
雅长保护区老龄林不同林层功能性状多样性及其影响因素分析
2024-10-09 00:00:00王耀仪王宏翔王永强曾文豪叶绍明
南京林业大学学报(自然科学版) 2024年5期
关键词:结构方程模型



摘要:【目的】功能性状多样性对森林生物多样性保护及生态系统功能稳定性的维持具有重要意义。探究不同林层的功能多样性特征及其形成原因,有助于深入理解生物多样性维持机制和植物群落构建过程。【方法】以广西雅长兰科植物国家级自然保护区1.6 hm2常绿阔叶天然老龄林监测样地植被调查数据为基础,运用K-means非层次聚类与高斯核密度估计方法划分林层,采集植物叶片性状和环境因子数据,通过结构方程模型揭示不同林层物种多样性与环境因子(地形、土壤和光照)对功能多样性的直接、间接影响。【结果】①林下层多样性指数显著高于林冠层(Plt;0.05)。②林下层物种多样性对4个功能多样性指数均有显著的直接影响,而林冠层物种多样性仅对功能丰富度指数(FRic)与功能二次熵指数(RAOQ)有显著的直接影响(Plt;0.05)。③林冠开度对林下层FRic指数有极显著的间接效应(β= 0.278,Plt;0.01),对林下层功能均匀度指数(FEve)有极显著的直接效应(β= -0.593,Plt;0.01)。④地形因子主要影响林冠层功能多样性。其中,坡向对林冠层FEve指数(β= -0.420,Plt;0.01)与RAOQ指数(β= -0.300,Plt;0.05)均有显著的直接效应。⑤土壤因子主要通过物种多样性间接影响各林层的功能多样性。其中,林下层功能多样性主要受全磷、有效磷、碱解氮、有效钾含量的影响,而全钾含量对林冠层FRic与RAOQ指数有一定的影响。【结论】研究区内各林层功能多样性存在较大差异。林下层功能多样性主要受物种多样性、林冠开度和土壤养分的影响,而地形因子则是林冠层功能多样性的主要影响因素。本研究结果揭示了不同林层内功能性状多样性对生物及非生物环境变化的响应,拓展了以往对于林分垂直分层多样性的认识,为天然林生物多样性保护提供更进一步的参考依据。
关键词:老龄林;林冠层;林下层;功能性状多样性;结构方程模型(SEM)
中图分类号:S718"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)05-0028-11
Affecting factors analysis of functional diversity at different forest strata in an old growth forest community in Yachang Natural Reserve
WANG Yaoyi, WANG Hongxiang*, WANG Yongqiang, ZENG Wenhao, YE Shaoming
(College of Forestry, Guangxi Key Laboratory of Forest Ecology and Conservation, Guangxi University, Nanning 530004, China)
Abstract: 【Objective】 Functional diversity is crucial for forest biodiversity protection and maintenance of ecosystem stability. Exploring the characteristics and causes of functional diversity variation at different stratum levels will assist in understanding the maintenance mechanisms of biodiversity and plant community assembly processes. 【Method】 Leaf traits and environmental factors were collected from forest plot data from an 1.6 hm2 evergreen broadleaved forest in Guangxi Yachang Orchidaceae National Natural Reserve. K-means partitioning and Gaussian kernel density estimation were used for stratification. The direct and indirect effects of species diversity and environmental factors (light, topography, and soil) on functional diversity were explored in different forest strata using structural equation models. 【Result】 The diversity indices of the understory were greater than those of the canopy (Plt;0.05). Species diversity had a significant direct impact on all four functional diversity indices in the understory layer, whereas only the functional richness index (FRic) and quadratic entropy (RAOQ) indices were affected directly by species diversity in the canopy layer (Plt;0.05). A significant indirect effect (β = 0.278, Plt;0.01) of canopy openness on FRic index and a direct effect (β = -0.593, Plt;0.01) of canopy openness on functional evenness index (FEve) were found for understory trees. Topographic factors affected canopy functional diversity. Specifically, there was a relatively significant direct effect of aspect on FEve (β = -0.420, Plt;0.01) and RAOQ (β = -0.300, Plt;0.05) indices for canopy trees. Soil factors mainly affected the functional diversity of different forest layers through species diversity. Soil total phosphorus, available phosphorus, alkaline hydrolysis nitrogen, and available potassium content" had a greater effect on the functional diversity of the understory, whereas soil total potassium" content" affected the FRic and RAOQ indices of the canopy. 【Conclusion】 There were marked differences in the characteristics of functional diversity at different forest stratum in the study area. Functional diversity was mainly affected by species diversity, canopy openness, and soil nutrients of understory plants, whereas topographic factors determined the functional diversity of the canopy layer. This study reveals the effects of biological and abiotic environmental factors on functional diversity across different forest layers and expands the understanding of the diversity of vertical stratification. The findings provide a further reference for biodiversity protection in natural forests.
Keywords:old-growth forest; canopy; understory; functional diversity;" structural equation model (SEM)
维持和保护生物多样性是生态学研究长期关注的热点问题之一[1]。传统的生物多样性评估以物种多样性为代表,将所有物种视为彼此等同或不同,来研究生物多样性与生态系统功能之间的关系[2]。许多研究表明,物种丰富度越高的森林群落,生态系统功能更稳定[3]。然而,在物种数量相同的两个群落中,由于物种功能特性的相似或相异程度不同,生态系统功能发挥的作用可能会存在较大的差异[4]。作为物种差异程度的拓展,植物功能性状是植物在进化和生长过程中与外部环境相互作用的结果[5]。植物功能性状多样性(以下简称功能多样性)充分考虑了物种数量、多度及性状差异,它可以通过互补效应或功能冗余影响生态系统过程,从而促进生物多样性的形成[6-8]。因此,相比物种多样性,功能多样性可以更直接、全面、准确地预测生态系统功能。通过量化和深入理解功能多样性,可以制定更合理的生物多样性保护策略。
目前,针对功能多样性的量化主要从丰富度、均匀度与分离度3个相互独立的层面表达,应用较广泛的指标有功能丰富度指数(FRic)、功能均匀度指数(FEve)、功能分歧度指数(FDiv)和功能二次熵指数(RAOQ)等[9]。许多学者采用这些功能多样性指数对植物群落功能多样性的影响因素展开探讨,其探讨的内容主要分为两个方面:一方面,由于在计算功能多样性指数的过程中考虑了物种及其多度分布特征,物种多样性与功能多样性存在直接的关系,但受环境异质性、空间尺度或干扰强度等影响,物种多样性与功能多样性之间的关系未达成一致[10-12];另一方面,功能多样性受到不同环境因素的直接影响,即环境因子充当过滤器的作用,从性状水平影响植物的定植、存活和生长,海拔梯度、坡向、土壤磷含量及岩石裸露率等生境异质性均会导致功能多样性的差异[13-15]。由此可见,物种多样性、环境因子和功能多样性三者之间存在紧密的关联,物种多样性对功能多样性有直接的影响,而环境因子既可以直接影响也可以通过物种多样性间接影响群落的功能多样性。
垂直分层现象是森林群落结构的基本特征之一,生态位分化塑造了垂直分层多样性特征并促进了森林群落的物种共存[16-17]。已有研究表明,分层结构改变了林层内部的光照、温度和湿度等微环境,使得林下资源分配变得更复杂,植物的种类、数量和分布受到影响[18]。这种分层现象也导致了植物功能性状在不同垂直林层表现出明显的分异[19]。以往关于不同生活型植物的功能多样性研究发现,乔木层与灌木层植物功能多样性的主要影响因素不同[20-21]。然而,目前关于环境与森林植被功能多样性之间的关系主要集中于林分整体,基于垂直分层结构的功能多样性特征及其影响因素的研究鲜见报道。
常绿阔叶天然林是我国亚热带地区最具代表性的森林植被类型之一[22]。其中,处于演替后期的老龄林垂直结构明显,林分内光异质性强,生态系统更稳定[23]。研究其群落功能多样性对维持森林生态系统功能稳定性具有重要参考价值。基于上述背景,本研究以广西雅长自然保护区1.6 hm2常绿阔叶天然老龄林监测样地的植被调查数据为基础,通过采集植物叶片性状和环境因子(林下光照、地形、土壤)数据,拟探讨以下科学问题:①功能多样性在垂直层次上有何变化规律?②不同林层内,物种多样性与环境因子对功能多样性的影响有何侧重?对此问题的科学回答有助于更好地理解植物群落构建过程,为维持森林群落生物多样性提供科学参考依据。
1 材料与方法
1.1 研究区概况与样地设置
本研究区位于广西雅长兰科植物国家级自然保护区(106°11′31″~106°27′04″E,24°44′16″~24°43′58″N),该保护区总面积为22 062 hm2,地跨田林、乐业两县。保护区内有北盘江、南盘江和红水河等河流,地理位置独特,生态区位的重要性尤为突出。该区域属中亚热带季风气候,年均气温为16.3 ℃,年均相对湿度82%,年均降水量为1 051.7 mm。土壤类型包括褐红土、红壤和黄壤,植被类型主要为南亚热带常绿阔叶林[24]。
2020年7月,用全站仪在广西国有雅长林场九龙分场内建立了一块面积为1.6 hm2 (200 m×80 m)的固定样地,划分为40个样方(20 m×20 m)。样地基准点地理坐标为106°20′25″E、24°38′54″N,海拔1 700 m。对样地内胸径≥1 cm的木本植物进行调查,内容包括记录每一个木本植物个体的种类、胸径、树髙、坐标等,并挂牌标记。样地内现有木本植物33科,55属,84种(APG Ⅳ,被子植物系统发育研究组分类系统),10 077株。主要优势树种有:多花杜鹃(Rhododendron cavaleriei)、凹脉柃(Eurya impressinervis)、细齿柃(E. nitida)、榕叶冬青(Ilex ficoidea)、枫香(Liquidambar formosana)等。
1.2 植物功能性状的样品采集与测定
2021年8月,参照全球植物功能性状测量标准手册[25],采集并测量了76个确定树种的叶片性状。其中,常见种采集10株以上,少于10株的稀有种采集3~5株。本研究选取6个植物功能性状:叶干物质含量(leaf dry matter content, LDMC)、叶绿素相对含量(relative chlorophyll content, 以SPAD值表示)、叶片全磷含量(leaf phosphorus content, LPC)、叶片全氮含量(leaf nitrogen content, LNC)、比叶面积(specific leaf area, SLA)和最大树高(Hmax)。其中,SPAD、SLA、LNC和LPC为资源获取型性状;LDMC属于资源保守型性状;Hmax属于优势功能性状[26]。
采用SPAD-502叶绿素计(柯尼卡美能达,中国)测定植物叶绿素相对含量[27],测量叶片鲜质量,叶面积用扫描仪(EPSON,中国)扫描叶片原始图像,并用ImageJ 18.0软件计算图像中的叶面积[28]。将扫描后的叶片置于80 ℃烘箱中烘48 h至质量恒定,测量记录叶片干质量,从而获取比叶面积和叶干物质含量数据[25]。烘干后的叶片用球磨仪研磨至粉末状,用靛酚蓝比色法测量叶片全氮含量,钼锑抗比色法测量叶片全磷含量[29]。用Haglf(瑞典)超声波测高测距仪测量样地林木树高,最大树高查阅地方植物志并结合实际样地树高确定。
1.3 环境因子的测定
1.3.1 地形因子
本研究选取了3个地形因子:坡度(slope)、坡向(aspect)、凹凸度(convexity)。基于Harms等[30]的计算方法在Arcgis10.2中计算每个20 m×20 m样方的地形因子。具体计算原理为:将4个顶点的海拔平均值视为该样方的海拔高度;目标样方的海拔减去与之相邻8个样方的海拔平均值为该样方的凹凸度;分别连接每个样方的3个顶点形成4个三角形平面,每个平面与该样方投影夹角的平均值为目标样方的坡度;这4个三角形平面与正北方向的角度平均值为目标样方的坡向。由于海拔变异性不大,该研究不考虑海拔的影响。
1.3.2 土壤和光照因子
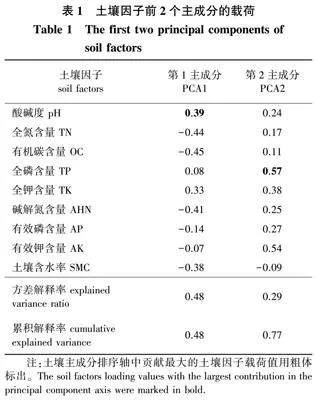
2021年8月,使用土钻在样方中5点取样(沿对角线和样方中心点),深度为0~20 cm,样品混合均匀后装袋带回实验室。用烘干法及时测定土壤含水率;酸碱度用电位法测定;全氮含量用流动分析仪测定;全磷含量用钼锑抗比色法测定;全钾和有效钾含量用火焰光度计法测定;有机碳含量用重铬酸钾容量法测定;碱解氮含量用碱解扩散法测定;有效磷含量用钼锑抗比色法测定[29]。为了减少土壤变量的数量,本研究将所有土壤理化性质数据标准化(平均值为0,标准差为1),以统一不同的测量单位,然后对标准化的数据进行了主成分分析(表1)。使用ImageJ 18.0软件计算林冠开度(canopy openness,Co)[31]。每个小样方的林下光照强度为样方内5点林冠开度的平均值,具体过程为:选择阴天或晴天的日出或日落时刻,将EOS 80D单反相机外接Sigma 4.5 mm F2.8EXDC鱼眼镜头(日本)转换器用脚架水平放置于离地面2 m处,调整脚架使顶部磁针与正北方向重合,在每个样方四角和对角线交点处镜头朝上拍摄半球面林冠影像3张,共计拍摄600张半球面林冠影像。
1.4 数据处理
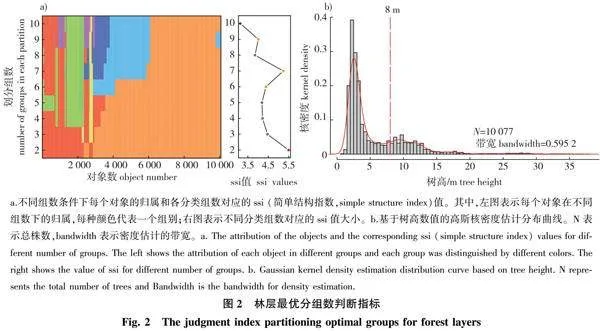
基于林木树高,用K-means聚类的方法将样地内胸径≥1 cm的林木个体进行分层,参照简单结构指数(simple structure index,“ssi”值)获得垂直结构层次和各层树高范围[17]。用高斯核密度估计法绘制树高密度分布曲线以检验最佳分层数[32]。为探究不同林层内功能多样性的主要影响因素,本研究针对不同林层的功能多样性指数构建结构方程模型,其概念模型见图1。本研究物种多样性采用物种丰富度(SR)、香农指数(Shannon)、辛普森指数(Simpson)和均匀度指数(Pielou)[33];功能多样性采用功能丰富度(FRic)、功能均匀度(FEve)、功能分歧度(FDiv)和二次熵指数(RAOQ)[10,34]。
由于结构方程模型分析受样本量与路径系数关系的限制,且变量之间可能存在高度共线性,因此需对解释变量进行筛选[33]。首先,为确定解释变量的重要程度,进行随机森林分析。依据随机森林模型中的均方误差增量(increase in mean squared error,记为%IncMSE值)分别对各林层功能多样性的影响因子进行排序。然后,对所有影响因素做相关性分析。以随机森林分析得到的排序列出各影响因子的相关性系数矩阵,若变量之间的相关系数大于或等于0.7,则排序靠前的变量保留,排序靠后的变量被排除,最终选择未被排除的所有变量作为结构方程模型的起始变量[7]。最后,参照模型修正指数(modification indices,mi值)、卡方检验结果、路径系数显著性和路径关系的实际意义等对结构方程模型进行模型修正[35]。本研究结构方程模型的运行基于R语言中的“lavaan”包[36],采用标准化路径系数β度量变量之间的关系,并通过卡方检验(χ, Chi-square, Pgt;0.05)、赤池信息准则(akaike information criterion, AIC)、近似误差平方根(root mean square error of approximation, RMSEA)、相对适配指数(comparative fit index, CFI)和标准化均方根残差(standardized root mean square residual, SRMR)对整个模型进行评价。另外,相关系数的计算调用“ggcorrplot”包[37],随机森林分析调用“RandomForest”包[38],ssi值和物种多样性指数的计算使用“vegan”包[39],功能多样性指数的计算使用“FD”包[40]。所有计算过程均在R 4.1.1[41]中完成。
2 结果与分析
2.1 物种多样性与功能多样性的分层特征
由聚类分析和高斯核密度估计可知,ssi值在分类组数为2时最大,树高密度曲线有2个“峰”(图2),确定最优组数为2。将最佳分类组数代入K-means聚类法中,得到林层从下到上的树高范围依次为[0.6,8.0)、[8.0,37.6) m。林下层有木本植株7 528株,隶属于30科47属69种(APG Ⅳ,下同);特有种22种,占总物种数的31.88%。林冠层有木本植株2 549株,隶属于26科41属59种;特有种12种,占总物种数的20.34%。
对于大部分多样性指数而言,林分整体显著高于分层,并且林下层显著高于林冠层(Plt;0.05)。其中,林分整体的SR、Shannon、Simpson、FRic指数显著大于分层,而林分FEve、FDiv指数分别与冠层和下层无显著差异,林分Pielou、RAOQ指数均与下层无显著差异;林下层SR、Shannon、Simpson、FRic、FEve、RAOQ指数显著大于林冠层,但林下层FDiv指数显著低于林冠层,林下层Pielou指数则与林冠层无显著差异(表2)。
2.2 物种多样性与环境因子对不同林层功能多样性的影响
由随机森林模型确定的各影响因子重要性排序图可以看出(图3),对林下层功能多样性指数而言,物种多样性指数的影响更大;而对林冠层功能多样性指数而言,环境因子的影响更大。具体来说,林下层FRic、FEve、RAOQ指数的影响因子重要性排序中,物种多样性指数的重要性值均大于环境因子;林冠层FEve、FDiv、RAOQ影响因子的重要性排序中,部分环境因子(坡向、凹凸度或土壤因子)的重要性值大于物种多样性指数。
变量的重要程度由均方误差增量(%IncMSE)表示,每一个值表示去掉该变量后模型预测误差的增量。The variable importance values are expressed by the percentage increase of mean square incresement (%IncMSE). Simpson.辛普森指数Simpson index;Shannon.香农指数Shannon index;Pielou.均匀度指数Pielou index; PCA1.土壤因子主成分排序轴1Each value represents the increase in the prediction error of the same model after the omission of a variable the first principal component axis for soil factors;PCA2.土壤因子主成分排序轴2 the second principal component axis for soil factors;Co.林冠开度canopy openness;Slope.坡度;Aspect.坡向;Convexity.凹凸度。下同。The same below.
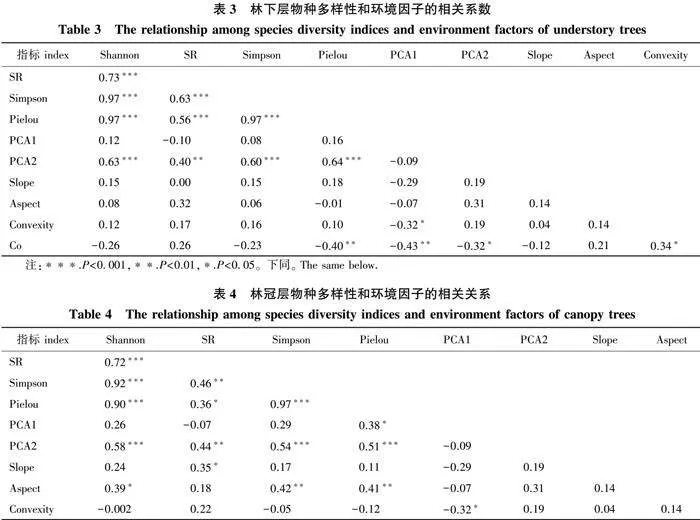
各林层Shannon指数与SR、Simpson、Pielou指数的相关系数均大于0.7,且Simpson指数与Pielou指数的相关系数也大于0.7(表3、表4),在结构方程模型中均按随机森林模型重要性排序的顺序依次剔除。
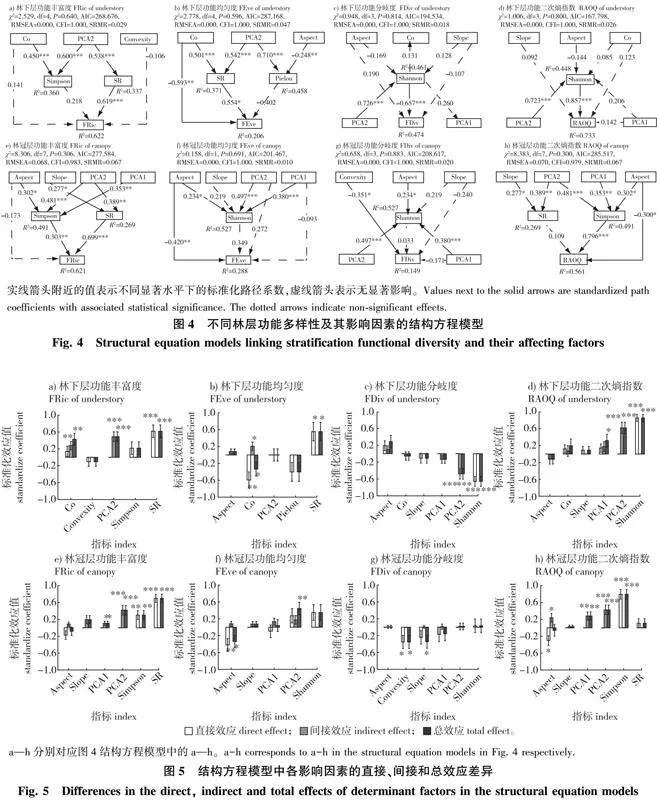
由功能多样性的结构方程模型可知,对于功能丰富度FRic指数而言,各林层均受SR指数和土壤因子PCA2的影响。在林下层,SR直接影响FRic(β = 0.619,Plt;0.001);林冠开度(Co)通过SR对FRic有较显著的间接正效应;土壤因子PCA2通过SR对FRic有显著的间接正效应;各因子总体解释度为0.622(图4a、5a)。在林冠层,SR和Simpson指数直接影响FRic(β = 0.699,Plt;0.001;β = 0.303,Plt;0.01);土壤因子(PCA1、PCA2)对FRic有显著的间接正效应;各因子总体解释度为0.621(图4e、5e)。
对于功能均匀度FEve指数而言,林下层主要受到SR指数和林冠开度Co的影响,而林冠层受坡向和土壤因子PCA2的影响较大。在林下层,SR和Co直接影响FEve(β = 0.554,Plt;0.05;β = -0.593,Plt;0.01);同时,Co通过SR对FEve有较显著的间接正效应,但总体上Co对FEve有较显著的负效应;各因子总体解释度为0.206(图4b、5b)。在林冠层,坡向直接影响FEve(β = -0.420,Plt;0.01);土壤因子PCA2对FEve有较显著的总体正效应;各因子总体解释度为0.288(图4f、5f)。
对于功能分歧度FDiv指数而言,林下层主要受到Shannon指数和土壤因子PCA2的影响,而林冠层受凹凸度和坡度的影响较大。在林下层,Shannon直接影响FDiv(β = -0.657,Plt;0.001),土壤因子PCA2通过Shannon对FDiv有显著的间接负效应;各因子总体解释度为0.474(图4c、5c)。在林冠层,凹凸度直接影响FDiv(β = -0.351,Plt;0.05),坡度对FDiv有较显著的总体负效应;各因子总体解释度为0.149(图4g、5g)。
对于功能二次熵RAOQ指数而言,林下层主要受到Shannon指数和土壤因子PCA2的影响,而林冠层主要受到Simpson指数和土壤因子(PCA1、PCA2)的影响。在林下层,Shannon直接影响RAOQ(β = 0.857,Plt;0.001);土壤因子(PCA2)通过Shannon对RAOQ有显著的间接正效应;各因子总体解释度为0.733(图4d、5d)。在林冠层,Simpson和坡向直接影响RAOQ(β = 0.796,Plt;0.001;β = -0.3,Plt;0.01);土壤因子(PCA1、PCA2)对RAOQ有较显著的间接正效应;各因子总体解释度为0.561(图4h、5h)。
3 讨 论
3.1 物种多样性与功能多样性在林层间的变化规律
本研究林下层物种多样性SR、Simpson与Shannon指数显著高于林冠层。其主要原因可能是该森林群落林下层不仅涵盖了少部分大径级优势树种的小树,还存在部分较多的林下特有种,且个体数与物种数远大于林冠层。另外,由于该样地林分整体或林下层的物种多度占比更均匀,而林冠层各物种多度占比相差较大,因此,Pielou指数在林下层与林冠层间无显著差异,而林冠层显著小于林分整体。不同林层物种多样性的变化规律与桂旭君等[17]在鼎湖山南亚热带常绿阔叶林群落研究的结果基本一致。
功能多样性的变化规律可能与每个功能多样性指数所表达的含义不同有关。功能丰富度FRic指数表示群落中物种所占的性状生态位空间[10]。本研究结果表明,林下层FRic指数显著高于林冠层,意味着林下层物种所占据的生态位空间大于林冠层。一般而言,随着林分的发展,林下光照异质性与土壤腐殖质逐渐形成,促进了林下物种多样性的提高,从而扩展了物种所占据的性状生态位空间;但林冠层优势树种丰富度较低且功能性状受到了较强的环境过滤作用,因而物种所占的生态位空间相对较小[19,42]。功能均匀度FEve指数可以理解为生态位空间中功能性状分布的均匀程度,若存在间隙分布,则会导致群落稳定性减弱[9,15]。本研究林下层FEve指数显著大于林冠层,表明该群落林下层物种及其多度在性状空间的分布相对较均匀,林下资源利用较充分。功能二次熵RAOQ指数是基于物种丰度权重的物种对间的距离之和,反映了成对物种之间的性状差异[34]。若性状差异较大,则群落可能受到相似性限制(竞争排斥)的影响;若性状差异较小,则群落性状受到生境过滤的作用较强,因此该指数具有较好的群落构建机制检测效果[42-43]。本研究结果显示,林下层RAOQ指数显著大于林冠层,暗示了林下层物种间的性状差异大于林冠层。这可能是因为在森林群落中,小树存在较强的竞争排斥作用,而大树是生境过滤的结果[44]。功能分歧度FDiv指数表示生态位空间中物种多度分布的离散程度,同时也体现了群落中物种间生态位的互补程度,若群落功能分歧度越高,生态位分化程度也越高,随之资源竞争程度更低[6,15]。本研究样地中,所有林层的平均FDiv指数接近最大阈值“1”,表明该群落生态位分化程度较高;但林下层FDiv指数显著低于林冠层,意味着林下层物种的生态位重叠效应更高,资源竞争可能相对更强。
3.2 林下层功能多样性与物种多样性及环境因子的关系
林下层物种丰富度SR指数越高,功能丰富度FRic指数越大,同时,林冠开度的增加促进了下层SR指数的提高,进而增加了功能丰富度。这可能是因为林冠开度反映了林下光资源可用性,越大的林冠开度能够满足更多物种的光需求[45]。一方面,林下光照增强会抑制林下耐阴草本与灌木层植被生存,但也促进了更多喜阳物种的共存[46]。本研究样地林下层存在较多优势树种的小树,如多花杜鹃、凹脉柃等。因此,林冠开度越大,越有利于林下层喜阳树种的存活,物种多样性与功能多样性随之增加。另一方面,适度的光照能够促进林下幼树的更新与存活[47]。本研究物种丰富度SR指数对林下功能均匀度FEve指数有较显著的正向影响,其原因可能是林下层物种数的增加弥补了性状生态位空间中物种频率分布的差异,从而促进了生态位空间中资源的利用效率[48]。对于RAOQ和FDiv指数而言,Shannon指数对林下功能分歧度FDiv指数有显著的负向影响,而Shannon指数对林下RAOQ二次熵指数有显著的正向影响)。一般来说,在功能性状生态位空间中,常见种占据生态位空间中心位置,而稀有种更倾向于占据生态位空间的边缘位置,同时稀有种的变化与Shannon指数呈正相关[22,49]。本研究由于林下层稀有种的数量更多,因而Shannon指数增大,生态位中心常见种多度分布的距离相对更近,FDiv指数变小。而RAOQ指数受物种多度权重的影响更大[34]。当Shannon指数增大时,稀有种多度权重变大,RAOQ指数也增大。由此可知,林下层FDiv指数与RAOQ指数从不同角度反映了功能性状的分离程度。
此外,土壤养分有效性是影响亚热带林下物种多样性的主要因素[42]。本研究中,土壤因子PCA轴2对所有林下层物种多样性指数(SR、Simpson、Shannon、Pielou)均存在正效应。这可能是因为该排序轴与多数有效性土壤养分(AHN、AP、AK)正相关。同时,许多研究证明土壤磷含量是林下物种多样性的主要限制因子,土壤磷含量较低的情况下,树种会发生生态位分化[46,50]。本研究结果与大多数研究一致,该样地土壤因子PCA轴2中,全磷的贡献最大且该林分平均土壤全磷含量较低,物种对土壤磷的充分利用促进了生态位分化,从而提高物种多样性。另一方面,由于土壤的酸碱度会制约土壤元素的有效性,且土壤钾含量也是影响植物生长的限制性元素之一,因此,较低的pH或钾含量可能会限制土壤养分的可用性[51]。本研究中,土壤因子PCA轴1主要反映了土壤pH和土壤全钾,且该样地土壤偏酸性,土壤全钾含量均值为12.32 g/kg,低于中国南方典型常绿阔叶林土壤全钾含量均值31.85 g/kg[46],因而土壤因子PCA轴1对林下物种多样性的影响不大。综上可知,林下层物种多样性与功能多样性的关系主要通过环境因子来调节。
3.3 林冠层功能多样性与物种多样性及环境因子的关系
林冠层功能丰富度的影响因素与林下层基本一致,SR与Simpson指数对冠层功能丰富度FRic指数有显著的正向影响。然而,Shannon指数对冠层均匀度FEve指数与功能分歧度FDiv指数无显著影响,坡向对FEve有较显著的负向影响,同时凹凸度对FDiv指数也有较显著的负效应。这可能是因为FEve和FDiv指数均与生态位空间中的物种多度分布情况有关,由于冠层林木优势物种数较少,生态位空间较窄,物种多度在生态位空间中分布的格局变化不明显[6]。Biswas等[11]研究发现,在物种数较少的情况下,生境因素对功能离散程度的影响更大,这与本研究中林冠层FEve和FDiv指数的研究结果相似。对于RAOQ指数来说,其受Simpson指数的影响最大,Simpson指数反映了物种多度分配的均匀程度,在RAOQ指数的计算中影响到物种权重;其次,RAOQ指数受坡向的负向影响,坡向差异可能会导致冠层植物太阳辐射量接收程度不同,从而影响植物功能性状的表型可塑性[15]。由此可见,林冠层RAOQ指数可能主要取决于物种多度及坡向变化导致的性状差异。此外,本研究土壤PCA 2对林冠层物种多样性的影响小于林下层,但土壤PCA1对林冠层物种多样性的影响大于林下层,这可能是因为土壤钾含量对大径级乔木的生境过滤作用较强[52]。总的来说,林冠层物种多样性与功能丰富度FRic和功能二次熵RAOQ紧密相关,但物种多样性在解释林冠层功能均匀度FEve和功能分歧度FDiv时并没有发挥太大作用,而地形因子是直接影响冠层功能多样性的重要环境因素。
综上所述,雅长常绿阔叶老龄林存在明显的垂直分层特征,林下层比林冠层具有更高的功能多样性,是整个群落功能多样性的重要组成部分。同时,林下层包含了更多的稀有种与常见种,其功能多样性受物种多样性的影响较大。由于林下微生境复杂,异质性较高,资源竞争激烈,因此下层功能多样性受到林下光照强度与土壤养分的直接或间接影响。而林冠层只有少数优势物种,部分冠层功能多样性指数与物种多样性的关系较弱,因为冠层植物与太阳辐射接触面更大,地形变化引起的太阳辐射量差异可能对其造成直接的影响。本研究揭示了亚热带天然老龄林在垂直层次上物种多样性与功能多样性的变化规律以及垂直分层内部功能多样性的驱动机制,拓展了以往关于垂直分层多样性的认识,为天然林生物多样性保护提供更进一步的参考依据。
参考文献(reference):
[1]马克平.生物多样性科学的热点问题[J].生物多样性,2016,24(1):1-2.MA K P. Hot topics for biodiversity science[J].Biodivers Sci,2016,24(1):1-2.DOI: 10.17520/biods.2016029.
[2]MORI A S,FURUKAWA T,SASAKI T.Response diversity determines the resilience of ecosystems to environmental change[J].Biol Rev,2013,88(2):349-364.DOI: 10.1111/brv.12004.
[3]XU W,LUO W X,ZHANG C Y,et al.Biodiversity-ecosystem functioning relationships of overstorey versus understorey trees in an old-growth temperate forest[J].Ann For Sci,2019,76(3):1-13.DOI: 10.1007/s13595-019-0845-8.
[4]CHAO A,CHIU C H,JOST L.Unifying species diversity,phylogenetic diversity,functional diversity,and related similarity and differentiation measures through hill numbers[J].Annu Rev Ecol Evol Syst,2014,45:297-324.DOI: 10.1146/annurev-ecolsys-120213-091540.
[5]何芸雨,郭水良,王喆.植物功能性状权衡关系的研究进展[J].植物生态学报,2019,43(12):1021-1035.HE Y Y,GUO S L,WANG Z.Research progress of trade-off relationships of plant functional traits[J].Chin J Plant Ecol,2019,43(12):1021-1035.DOI: 10.17521/cjpe.2019.0122.
[6]MASON N W H,MOUILLOT D,LEE W G,et al.Functional richness,functional evenness and functional divergence:the primary components of functional diversity[J].Oikos,2005,111(1):112-118.DOI: 10.1111/j.0030-1299.2005.13886.x.
[7]LUO Y H,CADOTTE M W,BURGESS K S,et al.Greater than the sum of the parts:how the species composition in different forest strata influence ecosystem function[J].Ecol Lett,2019,22(9):1449-1461.DOI: 10.1111/ele.13330.
[8]AGUIRRE-GUTIRREZ J,BERENGUER E,MENOR I O,et al.Functional susceptibility of tropical forests to climate change[J].Nat Ecol Evol,2022,6(7):878-889.DOI: 10.1038/s41559-022-01747-6.
[9]韩涛涛,唐玄,任海,等.群落/生态系统功能多样性研究方法及展望[J].生态学报,2021,41(8):3286-3295.HAN T T,TANG X,REN H,et al.Community/ecosystem functional diversity:measurements and development[J].Acta Ecol Sin,2021,41(8):3286-3295.DOI: 10.5846/stxb201903080442.
[10]VILLGER S,MASON N W H,MOUILLOT D.New multidimensional functional diversity indices for a multifaceted framework in functional ecology[J].Ecology,2008,89(8):2290-2301.DOI: 10.1890/07-1206.1.
[11]BISWAS S R,MALLIK A U,BRAITHWAITE N T,et al.Effects of disturbance type and microhabitat on species and functional diversity relationship in stream-bank plant communities[J].For Ecol Manag,2019,432:812-822.DOI: 10.1016/j.foreco.2018.10.021.
[12]SUREZ-CASTRO A F,RAYMUNDO M,BIMLER M,et al.Using multi-scale spatially explicit frameworks to understand the relationship between functional diversity and species richness[J].Ecography,2022(6):e05844.DOI: 10.1111/ecog.05844.
[13]许驭丹,董世魁,李帅,等.植物群落构建的生态过滤机制研究进展[J].生态学报,2019,39(7):2267-2281.XU Y D,DONG S K,LI S,et al.Research progress on ecological filtering mechanisms for plant community assembly[J].Acta Ecol Sin,2019,39(7):2267-2281.DOI: 10.5846/stxb201804260946.
[14]DING Y,ZANG R G,LU X H,et al.The effect of environmental filtering on variation in functional diversity along a tropical elevational gradient[J].J Veg Sci,2019,30(5):973-983.DOI: 10.1111/jvs.12786.
[15]盘远方,李娇凤,姚玉萍,等.桂林岩溶石山青冈群落植物功能多样性和环境因子与坡向的关联研究[J].生态学报,2021,41(11):4484-4492.PAN Y F,LI J F,YAO Y P,et al.Changes in plant functional diversity and environmental factors of Cyclobalanopsis glauca community in response to slope gradient in Karst hills,Guilin[J].Acta Ecol Sin,2021,41(11):4484-4492.DOI: 10.5846/stxb201906031169.
[16]李杰, 李远发, 陆道调, 等. 南盘江流域松栎混交林的分层多样性特征[J/OL].生态学杂志:1-13[2022-06-21]. LI J, LI Y F, LU D D, et al. The characteristics of stratification diversity of pine-oak mixed forests in the Nanpan River Basin[J/OL]. Chinese Journal of Ecology: 1-13[2022-06-21]. http://kns.cnki.net/kcms/detail/21.1148.Q.20220620.1345.014.html.
[17]桂旭君,练琚愉,张入匀,等.鼎湖山南亚热带常绿阔叶林群落垂直结构及其物种多样性特征[J].生物多样性,2019,27(6):619-629.GUI X J,LIAN J Y,ZHANG R Y,et al.Vertical structure and its biodiversity in a subtropical evergreen broad-leaved forest at Dinghushan in Guangdong Province,China[J].Biodivers Sci,2019,27(6):619-629.DOI: 10.17520/biods.2019107.
[18]LI Y,HE J,YU S,et al.Spatial structure of the vertical layers in a subtropical secondary forest 57 years after clear-cutting[J].iForest,2019,12(5):442-450.DOI: 10.3832/ifor2975-012.
[19]李艳朋,倪云龙,许涵,等.鼎湖山南亚热带常绿阔叶林植物功能性状变异与不同垂直层次个体生长的关联[J].生物多样性,2021,29(9):1186-1197.LI Y P,NI Y L,XU H,et al.Relationship between variation of plant functional traits and individual growth at different vertical layers in a subtropical evergreen broad-leaved forest of Dinghushan[J].Biodivers Sci,2021,29(9):1186-1197.DOI: 10.17520/biods.2021205.
[20]吴昊,肖楠楠,林婷婷.秦岭松栎林功能多样性与物种多样性和环境异质性的耦合关系[J].生态环境学报,2020,29(6):1090-1100.WU H,XIAO N N,LIN T T.Relationships between functional diversity and species diversity of pine-oak mixed forest in Qinling Mountains and their environmental explanations[J].Ecol Environ Sci,2020,29(6):1090-1100.DOI: 10.16258/j.cnki.1674-5906.2020.06. 003.
[21]CHENG X Q,TAIN P,LI Z Z,et al.Effects of environmental factors on plant functional traits across different plant life forms in a temperate forest ecosystem[J].New For,2022,53(1):125-142.DOI: 10.1007/s11056-021-09847-0.
[22]楼一恺,范忆,戴其林,等.天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系[J].生态学报,2021,41(21):8568-8577.LOU Y K,FAN Y,DAI Q L,et al.Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of Tianmu Mountain Natural Reserve[J].Acta Ecol Sin,2021,41(21):8568-8577.DOI: 10.5846/stxb202007301989.
[23]张田田,王璇,任海保,等.浙江古田山次生与老龄常绿阔叶林群落特征的比较[J].生物多样性,2019,27(10):1069-1080.ZHANG T T,WANG X,REN H B,et al.A comparative study on the community characteristics of secondary and old-growth evergreen broad-leaved forests in Gutianshan,Zhejiang Province[J].Biodivers Sci,2019,27(10):1069-1080.DOI: 10.17520/biods.2019059.
[24]李述万.广西雅长兰科植物国家级自然保护区维管束植物物种多样性研究[D].桂林:广西师范大学,2017.LI S W.Studies on species diversity of vascular plants in Yachang orchid national nature reserve of Guangxi[D].Guilin:Guangxi Normal University, 2017.
[25]PEREZ-HARGUINDEGUY N,DIAZ S,GARNIER E,et al.New handbook for standardised measurement of plant functional traits worldwide[J].Aust J Bot,2013,61(67-234).DOI:http://dx.doi,org/10.1017/BT12225.
[26]WANG L Q,ALI A.Climate regulates the functional traits-aboveground biomass relationships at a community-level in forests:a global meta-analysis[J].Sci Total Environ,2021,761:143238.DOI: 10.1016/j.scitotenv.2020.143238.
[27]柯娴氡,贺立静,苏志尧.南方4种木本植物相对叶绿素指标及其分布[J].中南林业科技大学学报,2010,30(8):82-86.KE X D,HE L J,SU Z Y.Relative chlorophyll content and its distribution of four woody species in south China[J].J Central South Univ For amp; Technol,2010,30(8):82-86.DOI: 10.14067/j.cnki.1673-923x.2010.08.025.
[28]熊映杰,于果,魏凯璐,等.天童山阔叶木本植物叶片大小与叶脉密度及单位叶脉长度细胞壁干质量的关系[J].植物生态学报,2022,46(2):136-147.XIONG Y J,YU G,WEI K L,et al.Relationships between lamina size,vein density and vein cell wall dry mass per unit vein length of broad-leaved woody species in Tiantong Mountain,southeastern China[J].Chin J Plant Ecol,2022,46(2):136-147.DOI: 10.17521/cjpe.2021.0060.
[29]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:1-336.LU R K.Methods of soil agrochemical analysis[M].China Agriculture Scientech Press,2000:1-336.
[30]HARMS K E,CONDIT R,HUBBELL S P,et al.Habitat associations of trees and shrubs in a 50-ha neotropical forest plot[J].J Ecol,2001,89(6):947-959.DOI: 10.1111/j.1365-2745.2001.00615.x.
[31]濮毅涵,徐丹丹,王浩斌.基于数码相片的林冠郁闭度提取方法研究[J].林业资源管理,2020(6):153-160.PU Y H,XU D D,WANG H B.An approach on estimating canopy closure via digital images[J].For Resour Manag,2020(6):153-160.DOI: 10.13466/j.cnki.lyzygl.2020.06.024.
[32]唐丽丽,陈国平,冯小梅,等.基于系统发育的燕山东麓植物群落的构建机制[J].植物研究,2017,37(6):807-815.TANG L L,CHEN G P,FENG X M,et al.Community assembly rules of the east of Yanshan Mountain based on phylogeny[J].Bull Bot Res,2017,37(6):807-815.DOI: 10. 7525/j.issn.1673-5102.2017.06.002.
[33]马克平,刘玉明.生物群落多样性的测度方法 Ⅰ:α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.MA K P,LIU Y M.The measurement method of biodiversity Ι: α diversity measurement method(Ⅱ)[J].Chin Biodiversity,1994,2(4):231-239.
[34]BOTTA-DUKT Z.Raos quadratic entropy as a measure of functional diversity based on multiple traits[J].J Veg Sci,2005,16(5):533-540.DOI: 10.1111/j.1654-1103.2005.tb02393.x.
[35]石亚飞,石善恒,黄晓敏.基于R的结构方程模型在生态学中的应用[J].生态学杂志,2022,41(5):1015-1023.SHI Y F,SHI S H,HUANG X M.The application of structural equation modeling in ecology based on R[J].Chin J Ecol,2022,41(5):1015-1023.DOI: 10.13292/j.1000-4890.202203.016.
[36]ROSSEEL Y.Lavaan:an R package for structural equation modeling[J].J Stat Softw,2012,48(2):1-36.DOI: 10.18637/jss.v048.i02.
[37]KASSAMBARA A." Ggcorrplot: visualization of a vorrelation matrix using ‘ggplot2’ [EB/OL]. R package version 013.2019.[2022-08-10]. https://CRAN.R-project.org/package=ggcorrplot.
[38]BREIMAN L, CUTLER" A, LIAW" A, et al. RandomForest: Breiman and Cutlers Random Forests for Classification and Regression [EB/OL]. R package version 47-11. 2022. [2022-08-10]. https://CRAN.R-project.org/package=randomForest.
[39]OKSANEN" J, SIMPSON" G L, BLANCHET" F G, et al. Community Ecology Package [EB/OL]. R package version 26-2. 2022. [2022-08-10]. https://cran.r-project.org/web/packages/vegan/.
[40]MAMMOLA S,CARMONA C P,GUILLERME T,et al.Concepts and applications in functional diversity[J].Funct Ecol,2021,35(9):1869-1885.DOI: 10.1111/1365-2435.13882.
[41]R Core Team." R: a language and environment for statistical computing,2021. [EB/OL]. [2022-08-10]. https://www.R-project.org.
[42]SU X P,ZHENG G C,CHEN H Y H.Understory diversity are driven by resource availability rather than resource heterogeneity in subtropical forests[J].For Ecol Manag,2022,503:119781.DOI: 10.1016/j.foreco.2021.119781.
[43]BOTTA-DUKT Z,CZCZ B.Testing the ability of functional diversity indices to detect trait convergence and divergence using individual-based simulation[J].Methods Ecol Evol,2016,7(1):114-126.DOI: 10.1111/2041-210X.12450.
[44]SANAPHRE-VILLANUEVA L,DUPUY J,ANDRADE J,et al.Functional diversity of small and large trees along secondary succession in a tropical dry forest[J].Forests,2016,7(12):163.DOI: 10.3390/f7080163.
[45]郑芬,李兆佳,邱治军,等.广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响[J].生态学报,2020,40(13):4516-4527.ZHENG F,LI Z J,QIU Z J,et al.Effects of understory light on functional traits of evergreen broad-leaved forest saplings in Nanling Mountains,Guangdong Province[J].Acta Ecol Sin,2020,40(13):4516-4527.DOI: 10.5846/stxb201911152453.
[46]谭一波,申文辉,付孜,等.环境因子对桂西南蚬木林下植被物种多样性变异的解释[J].生物多样性,2019,27(9):970-983.TAN Y B,SHEN W H,FU Z,et al.Effect of environmental factors on understory species diversity in southwest Guangxi Excentrodendron tonkinense forests[J].Biodivers Sci,2019,27(9):970-983.DOI: 10.17520/biods.2019133.
[47]徐耀粘,刘检明,万丹,等.林冠结构和地形对亚热带常绿落叶阔叶林林下幼苗物种多样性和功能多样性的影响[J].植物科学学报,2020,38(6):733-742.XU Y Z,LIU J M,WAN D,et al.Effects of canopy structure and topography on seedling species diversity in an evergreen and deciduous broad-leaved mixed forest[J].Plant Sci J,2020,38(6):733-742.DOI: 10.11913/PSJ.2095-0837.2020.60733.
[48]LIANG J J,ZHOU M,TOBIN P C,et al.Biodiversity influences plant productivity through niche-efficiency[J].Proc Natl Acad Sci USA,2015,112(18):5738-5743.DOI: 10.1073/pnas.1409853112.
[49]MOUILLOT D,BELLWOOD D R,BARALOTO C,et al.Rare species support vulnerable functions in high-diversity ecosystems[J].PLoS Biol,2013,11(5):e1001569.DOI: 10.1371/journal.pbio.1001569.
[50]MAO Q G,CHEN H,GURMESA G A,et al.Negative effects of long-term phosphorus additions on understory plants in a primary tropical forest[J].Sci Total Environ,2021,798:149306.DOI: 10.1016/j.scitotenv.2021.149306.
[51]BORDIN K M,MLLER S C.Drivers of subtropical forest dynamics:the role of functional traits,forest structure and soil variables[J].J Veg Sci,2019,30(6):1164-1174.DOI: 10.1111/jvs.12811.
[52]徐武美,宋彩云,李巧明.西双版纳热带季节雨林土壤养分空间异质性对乔木树种多样性的影响[J].生态学报,2015,35(23):7756-7762.XU W M,SONG C Y,LI Q M.Relationship between soil resource heterogeneity and tree diversity in Xishuangbanna tropical seasonal rainforest,southwest China[J].Acta Ecol Sin,2015,35(23):7756-7762.DOI: 10.5846/stxb201405120968.
(责任编辑 孟苗婧 郑琰燚)
基金项目:广西科技基地和人才专项(AD20297051);国家自然科学基金项目(32101520)。
第一作者:王耀仪 (1693843668@qq.com)。
*通信作者:王宏翔 (wanghongxiang168@126.com),讲师。
猜你喜欢
中国集体经济(2017年3期)2017-02-06 15:15:01
安徽理工大学学报·社会科学版(2016年6期)2017-01-16 10:37:47
现代商贸工业(2016年24期)2017-01-13 20:03:38
现代商贸工业(2016年24期)2017-01-13 18:24:17
高教探索(2016年12期)2017-01-09 21:55:40
财经问题研究(2016年7期)2016-12-26 14:38:25
价值工程(2016年31期)2016-12-03 23:01:46
软件导刊(2016年9期)2016-11-07 17:39:03
商业经济研究(2016年14期)2016-09-14 08:03:18
商场现代化(2016年17期)2016-07-11 16:51:58
