】The analysis revealed that the mean and standard deviation of WINDSD for the uniformly distributed sample points of the broadleaved-Korean pine forest were 0.538 and 0.213, respectively, with a distribution tendency degree of 1.069. Among the seedlings and saplings of the nine tree species studied, only Ulmus japonica and Fraxinus mandshurica saplings exhibited a lower average value and distribution tendency degree of WINDSD compared to the uniformly distributed sample points in the forest. The analysis also indicated that the diffuse radiation intensity within the saplings’ planting habitats for Pinus koraiensis, Picea koraiensis, Abies nephrolepis, and Tilia davidiana was weaker compared to those designated for seedling regeneration. The Monte Carlo and Mann-Whitney U tests highlighted that the WINDSD in the planting habitats of seedlings for Pinus koraiensis, Picea koraiensis, Abies nephrolepis, Ulmus davidiana, Acer pictum, Acer tegmentosum, and Fraxinus mandshurica was significantly higher than that of the uniformly distributed sample points. In contrast, the WINDSD values for the planting habitats of Acer ukurunduense and Tilia amurensis seedlings did not significantly differ from those of the uniformly distributed sample points in the forest. For saplings, the WINDSD values in the planting habitats of Pinus koraiensis, Picea koraiensis, and Abies nephrolepis were significantly higher compared to the uniformly distributed sample points, whereas those for the saplings of other species did not significantly differ. Additionally, the Mann-Whitney U test indicated significant differences in WINDSD between the planting habitats of saplings and seedlings for Ulmus japonica and Acer mono (Plt;0.05). 【Conclusion】The findings suggest that seedlings and saplings of the primary tree species in the broadleaved-Korean pine forest are usually found in areas with weaker light on the forest floor. Compared to seedlings, saplings tend to establish in habitats with slightly stronger light intensity. Different tree species exhibit varied responses to light intensity variations under the forest canopy. The differences in light intensity between habitats for seedlings and saplings are not pronounced for Pinus koraiensis, Picea koraiensis, and Abies nephrolepis, whereas the light intensity in the sapling habitats of Ulmus davidiana, Acer pictum, and Acer tegmentosum is significantly higher than in their seedling regeneration areas.
Keywords:broadleaved-Korean pine forest; natural forest regeneration; light intensity; weighted inside-boundary neighbor diffuse shading degree (WINDSD); Monte Carlo test
森林天然更新是林木通过自身繁殖能力,形成新一代幼林的过程,是森林生态系统自我繁衍、维持的手段,是森林群落动态的重要部分[1]。更新幼苗和幼树的定植及生长对种群的数量、发展乃至群落的结构组成均具有直接影响。林下光环境影响更新幼树、幼苗的生理特性与性状特征,及其发生、生长与死亡[2-5]。林分中,地表光环境的空间异质性是冠层对林冠上均质光气候再分配的结果[6],林隙、林冠下、林隙/冠层界面(林缘)生境的光环境特征具有明显差异[7],而冠层结构异质性造成的林下光环境多样性为林下更新的生态位分化提供了环境基础,是维持林下物种多样性的重要保证[8]。不同的植物具有不同的生理、性状特征,及其独特的光因子生态位[9-11],对林下光环境变化具有不同的响应策略[12-14]。探明不同树种更新幼树、幼苗的光环境需求,对理解生态系统的运行和群落组配机制、维持生态系统健康和稳定、诱导及促进群落进展演替等具有重要意义。
阔叶红松林是具有同龄斑块镶嵌结构的复层异龄混交林生态系统,而这种斑块镶嵌结构的形成与阔叶红松林林隙动态有关,阔叶红松林中光环境的时空动态是其系统维持稳定的内在驱动机制之一[15]。目前,对阔叶红松林光环境与更新关系已有了较多研究,如吴刚[16]探讨了林隙面积、林隙年龄与更新频率及更新物种组成间的关系;国庆喜[17]研究了林冠斑块与幼龄个体分布间的关系;刘妍妍等[18]研究了林隙与非林隙生境木本植物更新的密度、生长速率、新增量、死亡率的差异;Jin等[19]从不同发育阶段评价了红松林主要树种的林隙更新模式;Jin等[20]研究了林隙生境与非林隙生境中更新幼苗、幼树的物种多样性差异;董莉莉等[21]研究了不同间伐强度下冠层开阔度、叶面积指数及林下光辐射与林下更新数量特征间的关系等。目前,红松林下光环境与更新关系的研究多基于林冠/林隙分类、林隙面积分类或样地尺度冠层结构差异与林下光照差异。然而,林分地表光资源可用性为连续性环境变量,其与林隙大小、形态、林缘植株高度、林冠类型(针叶、阔叶)等均有一定关联[8],仅以林下微生境分类作为光环境梯度难以充分体现林下光环境的多样性。
太阳辐射可分为直射辐射与散射辐射,其相对比例受大气中云和气溶胶含量等的影响[22]。黑龙江省7月平均地表辐射量为158 kW·h/m2[23],而同期天文辐射约为460 kW·h/m2,故7月云量指数为0.343。基于该云量指数可由Frazer等[22]的经验公式求得7月黑龙江省散射辐射在总辐射中占比79.8%,即研究区夏季辐射以散射辐射为主。杜昕等[24]在对开敞度[25]进行改造的基础上建立了加权进界邻体散射荫蔽度(WINDSD),该光强测度指数能基于林分中林木空间分布坐标与树高模拟林下任意点位的散射辐射特征,在继承开敞度优点的基础上将指数的取值范围规划为0~1,在适当的参数选择下该指标与林下散射透射率具有强负相关关系,相较于开敞度其对林下散射透射率的模拟准确性提升超过200%。目前,WINDSD的计算方法已集成、开源于R包Understory.light(https://github.com/DuXinChina/Understory.Light)中。
本研究基于阔叶红松林的林分调查数据[24],在个体水平上计算阔叶红松林下红松(Pinus koraiensis)、红皮云杉(Picea koraiensis)、臭冷杉(Abies nephrolepis)、春榆(Ulmus davidiana)、色木槭(Acer pictum)、青楷槭(Acer tegmentosum)、花楷槭(Acer ukurunduense)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)共9种林分中数量最多的乔木树种更新幼苗、幼树定植生境的WINDSD,充分体现了林下光环境的连续性与多样性。研究旨在探索阔叶红松林中主要乔木树种更新幼苗、幼树定植生境的光环境特征,并分析各类树种更新幼苗、幼树分布规律的形成原因及内在生态机制,以期为阔叶红松林及其相关森林生态系统的科学经营提供理论与依据。
1 材料与方法
1.1 研究区概况
研究区为黑龙江省凉水国家级自然保护区(128°53′20″E,47°10′50″N),属小兴安岭南坡,达里带岭支脉东坡的凉水沟流域,为典型低山丘陵地貌。具有明显的温带大陆性季风气候,冬季寒冷干燥,夏季温热多雨,年平均气温-0.3 ℃,年平均最高气温7.5 ℃,年平均最低气温-6.6 ℃,年平均降水量676 mm,全年降雨多集中于6—8月,有效积温2 200~2 600 ℃,无霜期100~120 d。以阔叶红松林为代表的温带针阔混交林为研究区坡地地带性顶极植被;随坡位升高,土壤水分含量降低,林分中红松占比逐渐上升,林下土壤类型由潜育化暗棕色森林土向典型暗棕色森林土过渡。
1.2 样地布设与调查
在凉水自然保护区19林班内中、下坡位的阔叶红松林中平行等高线设置4块50 m×50 m 的样地(闭合差lt;1/200),各样地基本信息见表1。表1中树种蓄积组成以“十分法”表示,即各树种蓄积组成之和为10。当某一树种蓄积占比大于5%时其蓄积以数字表示,如1代表四舍五入后10%,2代表20%;如树种蓄积占比不足5%并大于2%时以“+”表示;树种蓄积占比小于2%并大于1%时以“-”表示。样地调查采用相邻格子法,将调查样地划分为100个5 m×5 m的调查单元,对达到起测胸径(5 cm)的林木进行每木检尺,以胸径尺测定胸径并以Vertex Ⅳ测树仪测定林木株高(精度为0.1 m),以皮尺测定冠幅并记录所属树种;记录林木在小调查单元中的相对位置,以便后续推算其在样地中的坐标。参考《东北、内蒙古天然次生林经营技术》(LY/T 1572—2000)中更新层的划定,将原始红松林内株高为[0.1,1.5) m的林木定义为更新幼苗[26];将株高大于等于1.5 m但未达到起测胸径的临界木作为更新幼树。逐株测定并记录更新幼苗幼树的树种、株高及其在小调查单元中的相对位置,以便后续推算坐标。各样地中,土壤取样按五点法进行。去除土壤表面杂草、枯落物与腐殖质后,挖开表土,在5~10 cm土层取土样,用于土壤含水率分析。
1.3 林下均匀布设样点及更新幼苗、幼树定植生境WINDSD的计算与概括性统计
1.3.1 WINDSD的计算
WINDSD是基于林木坐标与树高模拟林下散射辐射的光强测度指数,其通过林木树高与林木到研究点位的水平距离间的比值筛选参与指标计算的邻体林木;并以研究点位为中心划分4个象限,以“Levins生态位宽度指数”的形式构建了加权函数以表达研究点位周围邻体林木分布的均匀性。最终,WINDSD的取值被归一化为0~1,取值趋近于0代表邻体林木对研究点位的遮阴较弱,散射辐射强度较强;取值趋近于1说明邻体林木对研究点位的遮阴较强,研究点位散射辐射较弱。杜昕等[24]以爬山算法(hill climbing algorithm)对以WINDSD模拟阔叶红松林下散射辐射的参数进行寻优,得出在非二阶校正前提下,当局域临界角α为2.800、线性转换校正系数αline为0.5时,WINDSD与阔叶红松林下散射透射率间的Pearson及Spearman相关系数分别为-0.730与-0.694,其对林下散射辐射的模拟准确性相较于开敞度提升了237.3%。本研究中,将沿用上述参数组合。
为比较阔叶红松林下主要乔木树种更新幼树、幼苗定植生境的光环境倾向性,在设置5 m缓冲区以消除边缘效应后[27],分别在4块阔叶红松林样地下以2 m为间距均匀布设样点,形成21行×21列的点阵,计算各点的WINDSD,4块样地共布设1 764 个样点。同时计算林下更新数量最多的红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、花楷槭、紫椴、水曲柳9种乔木树种幼树、幼苗定植生境的WINDSD。
1.3.2 WINDSD的概括性统计
在计算阔叶红松林下均匀布设样点及主要乔木树种的幼树、幼苗定植生境的WINDSD后,结合各样地计算结果,计算样地下均匀布设样点及各树种幼树、幼苗定植生境WINDSD的均值、标准差及分布倾向度[28](distribution tendency degree,式中记为D)等概括性统计量。其中,分布倾向度用于评价幼苗与幼树定植生境散射辐射的倾向性。
D=∑Ni=1(N×ni)(N+1)/2×∑Ni=1ni。(1)
式中:N为将WINDSD的取值范围由0到1均等划分的份数,本研究中将WINDSD的取值范围划分为[0,0.1),[0.1,0.2),...,[0.8,0.9),[0.9,1]共10份;ni为第i份中幼苗、幼树的株数或均匀布设的样点数;D的取值范围为0到2,当Dgt;1时则幼树、幼苗或林下均匀布设样点的WINDSD倾向大于0.5,当Dlt;1时则幼树、幼苗或林下均匀布设样点的WINDSD倾向小于0.5。
1.4 更新幼苗、幼树定植生境WINDSD的差异显著性检验
1.4.1 更新幼苗、幼树定植生境与林下同等规模随机分布种群WINDSD差异的蒙特卡罗检验
在阔叶红松林下的1 764个样点中,有放回的抽取与各乔木树种更新幼苗、幼树数量相同的样点,并计算抽取样点的WINDSD分布倾向度。重复上述结果199次,绘制重复结果的核概率密度图,将核概率密度函数的2.5%~97.5%分位数范围作为阔叶红松林下与待检验种群具有同等种群规模的随机分布种群的WINDSD分布倾向度的取值范围。在2.5%与97.5%分位数处分别设置包络线,当乔木更新幼树或幼苗的WINDSD分布倾向度处于两包络线之间时,在Plt;0.05水平下幼苗或幼树定植生境的WINDSD与林下同等规模随机分布种群无显著差异;而当分布倾向度处于两包络线之外时,幼苗或幼树定植生境的WINDSD与林下同等规模随机分布种群具有显著差异。基于幼苗或幼树的WINDSD分布倾向度两侧的累积核概率密度,可计算检验显著性水平P。上述检验为蒙特卡罗双尾检验,用于判断幼苗、幼树定植生境的WINDSD是否与林下随机分布种群具有显著性差异,如需判定是否显著大于或小于同等规模的随机分布种群,可将P除以2,将检验转化为单尾。
1.4.2 更新幼苗、幼树定植生境WINDSD的Mann-Whitney U检验
采用Mann-Whitney U检验逐一分析研究涉及的9种乔木树种更新幼树、幼苗定植生境的WINDSD与林下均匀布设样点的WINDSD间的差异显著性;同时,以Mann-Whitney U检验分析相同树种更新幼树与幼苗定植生境的WINDSD间的差异显著性。
2 结果与分析
2.1 阔叶红松林下主要乔木树种幼苗与幼树的加权进界邻体散射荫蔽度特征
基于WINDSD模拟阔叶红松林下与红松林内9种主要乔木树种幼树、幼苗定植生境的散射辐射强度,结果如图1所示。
由图1可见,4块调查样地中各树种更新幼苗与幼树总体呈聚集分布,且多在林下WINDSD较高的位置形成聚集。
综合4块样地林下1 764个均匀分布样点计算阔叶红松林下WINDSD的均值为0.538,标准差为0.213,分布倾向度为1.069。红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、花楷槭、紫椴、水曲柳9种主要乔木树种更新幼苗及幼树的样本量及其定植生境的WINDSD的均值、标准差、分布倾向度见表2。
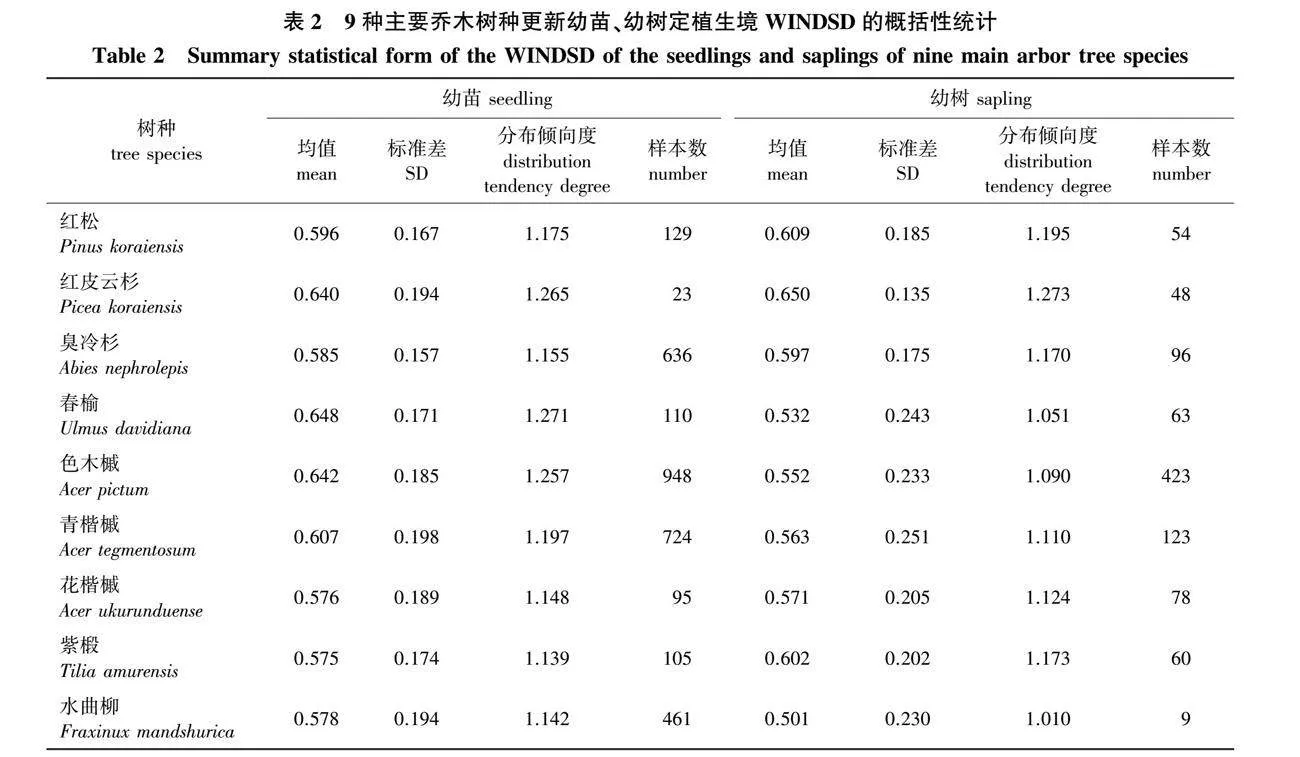
由表2可见,9种主要乔木更新幼苗定植生境WINDSD的均值与分布倾向度均大于阔叶红松林下均匀布设样点,这说明9种主要乔木树种更新幼苗主要分布在林下散射辐射较弱的区域内;同时,9种主要乔木树种更新幼苗定植生境WINDSD的标准差均小于阔叶红松林下均匀布设样点,这代表着在散射辐射强度梯度上,9种主要乔木树种更新幼苗在林下的分布位置存在特异性。9种主要乔木树种中,红松、红皮云杉、臭冷杉及紫椴的更新幼树定植生境WINDSD的均值与分布倾向度相较于对应树种的更新幼苗有所提升,其余树种更新幼树定植生境WINDSD的均值与分布倾向度相较于更新幼苗有所下降。9种主要乔木树种中,春榆及水曲柳更新幼树定植生境WINDSD的均值与分布倾向度小于阔叶红松林下均匀布设样点,其余树种均大于阔叶红松林下均匀布设样点。9种主要乔木树种更新幼苗定植生境WINDSD的均值及分布倾向度由小至大的综合排序为紫椴lt;水曲柳lt;花楷槭lt;臭冷杉lt;红松lt;青楷槭lt;红皮云杉lt;色木槭lt;春榆;幼树定植生境WINDSD的均值及分布倾向度综合排序为水曲柳lt;春榆lt;色木槭lt;青楷槭lt;花楷槭lt;臭冷杉lt;紫椴lt;红松lt;红皮云杉。红松林下均匀布设样点与9种乔木树种更新幼苗、幼树定植生境的加权进界邻体散射荫蔽度的概率密度直方图见图2。
由图2可见,红松林下均匀布设样点的WINDSD的概率密度直方图呈钟形分布,表现出一定的负偏态(negative skewness)倾向,概率密度在0.5~0.6时最高;9种主要乔木树种群更新幼苗定植生境的WINDSD的概率密度直方图亦多呈钟形状分布,相较于阔叶红松林下均匀布设样点,各树种更新幼苗定植生境的WINDSD在0~0.4时概率密度均较低,该散射辐射强度区间对应于林分中的林隙区域;且概率密度的峰值多出现在0.5~0.7范围内。9种主要乔木树种中,春榆、色木槭、青楷槭、水曲柳4树种更新幼树定植生境的WINDSD的概率密度直方图中,0~0.4范围的累积概率密度明显高于对应树种更新幼苗的概率密度;相对而言,其余树种更新幼树与更新幼苗定植生境的WINDSD的概率密度直方图间的差异规律性较弱。
2.2 更新幼苗及幼树定植生境WINDSD的差异显著性分析
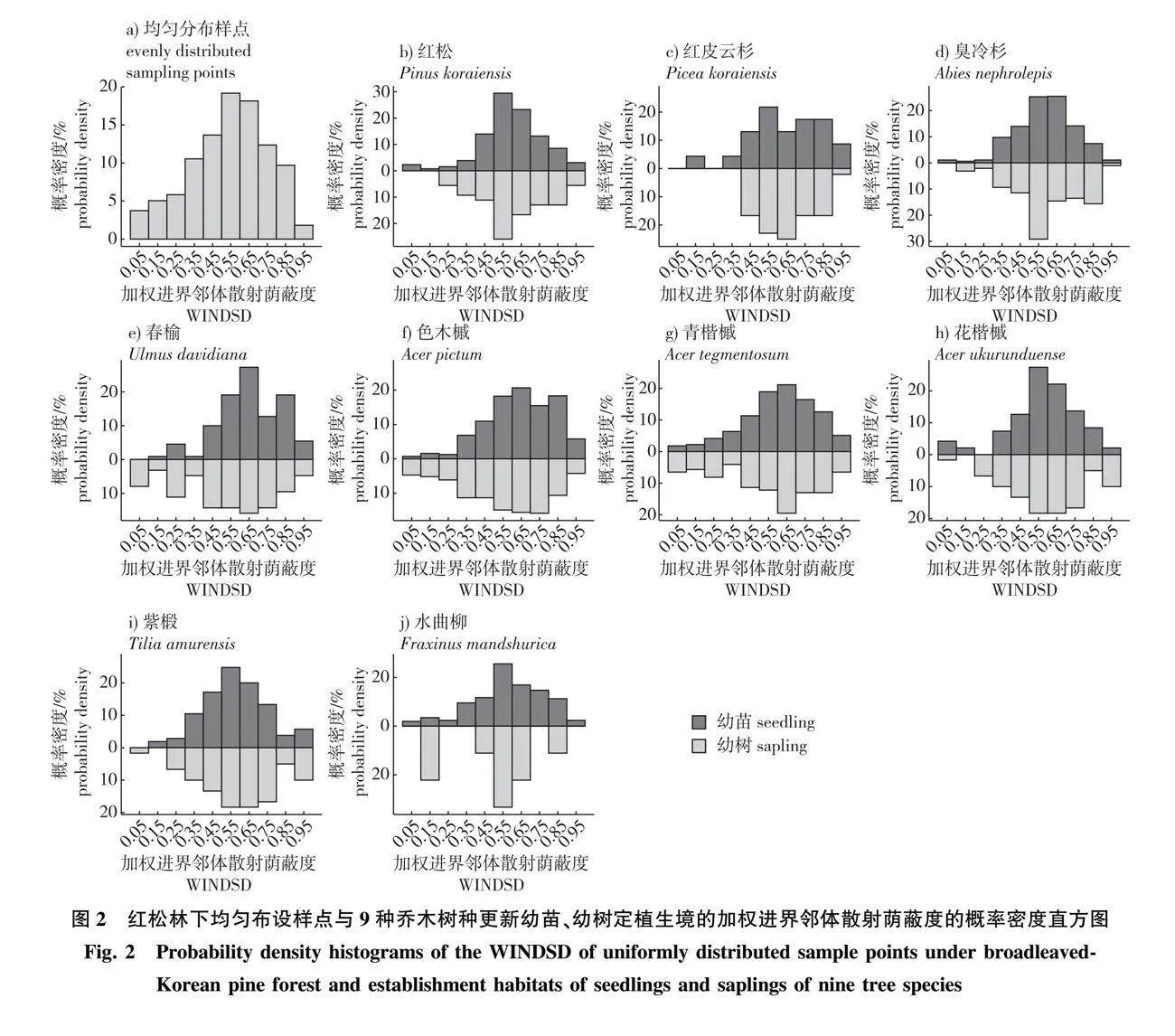
对9种乔木树种更新幼苗定植生境的WINDSD的分布倾向度与林下随机分布种群的WINDSD的分布倾向度间差异进行蒙特卡罗双尾检验,结果见图3。经检验,9种乔木树种中除花楷槭、紫椴更新幼苗定植生境WINDSD的分布倾向度与红松林下随机分布种群间的差异未达到显著水平,其余树种更新幼苗定植生境WINDSD的分布倾向度均显著区别于阔叶红松林下种群规模相同的随机分布种群;若进行单尾检验,则除紫椴外(P=0.052),其余树种幼苗定植生境的WINDSD均显著大于阔叶红松林下种群规模相同的随机分布种群(Plt;0.05)。
对9种乔木树种更新幼树定植生境的WINDSD的分布倾向度与林下随机分布种群的WINDSD的分布倾向度间差异进行蒙特卡罗双尾检验,结果见图4。经检验,在5%显著性水平下红松、红皮云杉与臭冷杉更新幼树定植生境的WINDSD的分布倾向度与阔叶红松林下同等规模随机分布种群存在显著差异,其余乔木树种无显著差异;如将双尾检验显著度转换为单尾,则紫椴幼树亦具有显著的分布于弱光生境中的倾向。
分别对阔叶红松林下均匀布设样点与红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、花楷槭、紫椴、水曲柳更新幼苗、幼树定植生境及相同树种更新幼苗、幼树定植生境间的WINDSD进行Mann-Whitney U检验,结果见表3。
结合表2中各树种幼苗、幼树定植生境WINDSD的均值与分布倾向度及表3中的差异显著性,可知在Mann-Whitney U检验的检验体系下,红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、水曲柳更新幼苗定植生境的WINDSD显著大于阔叶红松林下均匀布设样点,而花楷槭与紫椴更新幼苗定植生境的WINDSD与阔叶红松林下均匀布设样点无显著性差异;9种主要乔木树种中,红松、红皮云杉、臭冷杉更新幼树定植生境的WINDSD显著大于阔叶红松林下均匀布设样点,而其余树种均无显著性差异;相较于更新幼苗,春榆及色木槭更新幼树定植生境的WINDSD显著较小,而其余树种幼树幼苗定植生境间的WINDSD无显著差异。
3 讨 论
森林群落林下地被植物聚集形成的小斑块可称为小群落(minor community)或小群聚[29],其形成与林下环境异质性及植物幼龄个体的扩散限制具有一定关系,直观体现为群落的水平分化,即群落镶嵌现象[30]。基于本研究中4块样地的踏查结果,9种主要乔木树种幼龄植株(幼苗与幼树)总体呈聚集分布,且在林下WINDSD较大的位置密度较大,形成了以弱散射辐射区为聚集中心的空间镶嵌结构。经统计,研究所涉9种乔木树种的更新幼树、幼苗中,仅有春榆及水曲柳幼树定植生境的WINDSD的平均值与分布倾向度小于林下均匀布设样点。造成上述现象的原因可能包含以下几个方面:①本研究中调查的更新幼苗包括萌生苗与实生苗,而萌生苗多分布于与母树位置较近的弱光生境中,即繁殖体的扩散限制;②林下不同光强生境中乔木更新幼苗密度的差异与冠层植被及林下植被间的竞争网络关系[31]有关,红松林下林隙生境等光强较强的区域中灌、草层发育通常较为旺盛,而来自灌、草层的种间竞争会抑制乔木更新的生长,而冠层乔木树冠下的弱光生境抑制了灌木层与草本层的发育,进而在林下为乔木更新提供了生态位空缺;③阔叶红松林中林隙中心的土壤含水率高于林隙边缘[32],本研究中4块调查样地均位于中、下坡位,在雨季时样地内的林隙生境可能存在较长时间的水涝,林隙生境中较强的环境过滤减少了各类乔木树种的更新密度;④林隙生境中的强光环境会对针叶树种的越冬苗产生光氧化伤害[33],降低林隙中针叶树种幼苗的越冬率。
在分析9种乔木树种幼树、幼苗定植生境光照强度与林下均匀分布样点光照强度间的差异显著性时,基于分布倾向度的双尾蒙特卡罗检验与Mann-Whitney U检验的结果完全一致。两种检验均体现出红松、红皮云杉、臭冷杉、春榆、色木槭、青楷槭、水曲柳更新幼苗定植生境的WINDSD显著大于林分内平均水平,而花楷槭及紫椴更新幼苗定植生境的WINDSD则与林分内平均水平无显著性差异;同时,红松、红皮云杉、臭冷杉更新幼树定植生境的WINDSD显著高于林分内平均水平,其余树种与林分内平均水平无显著性差异。进一步将9种乔木树种更新幼苗、幼树定植生境的WINDSD合并,计算二者的株水平平均值,得出合并后乔木更新幼苗定植生境WINDSD的均值为0.608,更新幼树定植生境WINDSD的均值为0.569。这反映出阔叶红松林下强光生境对天然更新的株高生长总体上具有促进作用,与文献[19,34]中乔木幼苗在林隙内总体密度低于非林隙,而幼树总体密度在林隙中高于非林隙的观点具有一定的相似性。
9种主要乔木树种中,红松、红皮云杉、臭冷杉及紫椴的更新幼树定植生境WINDSD的均值与分布倾向度相较于对应树种的更新幼苗有所提升,其余树种相较于更新幼苗有所下降。阔叶红松林内,更新幼树WINDSD升高的树种红松、红皮云杉、臭冷杉及紫椴均具有生长至主林层的潜质,而更新幼树WINDSD降低的树种除水曲柳外均主要分布于亚林层及下木层中。由此可见,阔叶红松林内乔木树种的潜在最大树高与乔木树种幼龄阶段的光资源利用及光环境响应策略或存在协同效应。Zhang等[35]的研究显示,在C-S-R生长策略(C为竞争策略,S为忍耐策略,R为杂草策略)三角形中,红松更新幼树的C策略得分约为10%,S策略得分约为90%,R策略得分约为0%;色木槭、青楷槭、花楷槭3种槭树的C策略得分为35%~50%,S策略得分为20%~40%,R策略得分为35%~40%;春榆更新幼树的C策略得分约为25%,S策略得分约为40%,R策略得分约为40%;可见,相较于针叶树种红松,倾向分布于亚林层及下木层中的各类阔叶树种R策略得分均更高而S策略得分更低。紫椴幼龄植株对弱光环境的S策略在其叶性状中有所体现,紫椴的比叶面积具有较强的可塑性,金明月[36]及孙金伟等[37]的研究中均显示紫椴幼树的比叶面积显著高于紫椴成树(Plt;0.05),而提升比叶面积是植物应对弱光环境的典型形态学反应。红松更新的定植、生长需要一定的来源于上层阔叶树种的荫蔽[38],在面积较大的林隙内密度较低[39],这与本研究中红松幼树、幼苗定植生境WINDSD所表现出的特质一致;同时,红松更新的主要碳同化时期为早春及晚秋,利用上层阔叶树种完成展叶前及开始落叶后的光合积累,与上层阔叶树种的光资源竞争存在时间生态位分化[40]。与红松相同,云杉及常绿草本植物亦具有相同的适应性机制[41-42],即在早春及晚秋进行光合积累。上述因素或是红松、红皮云杉及臭冷杉等常绿针叶树种更新幼树定植生境WINDSD略高于更新幼苗的内在原因,因此,在林下针叶树植苗造林的过程中应避免将植苗位置选择在针叶树下。经Mann-Whitney U检验9种树种中春榆、色木槭更新幼苗与幼树定植生境间的WINDSD的差异达到显著水平(Plt;0.05),青楷槭更新幼苗与幼树定植生境间的WINDSD差异显著性仅次于色木槭与春榆,居于9种树种中的第3位。这表明林下辐射强度的增加对春榆、色木槭、青楷槭幼苗的生长具有明显的促进作用。
阔叶红松林中存在着以林隙动态为基础的中小尺度的局部群落演替系列,而林分内不同区域的局部群落组成的循环往复造就了阔叶红松林总体的流动镶嵌稳态(shifting mosaic steady state)[43],这种现象亦可称为阔叶红松林的群落内循环(cycle within community)[44]。梁星云[45]研究指出,在以阔叶红松林为顶极群落的演替系列中,比叶重(或比叶面积)是耐阴树种与不耐阴树种的最显著差异项。即使在阔叶红松林内林隙尺度的演替过程中乔木树种的耐阴性差异亦发挥着重要作用。基于本研究结果推断,相较于红松、红皮云杉、冷杉及紫椴等大乔木更新,春榆、色木槭、青楷槭更新的高生长对于强光环境的响应可能更为敏感。春榆、色木槭、青楷槭等树种的更新能在林下弱光生境中建立幼苗库,并在林隙形成初期优先占据林隙生境中的空缺生态位,进而为红松等树种的更新创造适宜的荫蔽条件;同时,相较于红松、紫椴等树种,色木槭、青楷槭、花楷槭、春榆等亚林层与下层木中的优势种生理寿命普遍较短,随着上层阔叶树种死亡,下层针叶树种逐渐进入主林层。由此可见,阔叶红松林内各树种在生态系统的稳定维持中均发挥着独特的生态功能,各树种通过种内、种间互作机制、空生态位分化形成了阔叶红松林特有的群落外貌与群落结构。因此,在次生林及人工针叶纯林的改造过程中应更加关注阔叶非目的树种的生态价值,以促进森林生态系统的恒续(continuous forest)经营。随着现阶段林业的属性由生产功能向生态、社会功能逐渐转变,在森林经营过程中,对传统杂木的处理应给予更多的思考。
参考文献(reference):
[1]王新程.环境科学大辞典[M].2版.北京:中国环境科学出版社,2008.WANG X C.Environmental science dictionary[M].2nd ed.Beijing:China Environmental Science Press,2008.
[2]黄一鑫,程艳霞.森林光环境对4种乔木幼树光合和光谱反射特性的影响[J].生态学报,2022,42(22):9121-9129.HUANG Y X,CHENG Y X.Photosynthetic characteristics and spectral reflectance characteristics of four natural tree saplings under forest light environment[J].Acta Ecol Sin,2022,42(22):9121-9129.DOI: 10.5846/stxb202105171289.
[3]刘子宣,贾存,秦志强,等.华北落叶松林下光环境对白扦幼树生长的影响[J].南京林业大学学报(自然科学版),2020,44(6):111-117.LIU Z X,JIA C,QIN Z Q,et al.Effects of light conditions on the growth of understory Picea meyeri sapling in Larix principis-rupprechtii forest[J].J Nanjing For Univ (Nat Sci Ed),2020,44(6):111-117.DOI: 10.3969/j.issn.1000-2006.201909032.
[4]WILKENS J F,WAGNER S.Empirical survival model for European beech (Fagus sylvatica L.) seedlings in response to interactive resource gradients and (a-) biotic conditions within an experimental canopy gap study[J].For Ecol Manag,2021,499:119627.DOI: 10.1016/j.foreco.2021.119627.
[5]XUE W X,GUO Q J,ZHU J,et al.Research on the effects of light intensity and seeding density on the seed germination and seedling growth of Liriodendron chinense[J].J Agric Chem Environ,2022,11(1):24-41.DOI: 10.4236/jacen.2022.111003.
[6]RITTER E,DALSGAARD L,EINHORN K S.Light,temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark[J].For Ecol Manag,2005,206(1/2/3):15-33.DOI: 10.1016/j.foreco.2004.08.011.
[7]COATES K D,BURTON P J.A gap-based approach for development of silvicultural systems to address ecosystem management objectives[J].For Ecol Manag,1997,99(3):337-354.DOI: 10.1016/S0378-1127(97)00113-8.
[8]ROUVINEN S,KOUKI J.Tree regeneration in artificial canopy gaps established for restoring natural structural variability in a Scots pine stand[J].Silva Fenn,2011,45(5):1079-1091.DOI: 10.14214/sf.88.
[9]SCHEIRE A. Functional traits of lianas: trait coordination and how they shape species light and nutrient niches[D]. Wageningen: Wageningen University, 2019.
[10]LONG W X,ZANG R G,SCHAMP B S,et al.Within and among-species variation in specific leaf area drive community assembly in a tropical cloud forest[J].Oecologia,2011,167(4):1103-1113.DOI: 10.1007/s00442-011-2050-9.
[11]LAURANS M,MARTIN O,NICOLINI E,et al.Functional traits and their plasticity predict tropical trees regeneration niche even among species with intermediate light requirements[J].J Ecol,2012,100(6):1440-1452.DOI: 10.1111/j.1365-2745.2012.02007.x.
[12]GROGAN J,LANDIS R M,ASHTON M S,et al.Growth response by big-leaf mahogany (Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Pará,Brazil[J].For Ecol Manag,2005,204(2/3):399-412.DOI: 10.1016/j.foreco.2004.09.013.
[13]MUHAMED H,TOUZARD B,LE BAGOUSSE-PINGUET Y, et al.The role of biotic interactions for the early establishment of oak seedlings in coastal dune forest communities[J].For Ecol Manag,2013,297:67-74.DOI: 10.1016/j.foreco.2013.02.023.
[14]刘从,田甜,李珊,等.中国木本植物幼苗生长对光照强度的响应[J].生态学报,2018,38(2):518-527.LIU C,TIAN T,LI S,et al.Growth response of Chinese woody plant seedlings to different light intensities[J].Acta Ecol Sin,2018,38(2):518-527.DOI: 10.5846/stxb201611012221.
[15]徐化成.中国红松天然林[M].北京:中国林业出版社,2001.XU H C.Natural forests of Pinus koraiensis in China[M].Beijing:China Forestry Publishing House,2001.
[16]吴刚.长白山红松阔叶混交林林冠空隙树种更新动态规律的研究[J].应用生态学报,1998,9(5):2-5.WU G.Regeneration dynamics of tree species in gaps of Korean pine broad leaved mixed forest in Changbai Mountains[J].Chin J Appl Ecol,1998,9(5):2-5.DOI: 10.1088/0256-307X/15/12/025.
[17]国庆喜.阔叶红松林林冠斑块特征及其与林木更新的关系[J].应用生态学报,2002,13(12):1541-1543.GUO Q X. Characteristics of canopy patches related to natural regeneration of broad-leaved Korean pine forest[J].Chin J Appl Ecol,2002,13(12):1541-1543.
[18]刘妍妍,金光泽,李凤日.典型阔叶红松林林隙对幼苗建立的影响[J].科学通报,2014,59(24):2396-2406.LIU Y Y,JIN G Z,LI F R.Influence of forest gaps on seedling establishment in a mixed broadleaved-Korean pine(Pinus koraiensis) forest in Xiao Hinggan Mountains[J].Chin Sci Bull,2014,59(24):2396-2406.
[19]JIN G Z, TIAN Y, ZHAO F, et al. The pattern of natural regeneration by canopy gap size in the mixed broadleaved-Korean pine forest of Xiaoxing’an Mountains, China[J]. J Korean For Soc, 2007, 96: 227-234.
[20]JIN G Z,LIU Y Y,LIU S,et al.Effect of gaps on species diversity in the naturally regenerated mixed broadleaved-Korean pine forest of the Xiaoxing’an Mountains,China[J].J Ecol Environ,2007,30(4):325-330.DOI: 10.5141/jefb.2007.30.4.325.
[21]董莉莉,张慧东,毛沂新,等.间伐对红松Pinus koraiensis针阔混交林冠层结构及林下植被的影响[J].沈阳农业大学学报,2017,48(2):159-165.DONG L L,ZHANG H D,MAO Y X,et al.Influence of thinning on canopy structure and understory vegetation of coniferous-broad leaved Pinus koraiensis mixed forest[J].J Shenyang Agric Univ,2017,48(2):159-165.
[22]FRAZER G W, CANHAM C D, LERTZMAN K P. Gap light analyzer (GLA) Version 2.0: users manual and program documentation[R]. Vancouver: Simon Fraser University and the Institute of Ecosystem Studies, 1999.
[23]刘赫男,张洪玲,朱红蕊.1961—2010年黑龙江省太阳能资源特征分析与评估[J].气象与环境学报,2013,29(4):89-93.LIU H N,ZHANG H L,ZHU H R.Analysis and assessment of solar energy resource in Heilongjiang Province from 1961 to 2010[J].J Meteor Environ,2013,29(4):89-93.DOI: 10.3969/j.issn.1673-503X.2013.04.014.
[24]杜昕,董雪,谷会岩,等.基于易测林木因子的林下太阳辐射模拟(Ⅰ)——林下散射辐射模拟[J/OL].南京林业大学学报(自然科学版),[2024-06-03].https://link.cnki.net/urlid/32.1161.s.20240603.1546.002. DU X, DONG X, GU H Y,et al.Simulation of understory solar radiation based on easily measurable tree factors (Ⅰ): understory diffuse radiation simulation[J/OL].J Nanjing For Univ (Nat Sci Ed),[2024-06-03].https://link.cnki.net/urlid/32.1161.s.20240603.1546.002.
[25]罗耀华,陈庆诚,张鹏云.兴隆山阴暗针叶林空间格局及其利用光能的对策[J].生态学报,1984,4(1):10-20.LUO Y H,CHEN Q C,ZHANG P Y.The spatial pattern of coniferous forest in Xinglongshan Mountain and its strategies in using sun light energy[J].Acta Ecol Sin,1984,4(1):10-20.
[26]魏殿生.全国森林培育技术标准汇编:造林经营卷[M].北京:中国标准出版社,2003.
[27]惠刚盈.结构化森林经营原理[M].北京:中国林业出版社,2016.HUI G Y.Principles of structure-based forest management[M].Beijing:China Forestry Publishing House,2016.
[28]杜昕,董雪,谷会岩,等.基于分层 Voronoi 图的阔叶红松林叶面积指数的垂直与短程水平空间分布研究[J/OL].南京林业大学学报(自然科学版),[2023-11-17].https://link.cnki.net/urlid/32.1161.S.20231117.0856.002. DU X, DONG X, GU H Y,et al.A study of the vertical and short-range horizontal spatial distribution of leaf area index in broadleaved-Korean pine forest based on stratified Voronoi diagrams[J/OL].J Nanjing For Univ (Nat Sci Ed),[2023-11-17]. https://link.cnki.net/urlid/32.1161.S.20231117.0856.002.
[29]王献溥.小群聚(小群落)的概念及其在研究针叶、落叶阔叶混交林结构时的应用[J].植物生态学与地植物学丛刊,1963(S1):51-68.WANG X P.The concept of small community and its application in studying the structure of coniferous and deciduous broad-leaved mixed forest[J].Chin J Plant Ecol,1963(S1):51-68.
[30]孙龙,国庆喜.生态学基础[M].北京:中国建材工业出版社,2013.SUN L,GUO Q X.Ecological basis[M].Beijing:China Building Material Industry Publishing House,2013.
[31]张炜平,王根轩.植物邻体间的正相互作用[J].生态学报,2010,30(19):5371-5380.ZHANG W P,WANG G X.Positive interactions in plant communities[J].Acta Ecol Sin,2010,30(19):5371-5380.
[32]王丽霞.阔叶红松混交林林隙大小、土壤水分以及光照对植物的影响[D].哈尔滨:东北林业大学,2013.WANG L X.Effects of gap size,soil moisture and light on plants in broad-leaved Korean pine mixed forest[D].Harbin:Northeast Forestry University,2013.
[33]白淑菊,陶大立,靳月华.长白山常绿针叶树越冬期间光合能力的抑制[J].应用生态学报,1995,6(2):138-142.BAI S J,TAO D L,JIN Y H.Inhibition of photosynthetic capacity of evergreen conifers in Changbai Mountain during overwintering[J].Chin J Appl Ecol,1995,6(2):139-142.
[34]田悦颖.小兴安岭阔叶红松林林隙特征及其更新研究[D].哈尔滨:东北林业大学,2007.TIAN Y Y.Gap characteristics and regeneration of broad-leaved Korean pine forest in Xiaoxing’an Mountains[D].Harbin:Northeast Forestry University,2007.
[35]ZHANG X J,WANG S L.Joint control of plant ecological strategy by climate,regeneration mode,and ontogeny in northeastern Chinese forests[J].Ecol Evol,2021,11(11):6703-6715.DOI: 10.1002/ece3.7522.
[36]金明月.季节及树龄对东北阔叶树种叶性状及其权衡的影响[D].哈尔滨:东北林业大学,2019.JIN M Y.Effects of season and tree age on leaf characteristics and trade-offs of broad-leaved trees in northeast China[D].Harbin:Northeast Forestry University,2019.DOI: 10.27009/d.cnki.gdblu.2019.000348.
[37]孙金伟,姚付启,张振华.红松和紫椴叶片暗呼吸及其光抑制性在幼、成树间的差异[J].应用生态学报,2019,30(5):1463-1468.SUN J W,YAO F Q,ZHANG Z H.Differences of leaf dark respiration and light inhibition between saplings and mature trees of Pinus koraiensis and Tilia amurensis[J].Chin J Appl Ecol,2019,30(5):1463-1468.DOI: 10.13287/j.1001-9332.201905.008.
[38]王树力,武敬辉,史永纯.红松种群天然更新及幼年生长与林分结构关系的研究[J].吉林林学院学报,1998(1):8-12.WANG S L,WU J H,SHI Y C.Study on the relationship between natural regeneration,juvenile growth and stand structure of Korean pine population[J].J Jilin For Univ,1998(1):8-12.
[39]贺丹妮,杨华,温静,等.长白山云冷杉针阔混交林不同林隙下幼苗幼树密度及空间分布[J].应用生态学报,2020,31(6):1916-1922.HE D N,YANG H,WEN J,et al.Density and spatial distribution of seedlings and saplings in different gap sizes of a spruce-fir mixed stand in Changbai Mountains,China[J].Chin J Appl Ecol,2020,31(6):1916-1922.DOI: 10.13287/j.1001-9332.202006.004.
[40]周光.林下红松生存策略与季节光环境驱动机制[D].北京:北京林业大学,2019.ZHOU G.Survival strategy of Korean pine under forest and driving mechanism of seasonal light environment[D].Beijing:Beijing Forestry University,2019.DOI: 10.26949/d.cnki.gblyu.2019.000156.
[41]王乾.川西亚高山森林生态系统辐射传输研究[D].成都:中国科学院研究生院(成都生物研究所),2006.WANG Q.Study on radiation transmission of alpine forest ecosystem in western Sichuan[D].Chengdu:Chengdu Institute of Biology,Chinese Academy of Sciences,2006.
[42]李秋果,卞英捷,梁照,等.匍匐翦股颖品种(系)的坪用性状与耐热性评价[J].南京农业大学学报,2023,46(6):1096-1106.LI Q G,BIAN Y J,LIANG Z,et al.Evaluation of turf characteristics and heat tolerance of creeping bentgrass cultivars(strains)[J].J Nanjing Agric Univ,2023,46(6):1096-1106.DOI: 10.7685/jnau.202210010.
[43]ZENNER E K,PECK J E,HOBI M L.Development phase convergence across scale in a primeval European beech (Fagus sylvatica L.) forest[J].For Ecol Manag,2020,460:117889.DOI: 10.1016/j.foreco.2020.117889.
[44]王伯荪.植被的镶嵌体系[J].生态科学,1998,17(2):3-9.WANG B S.Mosaic system of vegetation[J].Ecol Sci,1998,17(2):3-9.
[45]梁星云.长白山阔叶红松林演替系列主要树种叶片功能性状与化学计量学研究[D].北京:中国林业科学研究院,2017. LIANG X Y.Study on leaf functional characters and chemometrics of main tree species in succession series of broad-leaved Korean pine forest in Changbai Mountain[D].Beijing:Chinese Academy of Forestry,2017.
(责任编辑 李燕文)
猜你喜欢
数字技术与应用(2017年8期)2017-10-30 16:28:48
绿色科技(2017年17期)2017-09-30 09:29:50
现代农业科技(2017年10期)2017-07-12 14:24:02
现代农业科技(2017年3期)2017-03-28 15:27:59
中学生物学(2017年2期)2017-03-20 07:30:12
现代农业科技(2017年1期)2017-03-06 12:38:31
农业与技术(2016年20期)2016-12-08 21:57:43
安徽农学通报(2016年16期)2016-10-24 19:43:15
湖南师范大学学报·自然科学版(2016年3期)2016-06-25 06:22:53
热带农业科学(2015年7期)2015-08-11 04:32:17
