
台湾甲腹茧蜂触角行为反应及其感受器数量观察
2024-10-09 00:00:00蔡秋旧贾静静黄伟康林珠凤冯青姚其吉训聪
热带作物学报 2024年9期
关键词:台湾甲腹茧蜂;触角;行为观察;感受器
中图分类号:S476.3 文献标志码:A
台湾甲腹茧蜂(Chelonus formosanus)属膜翅目(Hymenoptera)茧蜂科(Braconidae)甲腹茧蜂属(Chelonus)[1-2],主要分布于我国台湾[3]、广东[4]、浙江[5]、海南[6]等地。该蜂为多种夜蛾科害虫的卵-幼虫的寄生性天敌。余姿蓉[7]通过研究发现,当甜菜夜蛾(Spodoptera exigua)、斜纹夜蛾(S. litura)和草地贪夜蛾(S. frugiperda)3 种害虫同时存在时,台湾甲腹茧蜂更偏向于寄生草地贪夜蛾卵块,且被寄生后的草地贪夜蛾幼虫的体重、体长、头宽和取食量均显著减小。表明台湾甲腹茧蜂对害虫的生物防治具有较大的利用价值。
触角是寄生蜂嗅觉感觉系统的重要组成部分,主要用于昆虫远、近距离感应和识别来自生境、寄主植物、同种或异种生物的挥发性化合物刺激的器官[8-10]。昆虫触角通常被称为“鼻子”,其主要分为柄节、梗节、鞭节3 部分[11]。每部分均分布着不同类型的感受器,每种感受器均具有其相应的功能[12]。昆虫在生命周期中会进行取食、生殖、通讯、防御等行为活动,这些行为主要依靠以触角为主导器官的嗅觉感受系发挥调控作用[13]。而一种寄生蜂的种群繁衍主要依赖寄生蜂的寿命长短和搜寻寄主的效率及其生殖力强弱[14]。观察发现台湾甲腹茧蜂在活动过程中,触角会有不同程度的残缺,从而使其找到目标物的时间不同。因此,本研究通过切除不同比例触角观察雌雄成蜂寻找蜜源和寄主的时间,及其在不同处理后对寿命长短、寄生率和子代孵化率的影响,并统计分析触角感受器数量。本研究探究了台湾甲腹茧蜂触角的嗅觉作用,深入阐明触角感受器的功能机理,表明切除触角并未直接影响其生命体征,为寄生性天敌台湾甲腹茧蜂的控害作用提供理论依据。
1 材料与方法
1.1 材料
台湾甲腹茧蜂采自海南省海口市秀英区道育村,于海南省农业科学院植物保护研究所智能人工饲养室(宁波赛福实验仪器有限公司)以草地贪夜蛾为寄主进行饲养,饲养30 代以上。饲养室环境条件为温度(26±2)℃,相对湿度为(65± 10)%,光周期12 L∶12 D。成蜂以20%蜂蜜水补充营养。
草地贪夜蛾采自海南省海口市秀英区石山镇石岩村玉米地,饲养条件同寄生蜂一致。
1.2 方法
1.2.1 台湾甲腹茧蜂触角不同切除处理行为学观察 (1)不同处理触角切除方法。收集羽化24 h内的台湾甲腹茧蜂雌雄蜂,用手轻捏其胸部,在体视解剖镜(NikonSMZ745)下,用手术刀分别切除1/3 鞭节(T)、切除2/3 鞭节(T)、切除全部鞭节(T)、切除全部触角(T),以保持完整触角为对照(CK)。每个处理的应激反应恢复时间均为12 h。
(2)雌雄蜂搜寻蜜源测定。在26 ℃条件下,用移液枪将20%蜂蜜水在培养皿中点出一个直径约为8 mm 的水滴作为蜜源,再将以上不同处理的台湾甲腹茧蜂从蜜源另一侧引入培养皿中,以600 s 为限,使用相机(佳能6D2)记录不同处理台湾甲腹茧蜂找到蜜源的时间。试验前需饥饿处理12 h,每个处理重复30 次。
(3)雌蜂搜寻寄主测定。试验条件与上述搜寻蜜源测定一致,在培养皿内一侧放入约150 粒草地贪夜蛾卵块,然后在另一侧引入经不同处理的台湾甲腹茧蜂雌蜂,使用相机(佳能6D2)记录不同处理台湾甲腹茧蜂找到草地贪夜蛾卵块的时间。每次试验持续600 s,每个处理重复30 次。
(4)雌蜂搜寻到寄主后的寄生和孵化测定。取出上述试验中已被台湾甲腹茧蜂雌蜂寄生的草地贪夜蛾卵,在室内条件下进行培育,直至卵粒全部孵化最后长成蜂茧,再分别比较不同处理台湾甲腹茧蜂雌蜂的寄生率以及寄生后子代孵化率。
(5)雌雄蜂寿命测定。分别将经过不同触角切除处理的雌雄蜂依次放入单独的试管内,然后用棉花塞住管口,以20%蜂蜜水饲养,每天统计雌雄蜂的存活情况,并补充20%蜂蜜水。
1.2.2 台湾甲腹茧蜂触角扫描电镜观察 取羽化24 h 内的雌雄成虫各30 头,分别在体视解剖镜(NikonSMZ745)下切除1/3 鞭节(T)、2/3 鞭节(T)、全部鞭节(T),以保持完整触角为对照(CK)。将切下的触角样品置于100 Hz 超声波中清洗15 min,然后在4 ℃条件下用2.5%戊二醛固定液固定24 h。固定样品用PBS 缓冲液清洗3 min,再分别用不同浓度乙醇溶液(30%、50%、70%、90%、100%)脱水,每个处理脱水20 min。样品于无尘室内自然干燥后贴于导电胶上,通过JYSC-110 喷金设备(广州竞赢科学仪器有限公司) 在真空环境下喷金, 然后使用日立台式TM4000 扫描电子显微镜观察并拍照,统计不同处理触角感受器的数量。
1.3 数据处理
使用Excel 软件统计试验数据,采用SPSS 软件分别对数据进行正态性检验和方差齐性检验,再进行显著性分析。不同处理成虫搜寻蜜源和寄主卵的数据不符合正态分布则采用非参数分析,用Mann-whitney U 进行检验;不同处理成虫搜寻蜜源和寄主卵,及其产卵寄生和孵化,以及不同处理触角感受器数量的差异则通过ANOVA 单因素方差分析,以Tukey 法进行事后两两比较;采用独立样本T 检验分析不同处理雌雄蜂寿命和触角感受器数量的差异;根据SCHNEIDER[15]报道的昆虫触角感受器对台湾甲腹茧蜂触角感受器进行分类。使用Graphpad Prism 软件绘图。
2 结果与分析
2.1 台湾甲腹茧蜂触角不同切除处理行为学
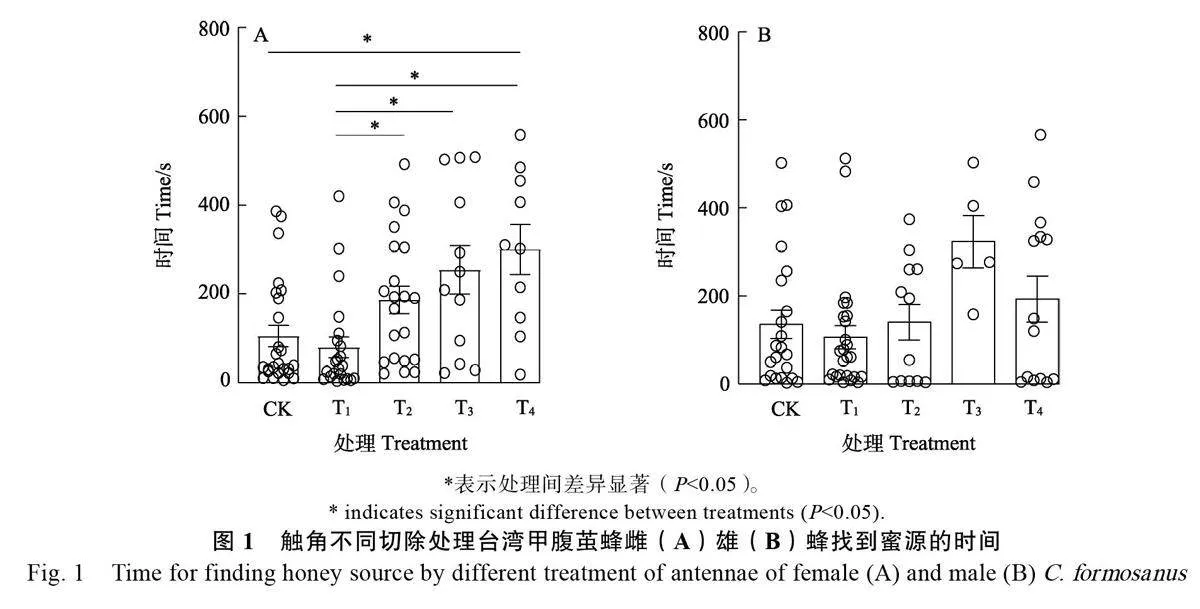
2.1.1 触角不同切除处理对雌雄蜂搜寻蜜源的影响 结果表明,台湾甲腹茧蜂触角不同切除处理显著影响雌蜂对蜜源的搜寻时间(P<0.05),其中,切除1/3 鞭节雌蜂找到蜜源的时间最短, 为(79.59±23.22)s,并且与其他处理均有显著性差异。CK 找到蜜源时间为(94.83±23.10)s,仅与切除全部触角处理(300.20±56.21)s 差异显著,与其他处理相比,切除全部触角雌蜂找到蜜源的时间最长(图1A)。
台湾甲腹茧蜂触角不同切除处理对雄蜂搜寻蜜源时间无显著影响,其中,切除1/3鞭节雄蜂找到蜜源的时间最短,为(106.52±26.71)s,该结果与雌蜂试验结果一致,但与其他处理之间无显著差异。由图1B可知,切除全部鞭节找到蜜源时间最长, 为(269.50±72.58)s,其次为切除全部触角(193.14±52.70)s。
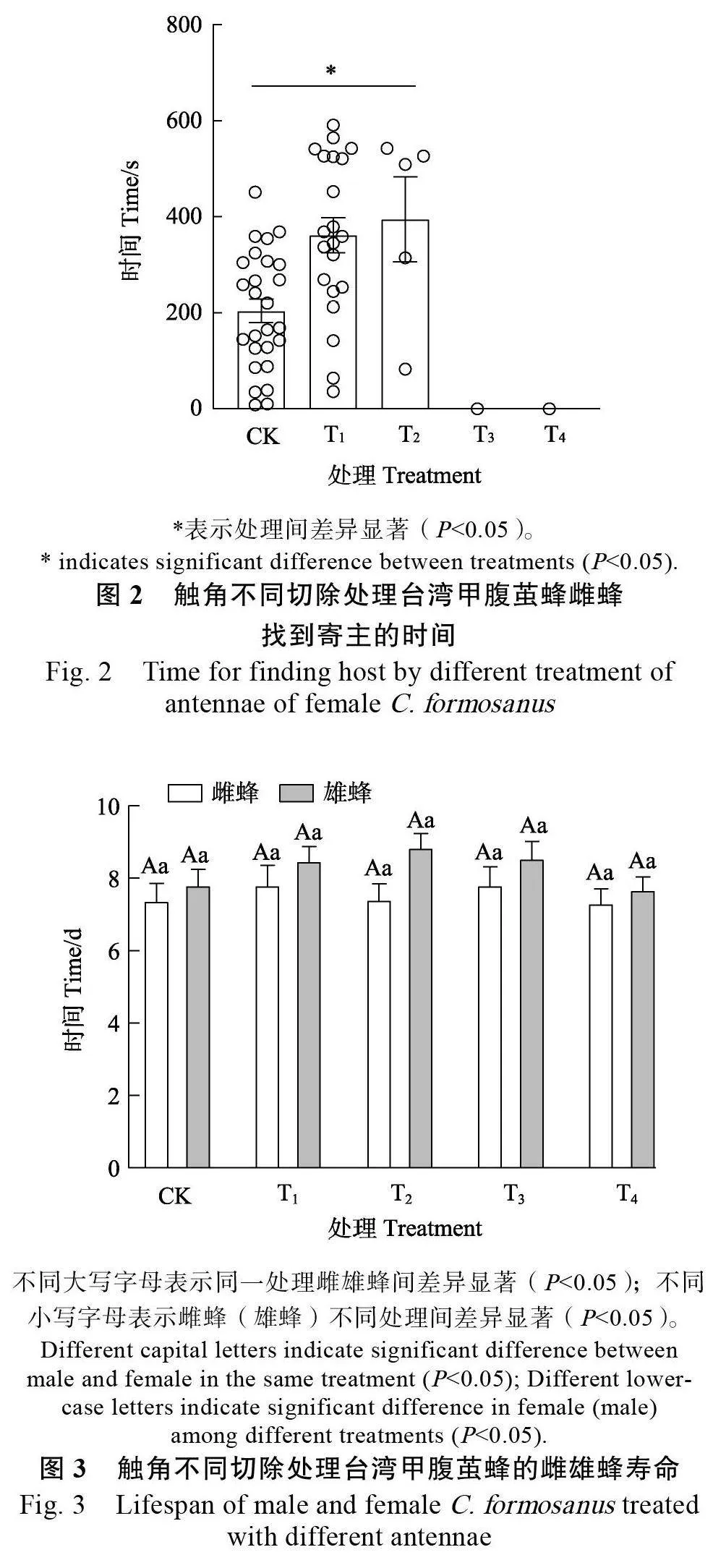
2.1.2 触角不同切除处理对雌蜂搜寻寄主的影响台湾甲腹茧蜂触角不同切除处理显著影响雌蜂对寄主卵的搜寻时间(P<0.05),CK雌蜂找到寄主卵的时间为(204.38±24.01)s,显著短于切除2/3鞭节雌蜂(394.80±88.22)s(图2)。此外,切除全部鞭节雌蜂和切除全部触角雌蜂完全找不到寄主,表明切除不同比例触角对雌蜂的寄主搜寻行为能够产生较大的影响。
2.1.3 触角不同切除处理对雌雄蜂寿命的影响台湾甲腹茧蜂雌雄成蜂的平均寿命均超过7 d,CK雄蜂的平均寿命为(7.83±0.42)d,较长于雌蜂的(7.40±0.47)d;每个处理中的雄蜂寿命均长于雌蜂,但每个处理中雌雄蜂之间均差异不显著(图3)。表明切除触角并不会直接影响台湾甲腹茧蜂寿命。
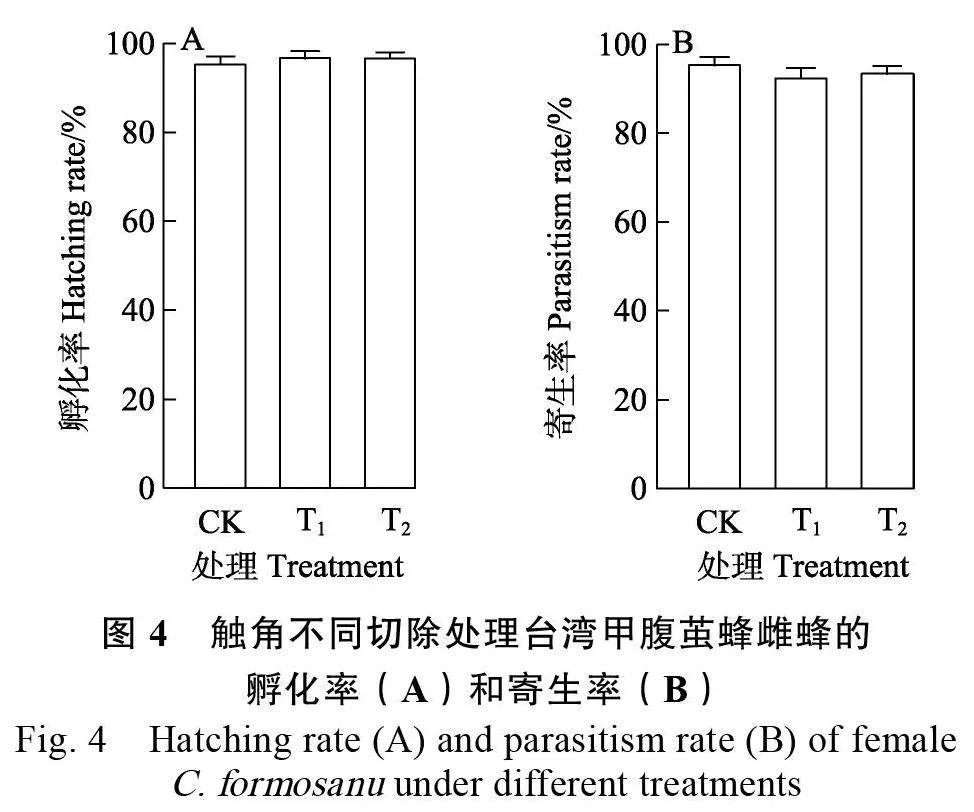
2.1.4 触角不同切除处理对雌蜂寄生卵孵化和寄生的影响 切除台湾甲腹茧蜂不同比例触角对其子代孵化率无显著影响(F=0.88, P=0.43>0.05)。CK 雌蜂寄主子代孵化率为(96.01±3.29)%,与切除1/3 鞭节雌蜂和切除2/3 鞭节雌蜂之间均差异不显著(图4A)。切除台湾甲腹茧蜂不同比例触角对其寄生率也无显著影响(F=1.19, P= 0.32>0.05),其中,CK 雌蜂寄生率最高,为(96.10± 3.15)%,其次为切除2/3 鞭节和1/3 鞭节寄生率,分别为(94.18±3.90)%和(93.11±5.01)%(图4B)。
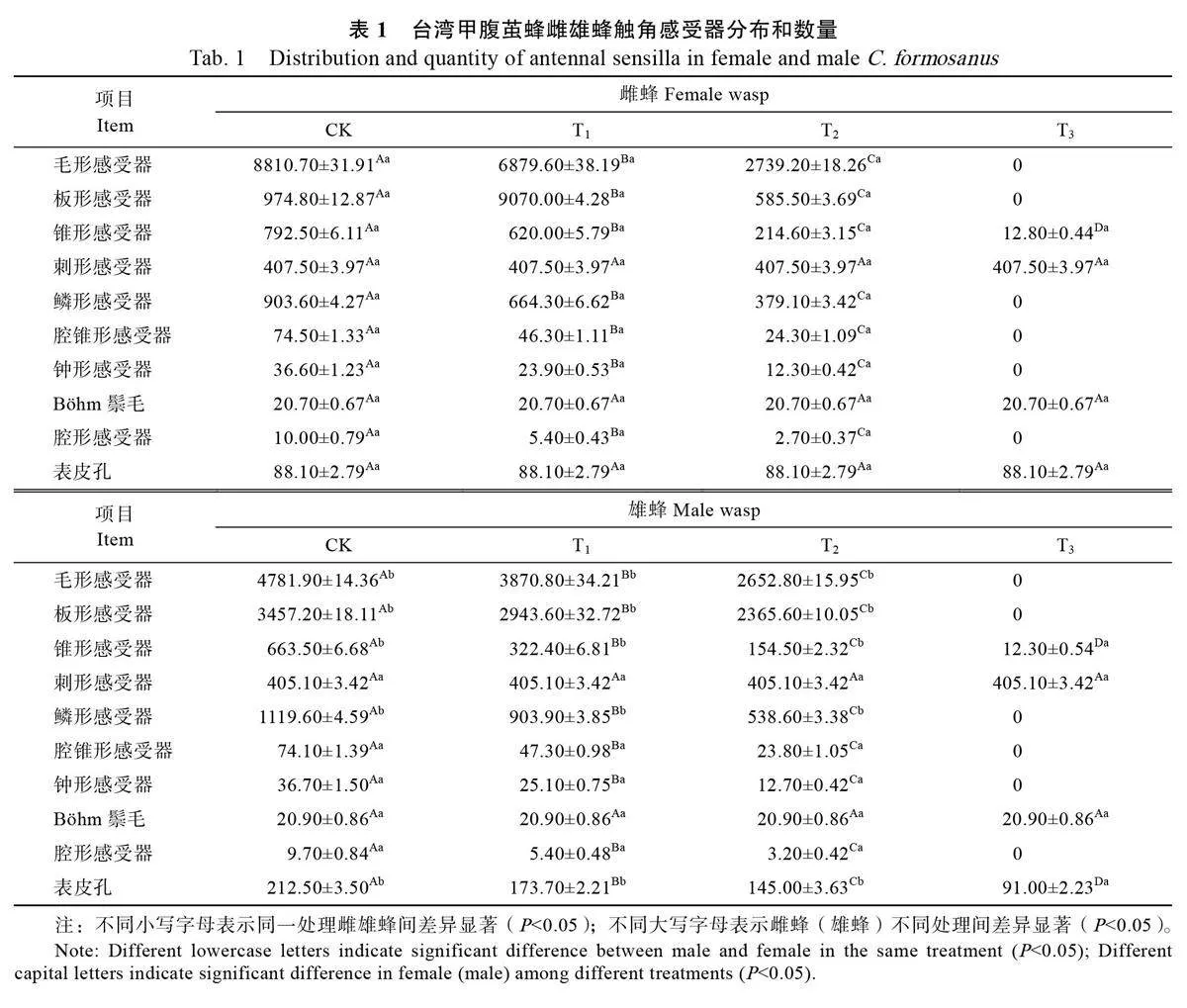
2.2 触角不同切除处理感受器类型、数量和分布
在完整触角雌雄蜂之间,其毛形感受器、板形感受器、锥形感受器、鳞形感受器以及表皮孔数量均存在显著差异(表1)。雌蜂的毛形感受器数量和锥形感受器数量分别为8810.70±31.91、792.50±6.11,均显著多于雄蜂。与之相反,雌蜂的板形感受器数量、鳞形感受器数量和表皮孔数量分别为974.80±12.87、903.60±4.27、88.10±2.79,均显著少于雄蜂。在不同切除处理之间,雌雄蜂的毛形感受器、板形感受器、锥形感受器、鳞形感受器、腔锥形感受器、钟形感受器、雄蜂表皮孔的数量均存在显著差异。毛形感受器、板形感受器和鳞形感受器只分布于触角鞭节,而刺形感受器、Böhm 鬃毛以及雌蜂的表皮孔只分布在柄节上。完全切除触角处理组不存在感受器。
3讨论
本研究发现,与CK 相比,切除1/3 鞭节触角的台湾甲腹茧蜂雌雄蜂找到蜜源的速度更快,这与丽蝇蛹集金小蜂(Nasonia vitripennisp)切除1/3 鞭节触角能够更快找到寄主卵块[16]的研究结果相似。出现此现象的原因可能是切除部分触角后,昆虫对周边的化学信息感知降低,定位蜜源的干扰减少,推测台湾甲腹茧蜂定位蜜源的关键不在触角端部区域内。本研究还发现,与切除鞭节相比,台湾甲腹茧蜂雄蜂完全切除触角后能更快找到蜜源。研究中观察到,将完整切除触角的雄蜂从试管转移到培养皿时,虫体东倒西歪,不能保持平衡,而雌蜂体重较重其活跃度弱于雄蜂,能较快平衡身体。切除完整鞭节和切除完整触角的2 个处理间的差异在于是否切除柄节和梗节,可见柄节和梗节上有保持身体平衡的感受器。台湾甲腹茧蜂与斜纹夜蛾侧沟茧蜂(Microplitis prodeniae)[17]和白蛾黑棒啮小蜂(Tetrastichus septentrionalis)[18]相似,其感受器只分布Böhm 鬃毛和刺形感受器。已有研究表明Böhm 鬃毛和刺形感受器属于机械感受器[19]。因此,切除整个触角后,触角上不存在Böhm鬃毛和刺形感受器,导致身体极度不平衡,以至于在平衡过程中碰巧找到蜜源的几率增加。
切除台湾甲腹茧蜂鞭节后,雌蜂无法找到寄主卵块,此时触角只有刺形感受器、Böhm 鬃毛、锥形感受器和表皮孔感受器,说明这些感受器与雌蜂寄主定位及识别无关。锥形感受器一般分为能感受到外界化学信息的微孔型锥形感受器[20-23]和只能感受机械刺激的锥形感受器,即无孔型锥形感受器[19, 24],所以推测台湾甲腹茧蜂梗节上的锥形感受器是无孔亚型。
从扫描电镜结果中可知,毛形感受器、板形感受器和鳞形感受器是台湾甲腹茧蜂触角上数量最多的感受器,其中只有鳞形感受器是机械感受器,其作用是平衡整个触角,以保持整个身体的平衡[25]。毛形感受器具有机械感受器和化学感受器的功能[13, 26-28]。SACHSE 等[29]发现昆虫毛形感受器能接收外部信息,在寻找异性个体的过程中可以感知高度专一化的性信息素[12, 30]。BARLIN等[21]发现小蜂总科的板型感受器2 种不同类型,其中一种为孔数较多的薄壁型,另一种为孔数较少的厚壁型。另外,PATRICCK 等[30]、AKERS等[31]、PETTERSSON 等[32]学者根据板形感受器表面微孔和感受器内腔多个神经元细胞的特征,表明其具有化学感受功能。研究发现台湾甲腹茧蜂与二化螟盘绒茧蜂(Cotesia chilonis)[33]和棉铃虫齿唇姬蜂(Campoletis chlorideae)[23]相似,均是雌蜂的毛形感受器数量显著多于雄蜂,而板形感受器则是雄蜂多于雌蜂,根据以上研究结果,推测毛形感受器可能主要用于感受雄性信息素,板形感受器可能主要用于感受雌性信息素。
本研究结果表明,切除不同比例触角显著影响台湾甲腹茧蜂雌雄蜂对蜜源和寄主卵的搜寻行为,而对雌雄蜂的寿命以及雌蜂的寄生率和子代孵化率无显著影响。说明切除触角只影响台湾甲腹茧蜂寻找食物和寄主等嗅觉相关活动,并未直接影响到生命体征。本研究从行为学入手,再通过扫描电镜观察,并结合前人的研究结果,对台湾甲腹茧蜂触角感受器的功能进行了推测分析,今后将利用透射电镜以及电生理等技术对其进行更深入的研究。
