
茶树一种新枝条枯萎病病原菌鉴定及防治药剂筛选
2024-11-08 00:00:00王娟涂一怡吕务云陈易佳李士朴王玉春陈雅楠
茶叶科学 2024年5期


摘要:在浙江省瑞安市茶园种植的嘉茗1号茶树上,夏秋季大面积发生一种未见报道的枝条枯萎病,发病枝条出现褐色不规则病斑,病斑可扩大蔓延至芽叶,造成茶芽枯萎,叶片卷曲皱缩,进而整个枝条枯萎死亡。利用组织分离法从发病枝条上分离得到1株真菌,结合菌株的形态特征以及ITS、TUB2和TEF1-α序列构建的多基因系统发育树分析,鉴定该菌株为梭孢葡萄座腔菌(Botryosphaeria fusispora)。室内致病力测试表明,B. fusispora通过伤口侵染茶树枝条造成枯萎病的发生。以上结果表明,作为新纪录种,B. fusispora是引起茶树该新病害发生的病原菌。采用室内菌丝生长速率抑制法测定了3种杀菌剂(百菌清、甲基硫菌灵和吡唑醚菌酯)对病原菌的抑制效果。结果表明,甲基硫菌灵和吡唑醚菌酯对B. fusispora均有较强的毒力,EC50值分别为1.91 μg·mL-1和2.25 μg·mL-1。
关键词:茶树;枝条枯萎病;葡萄座腔菌属;室内毒力测定
中图分类号:S571.1;R435.711 文献标识码:A 文章编号:1000-369X(2024)05-807-09
Identification of the Pathogen Causing New Twig Wilting on Tea Plants and Screening of Control Chemicals
WANG Juan1, TU Yiyi1, LÜ Wuyun1, CHEN Yijia1, LI Shipu2, WANG Yuchun1*, CHEN Yanan1*
1. College of Tea Science and Tea Culture, Zhejiang A & F University, Hangzhou 311300, China;
2. Zhejiang Wanfu Tea Industry Development Co. Ltd., Wenzhou 325200, China
Abstract: During summer and autumn, serious new twig wilting occurs on a large scale in ‘Jiaming No. 1’ tea gardens of Ruian City, Wenzhou, Zhejiang Province, China. The disease initially appears as irregular brown spots on the twigs, which later spread to shoots and leaves, causing the tea shoots to wilt and the leaves to curl and shrink, and eventually the whole branch to die. In this study, a strain was isolated and purified using the tissue isolation method. Combining the morphological characteristics with the phylogenetic analyses based on the sequenyx68PXNA42hSJfz8Am0gaQ==ces of the internal transcribed spacer regions (ITS), β-tubulin (TUB2), and the translation elongation factor1 alpha (TEF1-α), the isolated strain was identified as Botryosphaeria fusispora. Laboratory pathogenicity tests show that B. fusispora was the pathogen causing the twig wilting disease present on ‘Jiaming No. 1’. These results indicate that B. fusispora can be a new record species causing twig wilting on tea plants. In addition, mycelial growth inhibition tests were conducted to examine the sensitivity of the pathogen to three commercial fungicides, including chlorothalonil, thiophanate-methyl and pyraclostrobin. Among the tested fungicides, thiophanate-methyl was found to be the most effective in suppressing the radial growth of the strain, with an EC50 of 1.91 μg·mL-1, followed by pyraclostrobin, with an EC50 of 2.25 μg·mL-1.
Keywords: tea plants, twig wilting, Botryosphaeria spp., indoor toxicity test
茶树[Camellia sinensis (L.) O. Kuntze]属于山茶科山茶属植物[1],是我国重要的经济作物。2023年我国干毛茶总产量达333.95万t,同比增长4.98%[2]。我国茶园主要分布在气候温暖、雨量充沛的地区,这为病害的发生提供了有利环境。据统计,目前我国茶树病害约有140种,包括茶炭疽病、茶饼病、茶云纹叶枯病等叶部病害;茶枝梢黑点病、茶黑痣病、茶粗皮病等枝条病害;茶根癌病、茶红根腐病、茶苗白绢病等根部病害[3-5]。近年来,由于茶园种植面积不断扩大、茶树品种更替以及气候变暖和湿度增高等因素,陆续发现一些新的茶树病害。Jiang等[6]在安徽茶园中首次发现Muyocopron laterale能够引起茶树叶枯病;Chen等[7]对发生于重庆二圣地区的茶叶斑病样本进行分离,并结合形态特征和系统发育分析,鉴定病原菌为附球菌属真菌(Epicoccum layuense);Yang等[8]在湖北省恩施首次发现灰葡萄孢菌(Botrytis cinerea)在中国引起茶树灰霉病。2023年秋季,笔者在浙江省瑞安市高楼镇茶园嘉茗1号枝条上发现一种从未报道过的新病害,该病害多发于未木质化的枝条嫩梢,严重时蔓延至茶芽和叶片上,造成叶片卷曲皱缩和枝条枯萎,对茶叶产量和品质造成一定影响。然而,该病害的病原菌尚不清楚。
葡萄座腔菌科(Botryosphaeriaceae)真菌分布广泛,能够侵染多种寄主植物造成枝干腐烂、溃疡、流胶和枯死等症状,严重时甚至会导致植株死亡[9]。该科真菌最早由Theissen等[10]作为假球壳科(Pseudosphaeriaceae)的一个亚科引入,之后,Schoch等[11]通过构建多基因系统发育树,提出了一个新的葡萄座腔菌目(Botr-yosphaeriales),且只容纳葡萄座腔菌科这一单独的科。Slippers等[12]根据系统发育分析发现葡萄座腔菌科共包含17个属(Botryosphaeria spp.),之后Phillips等[13]详细描述了各个属的形态特征并作为分类鉴定依据。根据Rathnayaka等[14]报道,早在1983年就有研究发现葡萄座腔菌科真菌侵染山茶属(Camellia spp.)植物。2016年,Jayawardena等[15]首次从我国福建省漳州市茶树坏死叶片上分离并鉴定了两株葡萄座腔菌(Botryosphaeria dothidea);张永乐等[16]从山东日照东港区茶区茶树叶部发现一种茶褐枯病,结合形态学、多基因序列分析及柯赫氏法则等,验证其病原菌为围小丛壳菌山茶专化型(Glomerella cingulata f. sp. camelliae)和B. dothidea。2020年,李力等[17]收集福建省武夷山地区茶园茶树叶斑病病叶样品进行分离培养,根据多位点序列和聚类分析鉴定其病原真菌为B. dothidea。目前,Botryosphaeria属真菌主要引起茶树叶部病害,尚未报道可引起枝条发病。因此,本研究在新病害症状的基础上,对病原菌进行分离培养、鉴定、致病性测定,以及药剂敏感性检测,明确该病害病原菌并提出候选防控药剂,为未来该茶树枝条病害的科学防控提供参考。
1 材料与方法
1.1 试验材料
2023年9月27日,未木质化的发病枝条嫩梢采自浙江省温州市瑞安市高楼镇茶园(27°45′ N,120°17′ E),树龄为20 a的嘉茗1号(Jiaming No. 1,JM1)茶树品种。室内致病性测试茶树品种为龙井43(Longjing43,LJ43)、中茶108(Zhongcha108,ZC108)、中茶302(Zhongcha302,ZC302)及JM1。
1.2 试验方法
1.2.1 病原菌的分离与纯化
采用常规组织分离法分离病原菌[18]。从发病枝条病健交界处切取组织接种至马铃薯葡萄糖琼脂(Potato dextrose agar,PDA)培养基上,放入25 ℃培养箱避光培养3 d。长出菌落后,挑取不同形态菌落边缘的菌丝转移到新的PDA平板上进行纯化,获得菌落形态一致的菌株。将分离纯化的菌株转移至PDA斜面培养后于4 ℃冰箱中保存备用。
1.2.2 形态学观察
将各菌株在25 ℃黑暗条件下培养3 d后,利用灭菌的打孔器从菌落边缘打取直径为5 mm的菌饼,分别接种至新的PDA及2%麦芽汁琼脂(Malt extract agar,MEA)培养基平板中央,25 ℃避光培养,14 d后观察菌落形态特征,并采用十字交叉法测量菌落直径。菌落生长速率以测量菌落直径除以培养天数表示。每个试验设置5个生物学重复,并独立重复5次。菌落生长速率测定试验重复5次。将各菌株接种至含茶树幼嫩枝条的2%水琼脂(Water agar,WA)培养基上,25 ℃避光培养4周,于SOPTOP-
CX40RFL显微镜下观察分生孢子形态,并拍照记录。
1.2.3 DNA提取及PCR扩增和测序
将保存在4 ℃冰箱中的菌株接种到PDA平板上,25 ℃黑暗条件下培养7 d,用DEB法[19]少量快速提取菌丝基因组DNA,于﹣20 ℃冰箱保存。
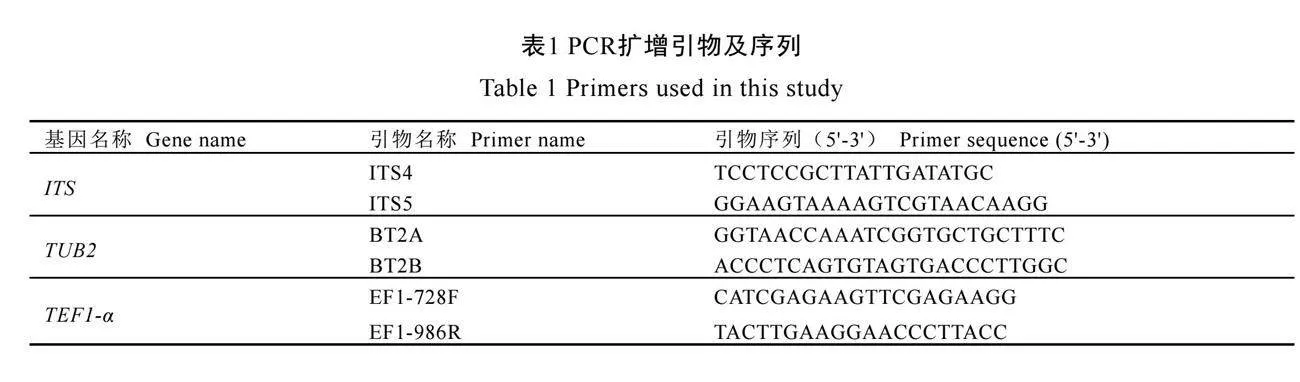
选取核糖体内转录间隔区(Internal transcribed spacer,ITS)、β-微管蛋白2(β-tubulin,TUB2)和翻译延伸因子(Translation elongation factor 1-alpha,TEF1-α)3个DNA片段进行PCR扩增,所用引物及序列[20]见表1。
PCR反应体系(25 μL):12 μL 2×Taq Master Mix,1 μL 10 μmol·L-1上游引物,1 μL 10 μmol·L-1下游引物,10 μL ddH2O和1 μL基因组DNA。PCR反应程序:94 ℃预变性5 min;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸60 s,32个循环;72 ℃延伸10 min。取7 μL PCR产物采用1%琼脂糖凝胶进行电泳检测,将目的条带样品送至杭州有康生物科技有限公司进行纯化测序。
1.2.4 系统发育分析
测序获得的基因序列在NCBI网站(www.ncbi.nlm.nih.gov)进行BLAST比对。利用MAFFT v. 7和MEGA v. 5软件进行多序列比对、拼接和手动矫正[21-22]。利用最大似然法在FastTree软件上构建系统发育树[23]。
1.2.5 致病性测定
以LJ43、JM1、ZC302和ZC108的健康新生一芽四叶枝条为材料,进行表面消毒后,挑取PDA平板上培养7 d的菌落边缘菌饼(直径为5 mm),接种于茶树枝条。接种方式采用有伤接种和无伤接种两种,每个菌株设置3个重复,对照组用无菌琼脂块处理。接种样品置于人工气候培养箱中25 ℃黑暗保湿培养3 d,观察发病情况。若观察到相似病斑,则对其重新分离并确认分离物与接种体是否为同一菌株,以完成柯赫氏法则。致病性试验重复2次。
1.2.6 杀菌剂敏感性测定
采用菌丝生长速率抑制法测定病原菌对3种杀菌剂的敏感性,包括75%可湿性粉剂百菌清、25%悬浮剂吡唑醚菌酯和70%可湿性粉剂甲基硫菌灵。在PDA培养基中分别加入不同浓度的杀菌剂,其中百菌清的质量浓度梯度设置为0.1、1、10、50、100、500、1 000 µg·mL-1;甲基硫菌灵的质量浓度梯度设置为0.01、0.1、1、2、5、10、20 µg·mL-1;吡唑醚菌酯浓度梯度为0.01、0.1、1、2、5、10、20 µg·mL-1。将各菌株接种至杀菌剂不同浓度梯度的PDA平板(Treated,T)以及不含有杀菌剂的PDA平板(Control,C)上,25 ℃黑暗培养3 d后,采用十字交叉法测量菌落直径,按照以下公式计算抑菌率(Inhibition,I):
I=(C-T)/(C-0.5)×100%
其中,I为菌丝生长抑菌率(%),C为对照组菌落直径平均值(mm),T为试验组菌落直径平均值(mm)。使用SPSS软件求取毒力回归方程并计算得出半数效应浓度(Median effect concentration,EC50)值。每个浓度梯度试验设置3个重复,并独立重复3次。
2 结果与分析
2.1 病害症状观察
该病害多发于未木质化的枝条嫩梢(图1),
发病部位初期呈现褐色不规则病斑(图1a),随后病斑不断扩大,可蔓延至发病枝条的茶芽和叶片上(图1b~1d),从叶片基部扩散至整个叶片,造成叶片卷曲皱缩,最终形成大面积枯萎组织(图1e~1f)。此外,在湿度较高时,病害枝条表面出现白色霉层(图1g)。
2.2 病原菌形态学特征描述
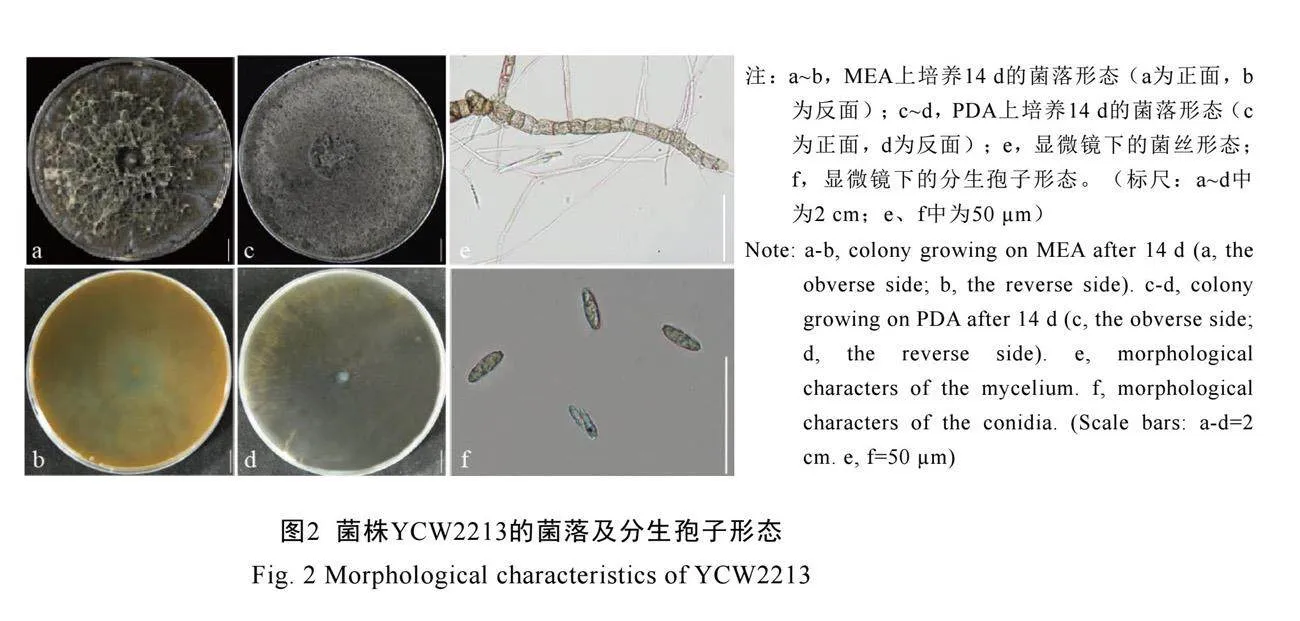
通过对发病枝条进行组织分离,纯化后获得菌株YCW2213。该菌株在培养基上的菌落蓬松,边缘不规则,气生菌丝最初为白色,培养10~14 d后颜色逐渐变深直至深灰色,并团成絮状,背面呈暗棕色(图2a~2d)。普通光学显微镜下观察,发现生长后期菌丝可膨大呈结节状
(图2e)。在MEA平板上生长速率为10 mm·d-1,在PDA平板上生长速率为15 mm·d-1。PDA和MEA平板上未观察到分生孢子,但在WA平板培养4周后可观察到极少量分生孢子。分生孢子透明,无分隔,呈椭圆形,两端尖(图2f)。
2.3 病原菌系统发育学分析
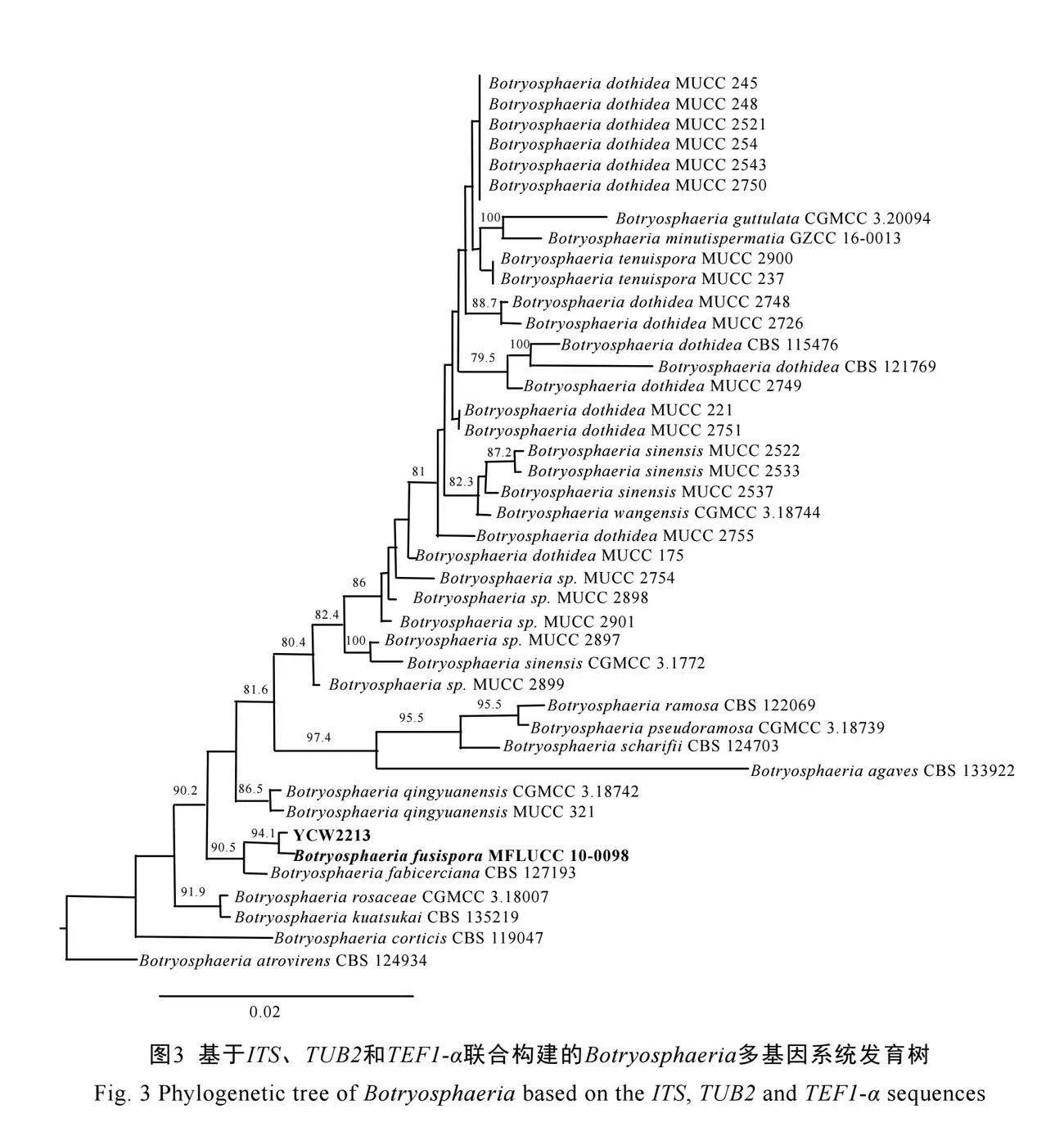
对测序获得的YCW2213菌株的ITS、TUB2和TEF1-α序列,首先在NCBI上进行BLAST,根据BLAST结果下载相近种的序列作为参考菌株,以Cophinforma atrovirens为外类群构建多基因系统发育树。结果表明,YCW2213与B. fusispora聚为1枝,且自展值达到94%(图3)。结合其形态学特征,最终将病原真菌YCW2213
鉴定为梭孢葡萄座腔菌(B. fusispora)。
2.4 致病性分析
将YCW2213菌株分别接种至LJ43、JM1、ZC302和ZC108新生健康枝条上,进行室内致病性试验。结果显示,有伤接种YCW2213处理3 d后,茶树枝条上均出现褐色病斑,且随着侵染时间延长,病斑不断扩大(图4)。此外,相较于其他品种,LJ43枝条病斑更大,且病斑表面出现白色霉层(图4d),发病症状与田间发病情况一致。
对发病枝条进行再分离,获得的菌株在PDA培养基上的菌落形态特征和2.2章节中B. fusispora的一致,而对照处理的枝梢未能分离
获得与B. fusispora一致的菌落。再分离菌株的ITS、TUB2和TEF1-α序列也与B. fusispora一致。B. fusispora能够引起健康茶树枝条发病,且在发病枝条病组织上能够再分离到该菌,以上结果表明,此病害是由B. fusispora通过伤口侵染所致。
2.5 杀菌剂敏感性测定
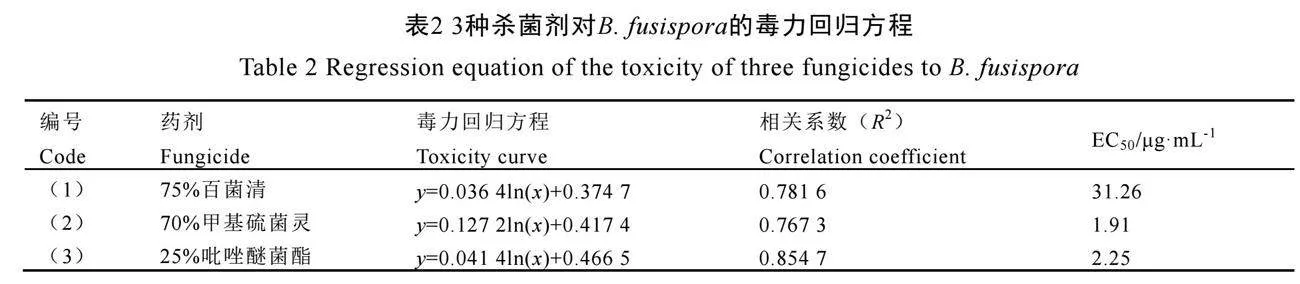
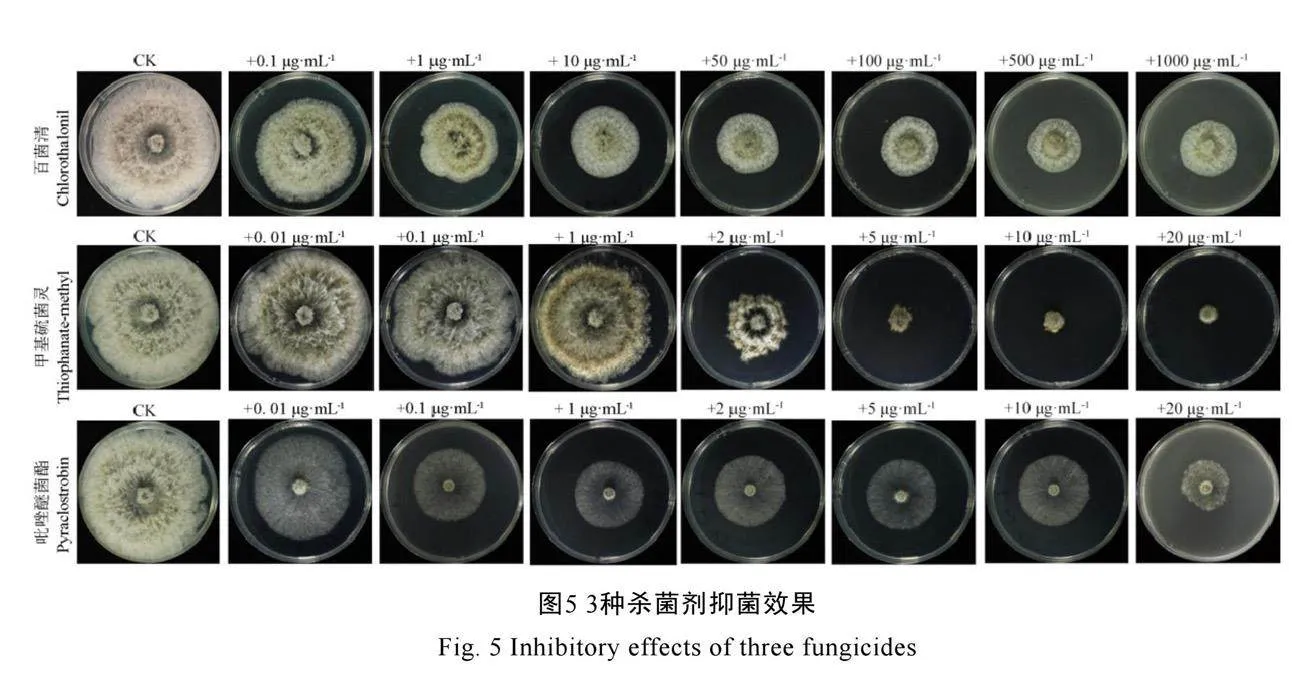
为了筛选田间防治该病害的药剂,本研究测试了B. fusispora菌株YzccGPr9+dnfAGH4bKvqL1g==CW2213对75%百菌清、70%甲基硫菌灵和25%吡唑醚菌酯等3种常用杀菌剂的敏感性。结果显示,YCW2213对5 μg·mL-1以上的甲基硫菌灵最敏感(图5)。通过测量处理组和对照组的菌落生长直径,得出3种杀菌剂的毒力回归方程及EC50值(表2)。3种杀菌剂的EC50值由小到大的顺序为70%甲基硫菌灵(1.91 μg·mL-1)、25%吡唑醚菌酯(2.25 μg·mL-1)、75%百菌清(31.26 μg·mL-1)。结果表明,70%甲基硫菌灵对B. fusispora菌株YCW2213的抑菌效果最好,其次是25%吡唑醚菌酯,抑菌效果较弱的是75%百菌清。
3 讨论
在浙江省瑞安市高楼镇茶园,从大面积出现的茶树嘉茗1号枯萎坏死枝条上分离得到1株真菌,其菌落形态不规则,且随着生长菌落从白色逐渐变深至深灰色(图2a~2d),分生孢子单孢、透明、无隔、椭圆形、两端尖(图2f),这与前人报道的B. fusispora形态特征一
致[24-25],结合构建的多基因系统发育树结果(图3),明确该菌株为B. fusispora。我们尝试在PDA或MEA培养基上诱导YCW2213产孢均未成功,而利用含茶树幼嫩枝条的2%水琼脂培养基培养,可诱导其产孢,但产孢数量仍较少,这与葡萄座腔菌属真菌(Botryosphaeria spp.)在自然及人工培育条件下不易产生无性或有性生殖结构有关[26]。由于通过多种方法诱导均无法产生大量孢子,故通过菌饼接种进行室内致病性测试,研究结果表明,该真菌可造成与田间表型一致的病状,且再分离的菌株与侵染菌株一致,说明B. fusispora是本次茶树新病害的病原菌。Botryosphaeria属真菌是世界范围内最广泛、最重要的树木枝干枯死病病原菌之一[27]。B. fusispora最早由Liu等[28]在泰国清莱黎敦山Entada spp.的干树皮中分离获得。随后Li等[27]发现B. fusispora在中国亚热带和热带地区种植的桉树上广泛分布,侵染枝干引起茎溃疡病、茎点枯病和枝枯病。此外,有报道从广东、广西、海南、福建、云南和四川等地的荔枝病果[26]、猕猴桃病果[29]、蓝莓腐烂茎[30]以及香榧病果[31]中分离到B. fusispora,这表明B. fusispora不仅寄主广泛还能够在较大温度范围内生存。可见,B. fusispora可能并非本研究新发现茶树病害的唯一病原菌,且仅在瑞安市高楼镇发现该病害的暴发,其他地区茶园是否已经出现,亦或茶树其他组织是否已经受到危害,需要进一步取样分析,提早做好风险控制。
化学防治仍是当前茶树病害的主要防治措施,室内毒力测定可为大田防治提供一定的理论基础。研究发现与B. fusispora同属的B. dothidea对百菌清[32-33]、甲基硫菌灵[33-34]、吡唑醚菌酯[35]均具有敏感性。因此,本研究采用菌丝生长速率抑制法分析测定了茶树枯萎病菌B. fusispora对这3种杀菌剂的敏感性。结果表明,75%百菌清、70%甲基硫菌灵和25%吡唑醚菌酯对B. fusispora均有毒力,EC50分别为31.26、1.91 µg·mL-1和2.25 µg·mL-1,其中70%甲基硫菌灵抑制效果最好,其次为25%吡唑醚菌酯(表2),这与胡冲等[33]和李诚等[34]的研究结果基本一致。
综上所述,本研究首次发现B. fusispora危害茶树枝条,引起茶树枝条枯萎病;通过室内毒力测定试验发现甲基硫菌灵和吡唑醚菌酯对B. fusispora的抑菌效果较好,可用于田间药效试验研究。本研究结果为预防和控制该茶树枝条枯萎病奠定了理论基础。
参考文献
[1] 马建强, 姚明哲, 陈亮. 茶树种质资源研究进展[J]. 茶叶科学, 2015, 35(1): 11-16.
Ma J Q, Yao M Z, Chen L. Research progress on germ-plasms of tea plant (Camellia sinensis) [J]. Journal of Tea Science, 2015, 35(1): 11-16.
[2] 梅宇, 梁晓. 2023年我国茶叶产销及进出口形势分析[J]. 中国茶叶, 2024, 46(4): 18-26.
Mei Y, Liang X. Analysis of China's tea production, sales, import and export situation in 2023 [J]. China Tea, 2024, 46(4): 18-26.
[3] 徐云, 梅红, 林莉, 等. 茶树病害诊断与防治专家系统研制[J]. 农业系统科学与综合研究, 2003(2): 93-96, 100.
Xu Y, Mei H, Lin L, et al. The study and exploitation of diagnosed and controlled tea disease's expert system [J]. System Sciences and Comprehensive Studies in Agriculture, 2003(2): 93-96, 100.
[4] 陈宗懋, 陈雪芬. 世界茶树病原名录(续)[J]. 茶叶科学, 1989, 29(1): 73-88.
Chen Z M, Chen X F. A world list of pathogens reported on tea plant [J]. Journal of Tea Science, 1989, 29(1): 73-88.
[5] 陈雪芬. 我国茶树病害的发生趋势与绿色防控[J]. 中国茶叶, 2022, 44(6): 7-14.
Chen X F. The occurrence trend and green control of tea diseases in China [J]. China Tea, 2022, 44(6): 7-14.
[6] Jiang H, Zhang M, Zhou Y, et al. First report of Muyocopron laterale causing a new leaf disease of Camellia sinensis in China [J]. Beverage Plant Research, 2022, 2(1): 1-4. doi:10. 48130/bpr-2022-0010.
[7] Chen Y J, Wan Y H, Zou L J, et al. First report of leaf spot disease caused by Epicoccum layuense on Camellia sinensis in Chongqing, China [J]. Plant Disease, 2020, 104(7): 2029. doi: 10.1094/PDIS-09-19-1906-PDN.
[8] Yang D, Yao J, Wang B, et al. First report of Botrytis cinerea causing gray mold on tea (Camellia sinensis) in China [J]. Plant Disease, 2024, 108(1): 203. doi: 10.1094/PDIS-01-23-
0022-PDN.
[9] López-Mora. Etiology of branch dieback and shoot blight of English Walnut caused by Botryosphaeriaceae and Diaporthe species in Southern Spain [J]. Plant Disease, 2020, 104(2): 533-550.
[10] Theissen F, Sydow H. Vorentwürfe zu den Pseudosphaeriales [J]. Annales Mycologici, 1918, 16(1/2): 1-34.
[11] Schoch C L, Shoemaker R A, Seifert K A, et al. A multigene phylogeny of the Dothideomycetes using four nuclear loci [J]. Mycologia, 2006, 98(6): 1041-1052.
[12] Slippers B, Boissin E, Phillips A, et al. Phylogenetic lineages in the Botryosphaeriales: a systematic and evolutionary framework [J]. Studies in Mycology, 2013, 76(1): 31-49.
[13] Phillips A J, Alves A, Abdollahzadeh J, et al. The Botryosphaeriaceae: genera and species known from culture [J]. Studies in Mycology, 2013, 76(1): 51-167.
[14] Rathnayaka A R, Chethana K W T, Phillips A J, et al. First report of Botryosphaeriaceae species on Camellia sinensis from Taiwan with a global checklist of Botryosphaeriaceae species on this host [J]. Chiang Mai Journal of Science, 2021, 48(5): 1199-1223.
[15] Jayawardena R S, Li X H, Xu W, et al. First report of Botryosphaeria dothidea causing leaf necrosis of Camellia sinensis in Fujian Province, China [J]. Plant Disease, 2016, 100(4): 854-854.
[16] 张永乐, 刘会香, 许永玉, 等. 围小丛壳菌山茶专化型与葡萄座腔菌复合侵染引致茶褐枯病[J]. 茶叶科学, 2018, 38(1): 87-93.
Zhang Y L, Liu H X, Xu Y Y, et al. The tea brown blight disease caused by co-infection of Glomerella cingulata f. sp. camelliae and Botryosphaeria dothidea [J]. Journal of Tea Science, 2018, 38(1): 87-93.
[17] 李力, 张渤, 范俐, 等. 侵染茶树的葡萄座腔菌分离鉴定及其拮抗菌筛选[J]. 宁德师范学院学报(自然科学版), 2023, 35(1): 75-80.
Li L, Zhang B, Fan L, et al. Isolation, identification and antagonistic bacterium screening of a fungal pathogen Botryosphaeria dothidea isolate infecting tea plant leaves [J]. Journal of Ningde Normal University (Natural Science), 2023, 35(1): 75-80.
[18] Cai L, Hyde K D, Taylor P W J, et al. A polyphasic approach for studying Colletotrichum [J]. Fungal Diversity, 2009, 39(1): 183-204.
[19] 吕务云. 禾谷镰刀菌细胞自噬途径相关基因的功能分析 [D]. 杭州: 浙江大学, 2019.
Lü W Y. Functional analysis of autophagy-related genes in Fusarium graminearum [D]. Hangzhou: Zhejiang University, 2019.
[20] Yukako H, Yuho A, Atsuko S, et al. Taxonomical study of noteworthy species of Botryosphaeria in Japan [J]. Mycobiology, 2021, 49(2): 122-132.
[21] Katoh K, Standley D M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability [J]. Molecular Biology and Evolution, 2013, 30(4): 772-780.
[22] Tamura K, Peterson D, Peterson N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods [J]. Molecular Biology and Evolution, 2011, 28(10): 2731-2739.
[23] Minh B Q, Schmidt H A, Chernomor O, et al. Corrigendum to: IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era [J]. Molecular Biology and Evolution, 2020, 37(5): 1530-1534.
[24] Li G Q, Liu F F, Li J Q, et al. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China [J]. Persoonia, 2018, 40(1): 63-95.
[25] Crous P W, Slippers B, Wingfield M J, et al. Phylogenetic lineages in the Botryosphaeriaceae [J]. Studies in Mycology, 2006, 55(1): 235-253.
[26] 凌金锋. 荔枝病果相关的四属菌物鉴定及分子系统发育分析 [D]. 广州: 华南农业大学, 2019.
Ling J F. Identification and phylogenetic analysis of four fungal genera isolates associated with diseased litchi fruits in China [D]. Guangzhou: South China Agricultural University, 2019.
[27] Li G Q, Slippers B, Wingfield M J, et al. Variation in Botryosphaeriaceae from Eucalyptus plantations in Yunnan Province in southwestern China across a climatic gradient [J]. IMA Fungus, 2020, 11(22): 1-49. doi: 10. 1186/s43008-020-00043-x.
[28] Liu J K, Phookamsak R, Doilom M, et al. Towards a natural classification of Botryosphaeriales [J]. Fungal Diversity, 2012, 57(1): 149-210.
[29] 刘平平, 莫权辉, 叶开玉, 等. 广西猕猴桃果实病害病原菌分离与鉴定[J]. 中国南方果树, 2023, 52(4): 92-99.
Liu P P, Mo Q H, Ye K Y, et al. lsolation and identification of fruit diseases pathogens of kiwifruit in Guangxi [J]. South China Fruits, 2023, 52(4): 92-99.
[30] 褚睿天, 豆志鹏, 贺伟, 等. 引起蓝莓茎溃疡病的葡萄座腔菌属Botryosphaeria二新种[J]. 菌物学报, 2021, 40(3): 473-486.
Chu R T, Dou Z P, He W, et al. Two novel species of Botryosphaerla causing stem canker of blueberries from China [J]. Mycosystema, 2021, 40(3): 473-486.
[31] 叶碧欢, 沈建军, 李海波, 等. 植物提取物对香榧病原真菌的抑菌活性研究[J]. 中国森林病虫, 2023, 42(1): 1-5.
Ye B H, Shen J J, Li H B, et al. Antifungal activity of plant extracts against pathogenic fungi of Torreya grandis ‘Merrillii’ [J]. Forest Pest and Disease, 2023, 42(1): 1-5.
[32] 周英, 欧阳慧, 蒋军喜, 等. 10种杀菌剂对葡萄座腔菌的室内毒力测定[J]. 中国南方果树, 2016, 45(6): 124-125, 130.
Zhou Y, Ouyang H, Jiang J X, et al. Indoor toxicity determination of 10 fungicides to Botryosphaeria dothidea [J]. South China Fruit, 2016, 45(6): 124-125, 130.
[33] 胡冲, 吴道军. 杀菌剂对樱花枝枯病菌葡萄座腔菌的室内毒力测定[J]. 农业与技术, 2020, 40(23): 19-20.
Hu C, Wu D J. Indoor toxicity determination of fungicides on Botryosphaeria dothidea of Cerasus cladosporioides [J]. Agriculture and Technology, 2020, 40(23): 19-20.
[34] 李诚, 蒋军喜, 冷建华, 等. 6种杀菌剂对猕猴桃主要腐烂病菌的室内毒力测定[J]. 中国南方果树, 2012, 41(1): 27-29.
Li C, Jiang J X, Leng J H, et al. Indoor toxicity test of six fungicides to main pathogenic fungi causing fruit rot of kiwifruit [J]. South China Fruits, 2012, 41(1): 27-29.
[35] 赵江, 余知和, 李其利, 等. 芒果葡萄座腔菌对3种杀菌剂的敏感性测定[J]. 南方农业学报, 2021, 52(4): 984-994.
Zhao J, Yu Z H, Li Q L, et al. Susceptibility of Botryosphaeriaceous isolates from mango to three fungicides [J]. Journal of Southern Agriculture, 2021, 52(4): 984-994.
