
低温胁迫对茶树生长发育和叶绿素荧光参数的影响
2024-12-05 00:00:00肖黄巧石玉玲赵龙许妍揭雨成
江苏农业科学 2024年23期

摘要:为探究低温胁迫对茶树叶片光系统Ⅱ(PSⅡ)荧光参数及SPAD值的影响,分析在不同低温条件下茶树叶片PSⅡ荧光参数及次生代谢物的变化规律。以黄金茶1号为试材,在智能人工气候箱中进行试验,共设置1、3、5 ℃ 3个低温,以25 ℃为对照,处理时间为3、5、7、9 d。研究表明,在低温胁迫处理下茶叶的农艺性状与CK相比均存在不同幅度的下降;次生代谢物质、抗逆性指标和叶绿素荧光参数等指标较CK均存在不同幅度的上升或下降,其中茶叶的TP含量较CK总体下降了0.11%~31.97%;游离氨基酸含量较CK总体增加了5.24%~104.35%;MDA含量较CK总体增加了23.56%~126.68%;SS含量较CK总体增加了22.36%~236.13%;PhiNPQ、SPAD值、Phi2、Fv/Fm较CK总体分别下降了9.94%~41.10%、14.29%~34.85%、10.25%~51.24%、11.39%~48.53%;而PhiNO、NPQ较CK总体分别增加了29.28%~125.12%、22.13%~70.99%。综上所述,茶树在低温胁迫下叶长、叶宽、TP含量、PhiNPQ、SPAD值、Phi2、Fv/Fm总体随着温度的降低和胁迫时间的延长而下降;MDA含量、PhiNO、NPQ随着温度的降低和胁迫时间的延长总体呈上升趋势;游离氨基酸含量总体随温度降低呈先升后降的趋势,但随胁迫时间的延长变化趋势不同;SS含量随温度的降低逐渐上升,随胁迫时间的延长呈先升后降的趋势。由此可知,在低温胁迫下不仅会影响茶树生长发育,还会致使其叶片PSⅡ活性受损,光合效率下降,出现光抑制现象,影响茶树的生长发育,不利于其品质形成。
关键词:低温胁迫;茶树;生长发育;叶片;荧光参数;农艺性状;抗逆性指标
中图分类号:S571.101" 文献标志码:A
文章编号:1002-1302(2024)23-0147-09
肖黄巧,石玉玲,赵" 龙,等. 低温胁迫对茶树生长发育和叶绿素荧光参数的影响[J]. 江苏农业科学,2024,52(23):147-155.
doi:10.15889/j.issn.1002-1302.2024.23.021
收稿日期:2023-12-23
基金项目:国家重点研发计划(编号:2019YFD1002205-3)。
作者简介:肖黄巧(1999—),女,湖南洞口人,硕士研究生,主要从事逆境生物学研究。E-mail:1036244520@qq.com。
通信作者:揭雨成,博士,教授,主要从事作物种质创新与逆境生物学研究。E-mail:ibfcjyc@vip.sina.com。
茶树原产于我国西南地区,是我国重要的经济作物之一[1]。其中黄金茶是湘西北武陵山区保靖县的一个古老珍稀的地方品种,具有发芽早、产量高、品质优、抗性强等特性,享有“一两黄金一两茶”的美誉[2-3]。因为茶树具有喜暖怕寒的特性,所以低温是茶叶在生产过程中极易遭遇的非生物胁迫之一[4]。茶树低温胁迫主要有越冬期胁迫和芽萌发以后的“倒春寒”胁迫[5]。其中“倒春寒”胁迫是影响茶叶生产的气象灾害之一,它会造成茶树芽叶生长受阻,导致幼嫩新梢受冷变褐、焦枯坏死,不止会影响茶叶品质,还会对茶叶产量造成重大损失[6-8]。因此,在茶树生产中,了解低温胁迫对茶叶的影响机制,从而采取措施有效缓解低温危害,对提高茶叶品质和产量至关重要。已有研究表明,植物体内干物质的积累和产量90%左右都是叶片进行光合作用形成的,叶绿素荧光参数的变化不仅可以反映植物叶片光合作用的原初反应,还可以反映植物对环境的响应机制,可作为逆境胁迫研究的重要指标,对研究逆境与植物光合作用之间的关系具有重要意义[9-11]。叶绿素荧光与光合作用密切相关,叶绿素荧光动力学可以反映各种环境因子对光合作用的影响[12]。李庆会等的研究表明,茶叶在低温胁迫下光合能力减弱,直接损伤PSⅡ反应中心,致使PSⅡ反应中心积累大量过剩的激发能[13]。黄丽芳等以小粒种咖啡幼苗为研究对象,发现低温胁迫不仅抑制了幼苗叶片光合作用,还削弱了叶绿素荧光特性[14]。已有研究表明,叶片光合机构功能异常是植株在胁迫条件下其叶片组织受到损伤、大量积累丙二醛(MDA)导致的[15]。目前,低温胁迫对叶绿素荧光的影响在小麦[16]、木薯[17]、苜蓿[18]等植物上都有研究。对茶叶在低温胁迫方面的研究相对较少。本试验以黄金1号为试材,从叶绿素荧光参数入手,设置不同胁迫温度和时长,模拟低温胁迫试验,从生长发育、品质和荧光参数的角度,探讨低温胁迫对茶树叶片生长发育及叶绿素荧光参数的影响,分析不同低温条件下茶树叶片叶绿素荧光参数的变化规律,为茶树逆境栽培提供理论依据。
1" 材料与方法
1.1" 供试材料
以湘西土家族苗族自治州茶树科学研究所提供的黄金茶1号1.5年生左右扦插苗为研究试材。
1.2" 试验设计
于2022年10月将茶树从湘西土家族苗族自治州茶叶科学研究所运回湖南农业大学中学部,将茶树统一移栽于口径18 cm、高18.5 cm的塑料盆中,每盆1株,置于大棚内进行常规水肥、病虫害管理。于2023年4月选取长势良好、一致的茶树进行低温处理。设置低温胁迫温度为1、3、5 ℃,以25 ℃为对照(CK),处理时间为 3、5、7、9 d,每个处理供试茶树为4盆,共64盆。低温试验于4月10日在湖南省农业环境生态研究所的RXZ型智能人工气候箱(宁波江南仪器厂制造)进行,光周期设置昼夜 16 h/8 h,相对湿度为75%。待各处理结束后进行农艺性状等指标的测定,并取倒1~4叶置于 -80 ℃ 冰箱,用于后续测定茶多酚(TP)、游离氨基酸、丙二醛(MDA)、可溶性糖(SS)含量。
1.3" 测定指标及方法
1.3.1" 农艺性状测定" 于处理前及处理后利用游标卡尺对试验材料倒3叶进行长、宽的测定。
1.3.2" 叶片光系统Ⅱ(PSⅡ)荧光参数及SPAD值测定" 于处理前及处理后利用植物多功能仪PhotosynQ对试验材料倒3叶进行测定,其中每棵茶树取相同叶位功能叶片上的2个点进行测量,测定指标为PSⅡ实际光合速率(Phi2)、PSⅡ调节性能量耗散(PhiNPQ)、PSⅡ非调节性能量耗散(PhiNO)、相对叶绿素含量(SPAD值)、非光化学猝灭系数(NPQ)、PSⅡ原初光能转化效率(Fv/Fm)。
1.3.3" 茶多酚、游离氨基酸、丙二醛、可溶性糖含量测定" 均按试剂盒说明书进行测定。
1.4" 数据处理
采用Excel 2019对数据进行整理及绘图,利用SPSS 26.0软件对数据进行显著性检验(α=0.05)和相关性分析(Person)。
2" 结果与分析
2.1" 低温胁迫对茶叶农艺性状的影响
由表1可知,就叶长、叶宽处理前和处理后分析结果可知,CK处理后的叶长和叶宽相比处理前有所提升,分别增加了3.67%~12.89%、2.41%~21.09%,低温胁迫处理后除了5 ℃处理3 d的叶宽较处理前增加了1.80%外,其余低温胁迫后较处理前均呈下降趋势,5 ℃处理后的叶长和叶宽分别较处理前减少了0.99%~7.67%、1.53%~4.38%;3 ℃ 处理后的叶长和叶宽分别较处理前减少了2.71%~9.89%、0.64%~5.85%;1 ℃处理后的叶长和叶宽分别较处理前减少了4.12%~11.04%、2.80%~8.60%。就低温处理后的叶长、叶宽数据分析可知,随着处理温度的降低,胁迫时间为3 d时,5、3、1 ℃处理的叶长与CK相比,存在显著差异,而叶宽无显著差异;胁迫时间为5 d时,各低温处理后的叶长与CK相比无显著差异,而1 ℃的叶宽与CK存在显著差异,5 ℃和3 ℃与CK无显著差异;胁迫时间为7 d时,3个低温处理后的叶长与CK相比
茶叶在相同天数水平下不同温度水平间差异显著(Plt;0.05)。
均存在显著差异,叶宽除1 ℃与CK存在显著差异外,其余2个低温处理的叶宽与CK均无显著差异;胁迫时间为9 d时,5 ℃的叶长与CK存在显著差异,3 ℃和1 ℃与CK相比均无显著差异,而叶宽与CK相比无显著差异。
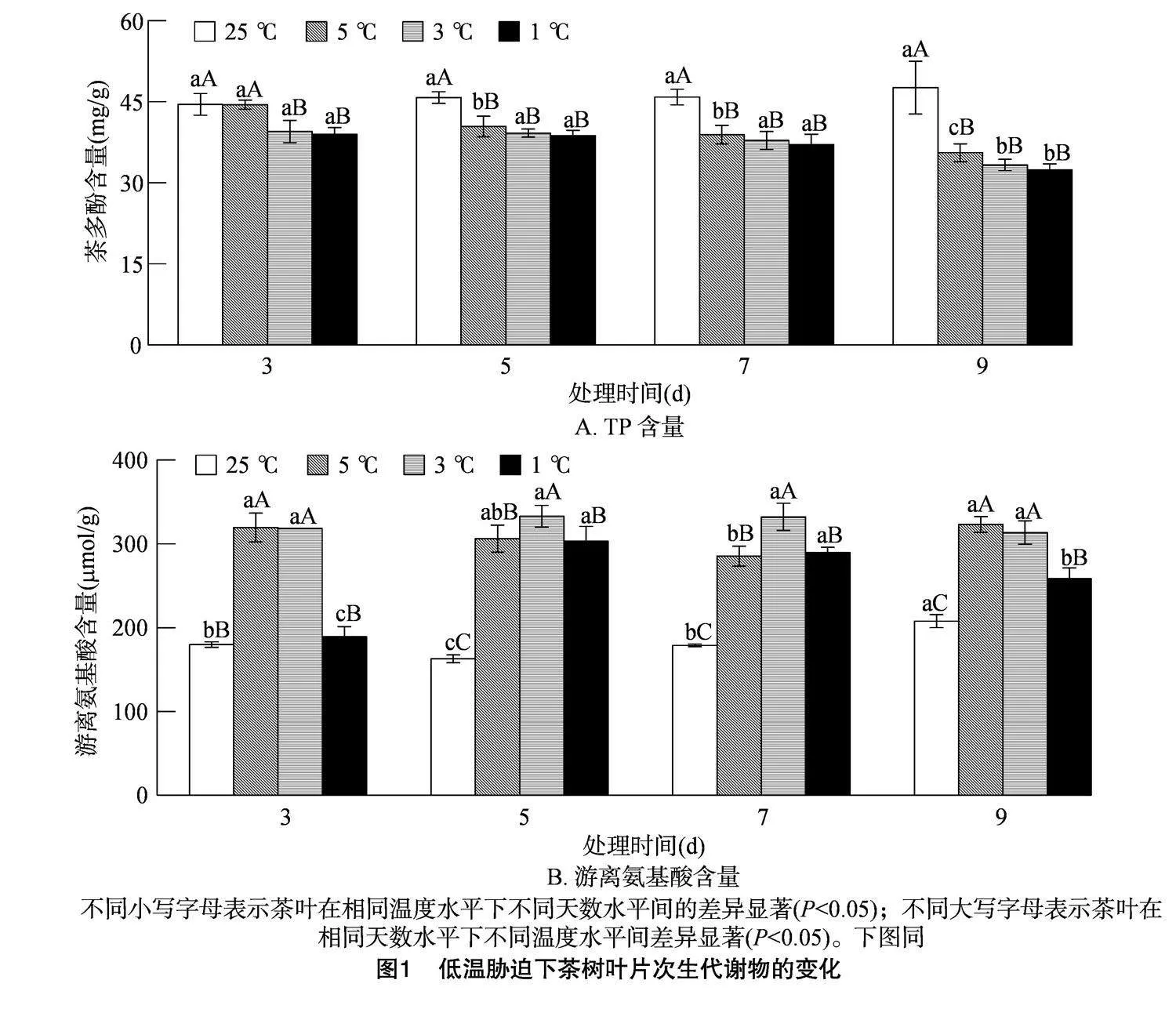
2.2" 低温胁迫对茶叶次生代谢物的影响
由图1-A可知,随着胁迫时间的延长,CK的茶多酚(TP)含量不断增加,但不存在显著差异,而其余3个温度下的TP含量均呈不断下降的趋势,其中5 ℃处理3 d时TP含量最高,较CK处理3 d时减少了0.11%;1 ℃处理9 d时TP含量最低,较CK处理9 d时减少了31.97%。随着处理温度的降低,各处理下的TP含量呈下降趋势,其中胁迫时间为 3 d 时,5 ℃ 的TP含量与CK无显著差异,而胁迫时间为5、7、9 d时TP的含量与CK相比均存在显著差异。表明除CK外,其余3个处理在不同温度梯度的胁迫下,黄金茶1号的鲜叶TP含量总体随温度的降低和胁迫时间的延长而逐渐降低。
由图1-B可知,随胁迫时间延长,CK和 5 ℃ 的游离氨基酸含量呈先下降后上升的趋势,而其余2个温度下的游离氨基酸含量均呈先上升后下降的趋势,其中5 ℃处理的最高峰出现在胁迫9 d时,较CK处理9 d时增加了55.53%;3、1 ℃处理的最高峰均出现在胁迫5 d时,较CK处理5 d时分别增加了104.35%、86.02%。随着处理温度的降低,各处理下的游离氨基酸含量呈先上升后下降的趋势,胁迫时间为3、9 d时,5、3 ℃之间无显著差异,而与CK和 1 ℃ 之间存在显著差异;胁迫时间为5、7 d 时,3 ℃ 与CK、5 ℃、1 ℃均存在显著差异。表明黄金茶1号的游离氨基酸含量随处理温度的降低总体呈先上升后下降的趋势,但随胁迫时间延长变化不同。
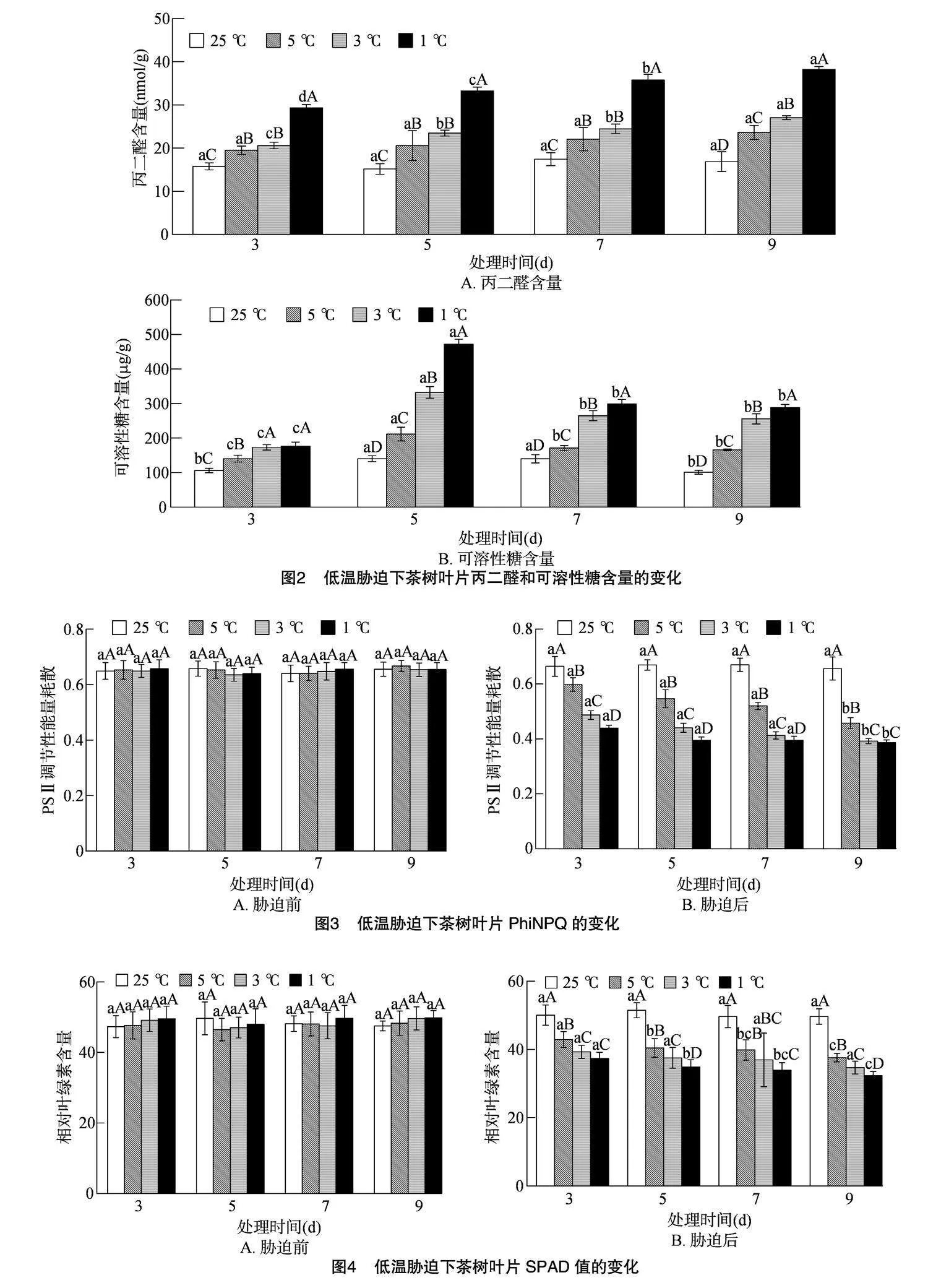
2.3" 低温胁迫对茶树叶片丙二醛和可溶性糖含量的影响
丙二醛含量表示植物在逆境条件下受损害的情况,丙二醛含量越高,表示植物在逆境下受伤害的程度越高。由图2-A可知,随着胁迫时间的延长,5、3、1 ℃的丙二醛(MDA)含量逐渐增加,其中5、3、1 ℃处理的最高峰均出现在胁迫9 d时,较CK处理9 d时分别增加了40.12%、60.41%、126.68%。随着处理温度的降低,各处理的MDA含量均呈上升趋势,胁迫时间为3、5、7 d时,5 ℃和 3 ℃ 之间的MDA含量无显著差异,但与CK和1 ℃之间均存在显著性差异;而胁迫时间为 9 d 时,5、3、1 ℃与CK之间均存在显著性差异。研究表明黄金茶1号的MDA含量总体随温度的降低和胁迫时间的延长而逐渐上升。
由图2-B可知, 随胁迫时间延长,各处理可溶性糖(SS)含量呈先上升后下降的趋势,其中5、3、1 ℃ 处理的最高峰均出现在胁迫5 d时,较CK处理5 d时分别增加了50.76%、136.62%、236.13%。随着处理温度的降低,各处理的SS含量均呈上升趋势,胁迫时间为3 d时,3 ℃和1 ℃处理间无显著差异,但与5 ℃和CK之间存在显著性差异;5、7、9 d时,5、3、1 ℃与CK之间均存在显著性差异。研究表明黄金茶1号的SS含量总体随温度的降低呈上升趋势,而随胁迫时间的延长呈先上升后下降的趋势。
2.4" 低温胁迫对茶树叶片叶绿素荧光参数的影响
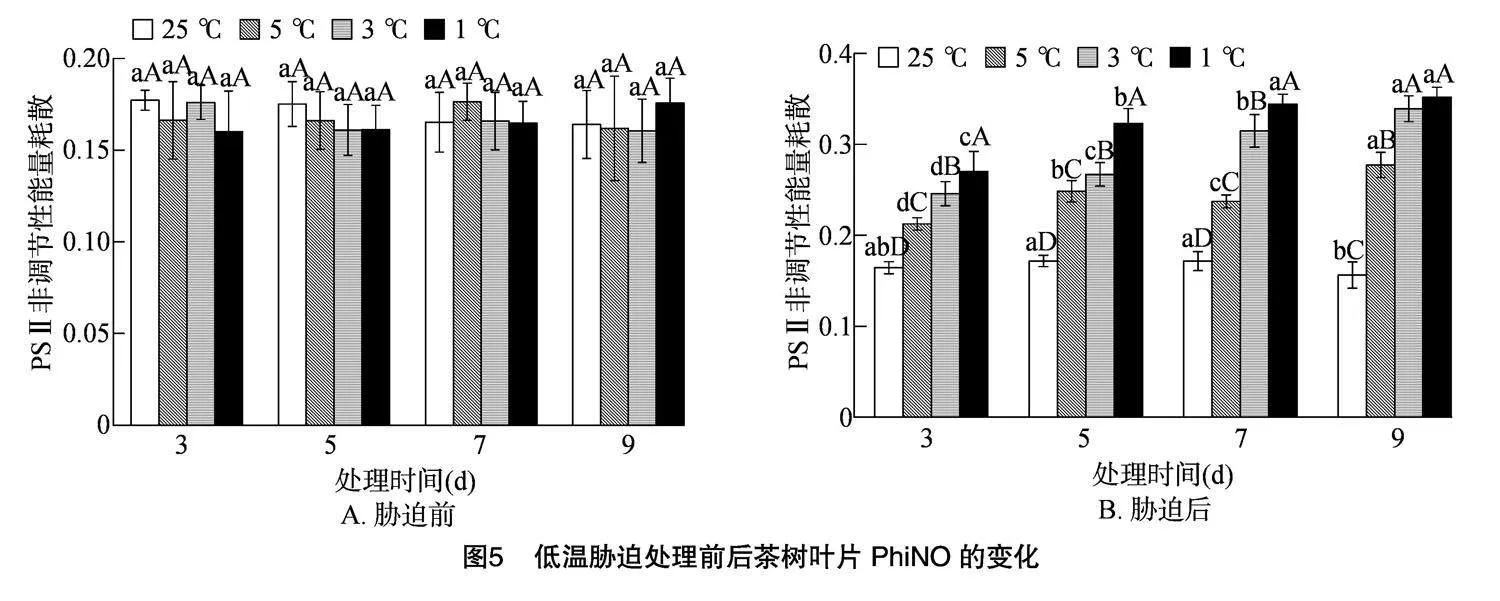
2.4.1" 低温胁迫对茶树叶片PhiNPQ的影响
PhiNPQ是光保护的重要指标之一,是植物利用过剩的能量来避免伤害的一种途径。由图3-A可知,在低温胁迫处理前各处理间的PhiNPQ无显著差异,PhiNPQ在0.636~0.667之间。由图3-B可知,随着胁迫时间的延长,5、3、1 ℃的PhiNPQ逐渐下降。随着处理温度的降低,各处理的PhiNPQ均呈下降趋势,各胁迫时间下的3个低温处理均与CK存在显著差异,在胁迫时间为3 d时,5、3、1 ℃较CK分别减少9.94%、26.65%、33.84%;在胁迫时间为 5 d 时,5、3、1 ℃较CK分别减少18.36%、34.17%、40.97%;在胁迫时间为7 d时,5、3、1 ℃较CK分别减少22.37%、38.45%、41.10%;在胁迫时间为9 d时,5、3、1 ℃较CK分别减少30.27%、40.26%、41.02%。研究表明黄金茶1号的PhiNPQ总体随温度的降低和胁迫时间的延长呈下降趋势。
2.4.2" 低温胁迫对茶树叶片SPAD值的影响
SPAD值代表叶片中叶绿素的相对含量,可以反映作物在某一程度上的生长情况。由图4可知,在低温胁迫处理前各处理间的SPAD值在46.48~49.84之间,而低温胁迫处理后各处理间的SPAD值均低于未处理前,其中CK较未处理前增加了3.00%~5.74%,而低温胁迫处理后5、3、1 ℃较未处理前分别减少了10.08%~22.23%、20.11%~30.16%、24.66%~35.14%。随着胁迫时间的延长,5、3、1 ℃ 的SPAD值逐渐下降。随着处理温度的降低,各处理的SPAD值均呈下降趋势, 在4个胁迫时间下的3个低温处理均与CK存在显著差异,在胁迫时间为3 d时,5、3、1 ℃较CK分别减少14.29%、21.50%、25.36%;在胁迫时间为5 d时,5、3、1 ℃较CK分别减少21.50%、27.13%、32.31%; 在胁迫时
间为7 d时,5、3、1 ℃较CK分别减少19.74%、25.59%、31.68%;在胁迫时间为9 d时,5、3、1 ℃较CK分别减少24.28%、30.14%、34.85%。研究表明黄金茶1号的SPAD值总体随温度的降低和胁迫时间的延长呈下降趋势。
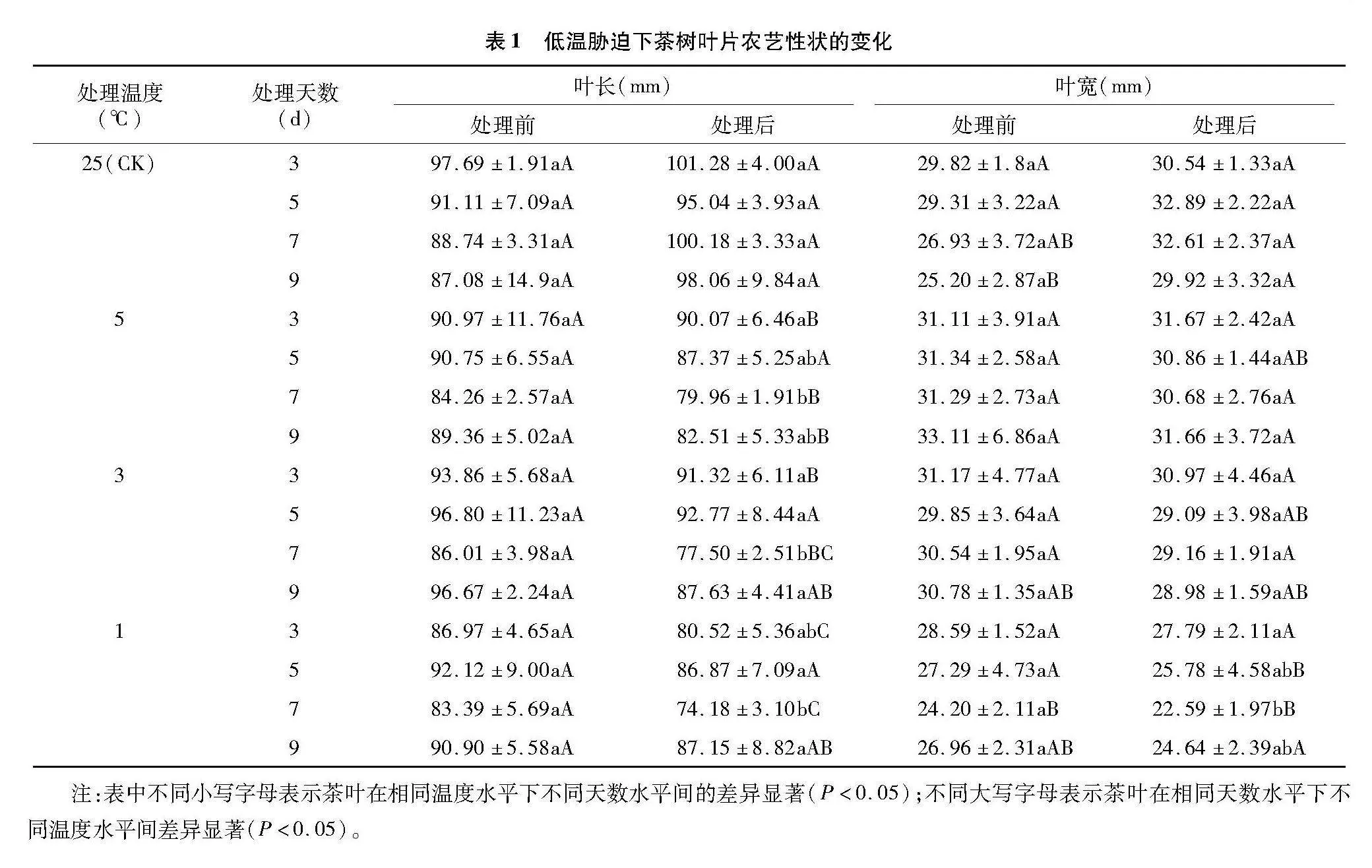
2.4.3" 低温胁迫对茶树叶片PhiNO的影响
由图5可知,低温胁迫处理前各处理间的PhiNO无显著差异,而低温胁迫处理后5、3、1 ℃的PhiNO与低温胁迫处理前相比分别增加了27.82%~71.35%、39.63%~111.29%、68.70%~108.64%。随着胁迫时间的延长,3、1 ℃的PhiNO呈不断上升趋势,而5 ℃的PhiNO则呈先升后降再升的趋势,5 ℃处理 9 d 时PhiNO最高,为0.277。随着处理温度的降低,各处理的PhiNO均呈上升趋势,在4个胁迫时间下的3个低温处理均与CK存在显著差异,在胁迫时间为3 d时,5、3、1 ℃较CK分别增加了29.28%、49.51%、64.33%;在胁迫时间为5 d时,5、3、1 ℃较CK分别增加了44.61%、55.46%、88.14%;在胁迫时间为7 d时,5、3、1 ℃较CK分别增加了38.24%、83.32%、100.44%;在胁迫时间为9 d时,5、3、1 ℃较CK分别增加了77.52%、117.04%、125.12%。研究表明黄金茶1号的PhiNO总体随温度的降低和胁迫时间的延长呈上升趋势。
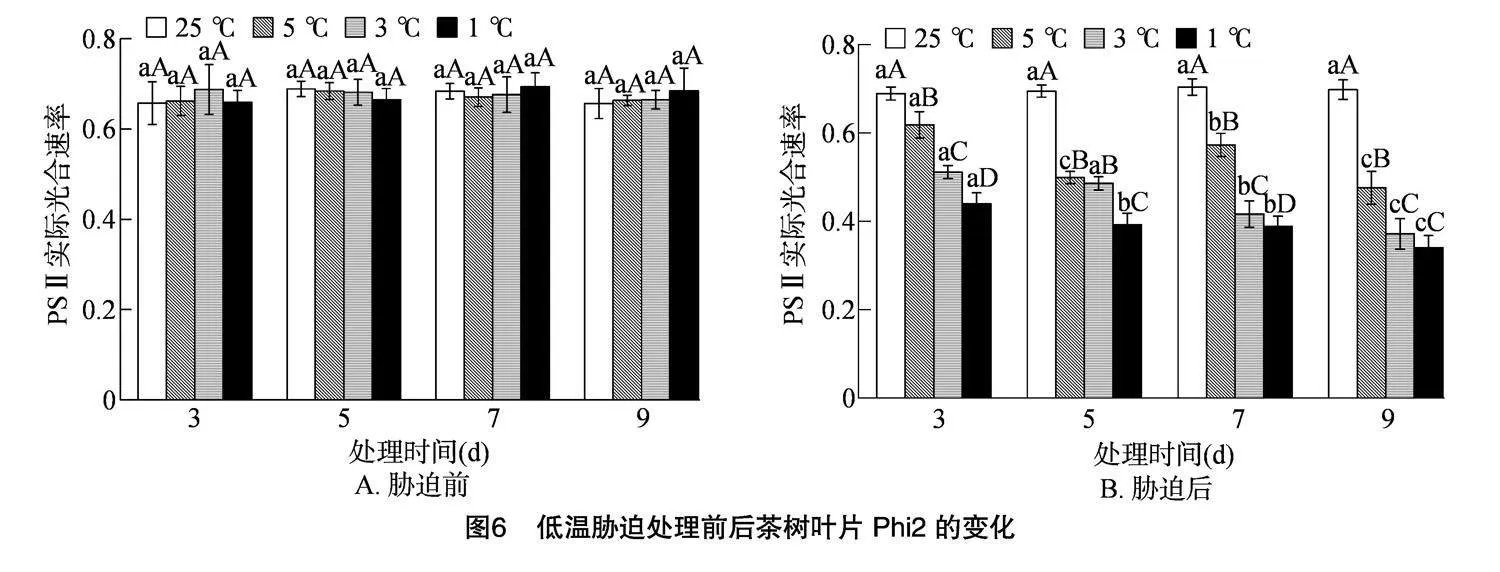
2.4.4" 低温胁迫对茶树叶片Phi2的影响
Phi2代表PSⅡ的实际光合效率,可以直接反映光能转换ATP和NADPH并最终合成糖的能力。由图 6-A 可知,在低温胁迫处理前各处理间的Phi2无显著差异,Phi2为0.657~0.694。由图6-B可知,随着胁迫时间的延长,3、1 ℃的Phi2呈不断下降趋势,而 5 ℃ 的Phi2则呈先降后升再降的趋势,5 ℃处理 9 d 时Phi2最低,为0.476。随着处理温度的降低,各处理的Phi2均呈下降趋势,在4个胁迫时间下的3个低温处理均与CK存在显著差异,在胁迫时间为3 d时,5、3、1 ℃较CK分别降低了10.25%、25.81%、36.22%;在胁迫时间为5 d时,5、3、1 ℃较CK分别降低了28.18%、30.03%、43.51%;在胁迫时间为7 d时,5、3、1 ℃较CK分别降低了18.64%、40.84%、44.78%;在胁迫时间为9 d时,5、3、1 ℃较CK分别降低了31.85%、46.77%、51.24%。研究表明黄金茶1号的Phi2总体随温度的降低和胁迫时间的延长呈下降趋势。
2.4.5" 低温胁迫对茶树叶片NPQ的影响
由图 7-A 可知,在低温胁迫处理前各处理间的NPQ无显著差异,NPQ为0.967~1.000。由图7-B可知,随着胁迫时间的延长,3、1 ℃的NPQ呈不断上升的趋势,而5 ℃的NPQ呈先上升后下降的趋势,5 ℃处理 7 d 时NPQ最高,为1.475。随着处理温度的降低,各处理的NPQ均呈上升趋势,在4个胁迫时间下的3个低温处理均与CK存在显著差异,在胁迫时间为3 d时,5、3、1 ℃较CK分别增加了22.13%、39.38%、52.89%;在胁迫时间为5 d时,5、3、1 ℃较CK分别增加了34.43%、54.27%、62.61%;在胁迫时间为7 d时,5、3、1 ℃较CK分别增加了42.09%、54.27%、67.71%;在胁迫时间为 9 d 时,5、3、1 ℃较CK分别增加了32.06%、52.66%、70.99%。研究表明黄金茶1号的NPQ总体随温度的降低和胁迫时间的延长呈上升趋势。
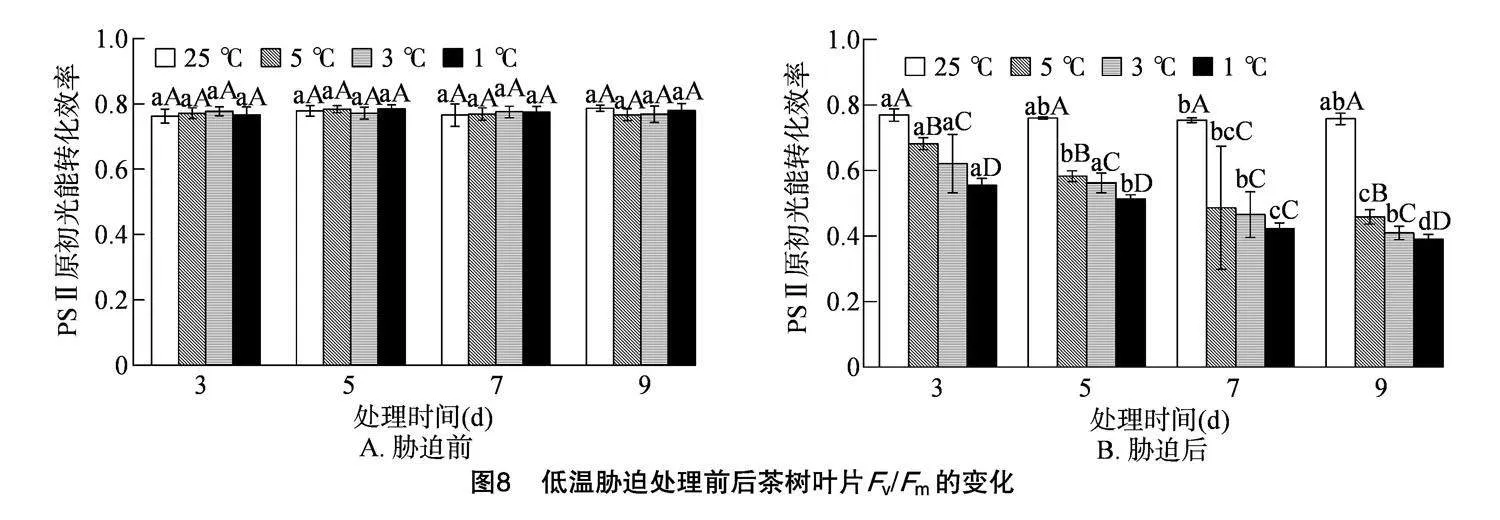
2.4.6" 低温胁迫对茶树叶片Fv/Fm的影响
Fv/Fm 的高低代表植物在逆境条件下的受伤害情况,值越高表明植株生长状况较好; 值越低表明植物受到的伤害越大。由图8-A可知,在正常条件下,茶树叶片的 Fv/Fm 比较稳定,在0.762~0.786之间,不存在显著差异。由图8-B可知,随着胁迫时间的延长,5、3、1 ℃的Fv/Fm呈不断下降的趋势。随着处理温度的降低,各处理的Fv/Fm均呈下降趋势,在胁迫时间为3 d时,5、3、1 ℃与CK存在显著性差异,较CK分别下降了11.39%、19.29%、27.88%;在胁迫时间为5 d时,5、3、1 ℃与CK存在显著性差异,较CK分别下降了23.34%、25.99%、32.45%;在胁迫时间为7 d时,5、3、1 ℃之间无显著差异,但与CK存在显著性差异,较CK分别下降了35.44%、38.21%、43.87%;在胁迫时间为9 d时,5、3、1 ℃与CK存在显著性差异,较CK分别下降了39.47%、45.92%、48.53%。研究表明黄金茶1号的Fv/Fm总体随温度的降低和胁迫时间的延长呈下降趋势。
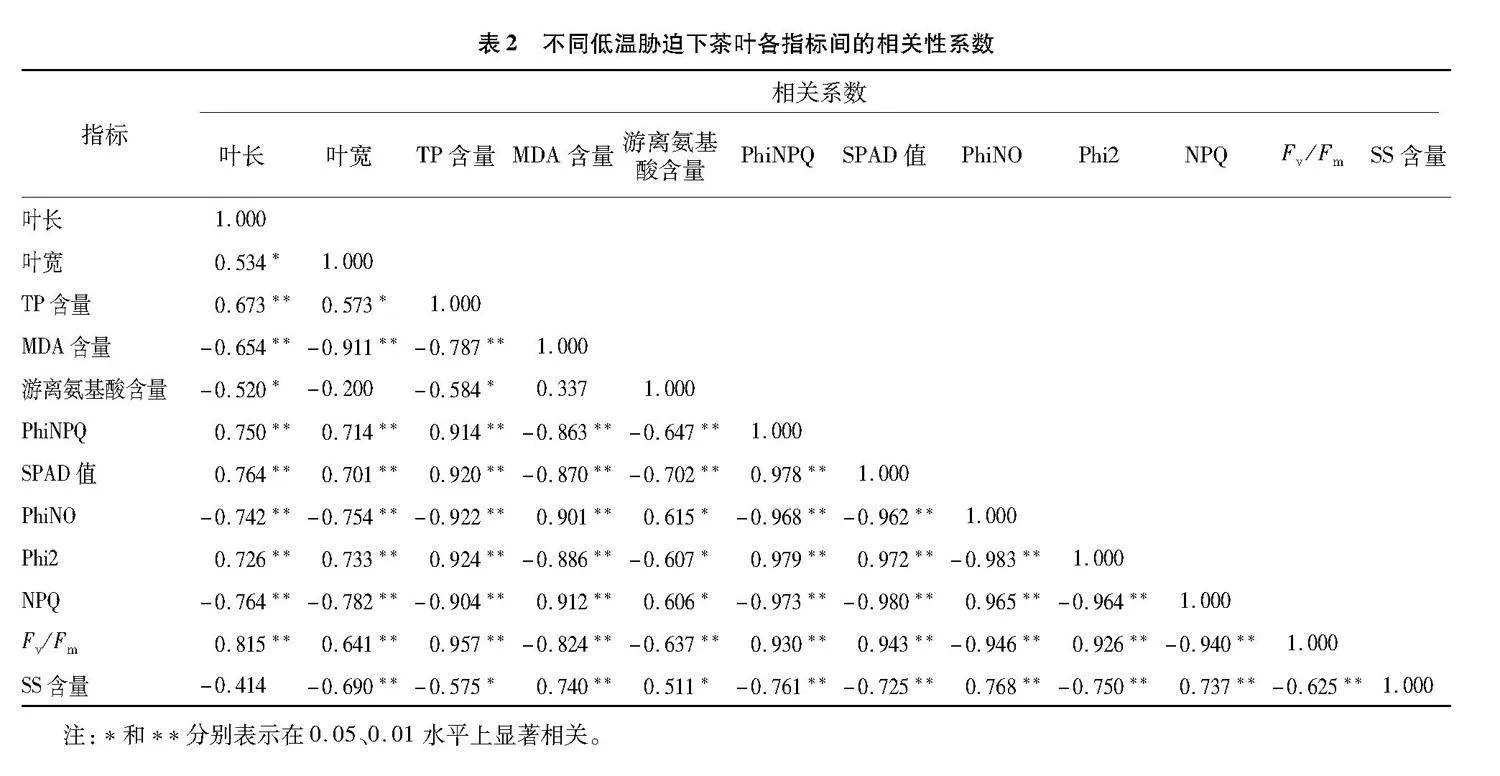
2.5" 不同低温胁迫下茶叶各项指标的相关性分析
由表2可知,叶长与叶宽呈显著正相关关系,与TP含量、PhiNPQ、SPAD值、Phi2、Fv/Fm呈极显著正相关关系,与游离氨基酸呈显著负相关关系,与MDA含量、PhiNO、NPQ呈极显著负相关关系;叶宽与TP含量呈显著正相关关系,与PhiNPQ、SPAD值、Phi2、Fv/Fm呈极显著正相关关系,与MDA含量、PhiNO、NPQ、SS含量呈极显著负相关关系;TP含量与游离氨基酸含量、SS含量呈显著负相关关系,与PhiNPQ、SPAD值、Phi2、Fv/Fm呈极显著正相关关系,与MDA含量、PhiNO、NPQ呈极显著负相关关系;MDA含量与PhiNO、NPQ、SS含量呈极显著正相关关系,与PhiNPQ、SPAD值、Phi2、Fv/Fm呈极显著负相关关系;游离氨基酸含量与PhiNO、NPQ、SS含量呈显著正相关关系,与Phi2呈显著负相关关系,与PhiNPQ、SPAD值、Fv/Fm呈极显著负相关关系;PhiNPQ与SPAD值、Phi2、Fv/Fm呈极显著正相关关系,与PhiNO、NPQ、SS含量呈极显著负相关关系;SPAD值与Phi2、Fv/Fm呈极显著正相关关系,与PhiNO、NPQ、SS含量呈极显著负相关关系;PhiNO与NPQ、SS含量呈极显著正相关关系,与Phi2、Fv/Fm 呈极显著负相关关系;Phi2与Fv/Fm呈极显著正相关关系,与NPQ、SS含量呈极显著负相关关系;NPQ与SS含量呈极显著正相关关系,与Fv/Fm呈极显著负相关关系;Fv/Fm与SS含量呈极显著负相关关系。
3" 讨论与结论
光合作用是植物进行能量转换和物质合成的主要方式,而叶片是进行光合作用的主要场所。茶树属于叶用植物,光合作用的利用效率对茶叶的品质和产量有很大的影响,其中叶绿素荧光参数对茶树的生长发育和逆境胁迫研究具有重要意义。茶多酚和游离氨基酸含量是茶叶中重要的品质指标,逆境胁迫下会影响茶叶的口感和质量,本研究发现在低温胁迫下,茶多酚含量与CK相比有所下降,游离氨基酸含量随着处理温度的降低总体呈先上升后下降的趋势,这与叶锦培等的研究结果[19]相似。MDA含量的高低反映逆境胁迫下植物膜系统受损害的程度;SS含量不仅反映植物在逆境下的抗性,也是植物体内重要的渗透调节物质,本研究结果表明,在低温胁迫下,MDA含量随着温度的降低及胁迫时间的延长而上升;SS含量随温度的降低而上升,随胁迫时间的延长呈先升后降的趋势。这与低温胁迫下王晓龙等的研究结果[20-22]相似。
叶绿素荧光参数的变化不仅可以反映植物叶片光合作用的原初反应,还可作为逆境胁迫研究的重要指标, 其中叶绿素含量的高低反映了植物光合能力的强弱,也是重要的光合色素之一,不仅可以作为植株健康的标准,还可以反映在逆境条件下植株的承受程度[23-24]。本研究结果表明,低温胁迫后茶树的SPAD值随温度的降低和胁迫时间的延长呈下降趋势,这与前人的研究结果[25]相似。Fv/Fm反映PSⅡ原初光能转化效率,该指标不仅能反映光能转化为化学能的效率,还能够反映植物在自然条件下是否受到胁迫。本试验结果表明,在低温胁迫下,茶树叶片的Fv/Fm与CK相比均呈下降趋势,说明了在低温胁迫下茶树PSⅡ系统遭到了破坏,叶片光能转化效率受阻,导致光合色素含量不足从而产生光抑制现象,这与黄艳花等的研究结果[25-26]相似。罗蛟的研究表明,黄瓜叶片在低温光胁迫下会致使Y(Ⅱ)和Y(NPQ)下降,Y(NO)升高[27];范思静等的研究表明,油菜叶片在低温胁迫下Fv/Fm、Y(NPQ)、NPQ均呈下降趋势,而Y(NO)呈上升趋势[28]。本研究结果表明茶树叶片在低温胁迫下PhiNO随温度的降低和胁迫时间的延长逐渐上升,PhiNPQ随温度的降低和胁迫时间的延长呈下降趋势,说明茶叶PSⅡ截获光能能力受损,主动避免光损伤的能力减弱。本研究发现在低温胁迫下茶树叶片NPQ与对照相比有所上升,而Phi2随着温度的降低和胁迫时间的延长而降低,表明低温限制了茶树叶片体内有机物质的合成和光合产物的积累,这与侯晓敏的研究结果[29]相似。
综上所述,在低温胁迫下,低温处理组随着温度的降低和胁迫时间的延长,叶长、叶宽、TP含量、PhiNPQ、SPAD值、Phi2、Fv/Fm总体来说均呈不同程度的下降趋势,MDA含量、PhiNO、NPQ总体呈上升趋势;游离氨基酸含量总体随温度降低呈先升后降的趋势,随胁迫时间变化不同;SS含量随温度的降低逐渐上升,随胁迫时间的延长呈先升后降的趋势,说明茶树叶片在低温胁迫下会导致叶绿素含量降低,叶绿素荧光参数发生改变,致使植物光合效率下降,从而抑制茶树叶片的光合作用和光合电子的传递,造成PSⅡ的活性受到损伤,导致植物出现光抑制现象,对茶树的生长发育和品质造成一定的损失。由于植物多功能仪对测定环境要求很高,稍有偏差都会对试验数据造成误差,而且其测量数据只包含了PS Ⅱ的一部分指标,并未涵盖全部指标,可见植物叶绿素荧光参数的测定结果还存在一定的不足,所以后续试验会结合叶绿素荧光仪综合开展研究。
参考文献:
[1]雷" 雨,段继华,黄飞毅,等. 茶树杂交F1真假杂种的SSR鉴定及遗传多样性分析[J]. 植物遗传资源学报,2021,22(3):748-757.
[2]刘" 振,宁" 静,赵" 洋,等. 黄金茶5个茶树品种(品系)的主要植物学特征与经济性状比较[J]. 茶叶通讯,2021,48(4):629-633.
[3]肖" 健,刘" 慧,郑福维,等. 不同海拔保靖黄金茶品质形成的气象因子初探[J]. 农学学报,2021,11(3):68-73.
[4]李梦菡,吉艳艳,张丽平,等. 植物激素在茶树应对非生物逆境中的作用研究进展[J]. 中国茶叶,2022,44(10):8-15.
[5]王新超,王" 璐,郝心愿,等. 茶树抗寒机制研究进展与展望[J]. 茶叶通讯,2022,49(2):139-148.
[6]Ni Z Q,Jin J,Ye Y,et al. Integrative transcriptomic and phytohormonal analyses provide insights into the cold injury recovery mechanisms of tea leaves[J]. Plants,2022,11(20):2751.
[7]李" 鑫,李梦菡,余红伟,等. 外源2,4-表油菜素内酯诱导茶树低温抗性的生理机制研究[J]. 茶叶通讯,2022,49(3):283-291.
[8]孙" 霞,宋大鹏,刘加英,等. 冻害胁迫下水处理对茶树抗寒生理指标的影响[J]. 中国茶叶,2022,44(4):41-49.
[9]曹正邓渊,文双雅,高志强. 甘蓝型油菜叶绿素荧光参数的高光谱预测[J]. 江苏农业科学,2023,51(18):185-190.
[10]Li D X,Liu H,Gao S K,et al. Effect of post-drought rehydration on winter wheat fluorescence and photosynthetic indices under different levels of nitrogen application[J]. Water,2023,15(2):305.
[11]王兰兰,李" 琦,宋晓卉,等. 环境条件对植物叶绿素荧光参数影响研究进展[J]. 沈阳师范大学学报(自然科学版),2019,37(4):362-367.
[12]Qin J Q,Jiang X D,Qin J H,et al. Effects of lead pollution on photosynthetic characteristics and chlorophyll fluorescence parameters of different populations of Miscanthus floridulus[J]. Processes,2023,11(5):1562.
[13]李庆会,徐" 辉,周" 琳,等. 低温胁迫对2个茶树品种叶片叶绿素荧光特性的影响[J]. 植物资源与环境学报,2015,24(2):26-31.
[14]黄丽芳,龙宇宙,李金芹,等. 低温胁迫对小粒种咖啡幼苗光合及叶绿素荧光特性的影响[J]. 分子植物育种,2024,22(14):4706-4714.
[15]何榜眼,刘世男,杨" 梅,等. 低磷胁迫对大花序桉幼苗叶片生理指标的影响[J]. 西南农业学报,2022,35(2):418-424.
[16]岳俊芹,张素瑜,李向东,等. 低温胁迫对小麦叶绿素荧光参数及产量的响应[J]. 麦类作物学报,2021,41(1):105-110.
[17]王" 馨,易" 拓,朱杰辉,等. 低温胁迫对木薯‘F200’幼苗叶片亚显微结构及其叶绿素荧光参数变化的影响[J]. 热带作物学报,2021,42(7):1925-1931.
[18]王晓龙,冯" 鹏,杨" 曌,等. 寒冷环境下不同苜蓿叶绿素荧光参数变化[J]. 饲料研究,2023,46(13):126-129.
[19]叶锦培,黄爱萍,唐世斌,等. 铝与温度对六堡茶茶叶品质的影响[J]. 湖北农业科学,2020,59(3):97-101.
[20]王晓龙,杨" 曌,来永才,等. 8个苜蓿品种对低温胁迫的生理响应及抗寒性评价[J]. 中国草地学报,2023,45(8):60-69.
[21]王茹华,丁久敏,张启发. 低温胁迫对几种月季砧木生理特性的影响[J]. 北方园艺,2023(12):59-65.
[22]王" 莹. 云雾贡茶低温胁迫下的转录组分析及CsPPO基因的功能鉴定[D]. 贵阳:贵州大学,2021:29-36.
[23]路佳明,任红茹,赵" 灿,等. 低温冷害对水稻孕穗期叶片光合特性及抗氧化酶系统的影响[J]. 扬州大学学报(农业与生命科学版),2023,44(4):15-23,36.
[24]崔" 晓,陈文彬,柳革命,等. 茶树的作物水分胁迫指数、光合有效辐射强度与光合参数的关系[J]. 江苏农业科学,2022,50(10):140-144.
[25]黄艳花,杨" 锐,孙庆海. 温度对裂片石莼生长及叶绿素荧光特性的影响[J]. 生物技术通报,2016,32(7):99-105.
[26]Habibi G. Effects of high light and chilling stress on photosystem Ⅱ efficiency of Aloe vera L. plants probing by chlorophyll a fluorescence measurements[J]. Iranian Journal of Science and Technology,Transactions A:Science,2019,43(1):7-13.
[27]罗" 蛟. 生长条件对黄瓜光系统低温抗性的影响[D]. 泰安:山东农业大学,2022:14-20.
[28]范思静,王亚男. 低温胁迫对油菜叶片光能吸收转换利用的影响[J]. 安徽农业科学,2021,49(24):53-54,59.
[29]侯晓敏. 干旱胁迫下喷施DPC对大豆生理特性的影响[D]. 哈尔滨:东北农业大学,2022:32-38.
猜你喜欢
江苏农业科学(2017年1期)2017-02-27 10:52:12
现代园艺(2016年7期)2017-01-09 14:06:28
农家科技下旬刊(2016年9期)2016-12-15 10:31:02
现代农业科技(2016年19期)2016-12-12 16:20:48
现代农业科技(2016年19期)2016-12-12 15:25:19
现代农业科技(2016年19期)2016-12-12 15:16:23
现代农业科技(2016年19期)2016-12-12 15:09:06
农业与技术(2016年19期)2016-12-12 03:13:59
价值工程(2016年31期)2016-12-03 23:54:47
中国中药杂志(2016年20期)2016-11-19 12:30:29
