
花生PLT基因家族全基因组鉴定与表达分析
2024-09-22 00:00王晓璇王金枝邱鼎殷冬梅
山东农业科学 2024年8期




摘要:PLT家族是植物特有的一类转录因子,在植物胚胎、干细胞、分生组织及器官生长发育等过程都起着重要作用。本研究利用生物信息学技术在栽培种花生基因组中鉴定到12个PLT家族基因,分布在11条染色体上:AhPLT蛋白大多含有2个保守的AP2结构域,编码439 - 713个氨基酸,预测定位在细胞核或叶绿体中;AhPLT基因结构复杂,具有多样的短外显子结构,外显子数在5-8个之间,不同家族成员中分布不同的保守基序:AhPLT家族成员启动子区存在生长素、赤霉素和茉莉酸等激素相关的顺式作用元件。qRT-PCR结果显示,AhPLT1 -B和AhPLT5-B分别在根中和种子中的表达量最高:两基因对激素IAA、6-BA和GA3有不同的响应,其中AhPLT1 -B受6-BA调控上调表达最显著,AhPLT5-B受6-BA、IAA和GA,调控呈先升高后降低的表达模式。本研究为花生PLT基因的生物学功能研究提供了参考。
关键词:花生:PLT转录因子:基因家族:生物信息学分析:表达分析
中图分类号:S565.2:Q781 文献标识号:A 文章编号:1001-4942(2024) 08-0001-09
PLETHORA(PLT)属于APETALA 2/ETHYL-ENE RESPONSE FACTOR(AP2/ERF)型转录因子,PLT家族成员在植物的新生分生组织中表达水平较高,参与新生分生组织的形成和维持以及器官的启动和发育过程。PLT基因的结构包括两个保守的AP2重复结构域,两者之间存在较大的结构差异。
PLT在根发育过程中扮演重要角色,在促进根尖分生组织(root meristem,RM)的形态建成和活性维持,特别是静止中心(quiescent center,QC)的定位中至关重要。QC在根尖分生组织中充当干细胞维持所需的组织中心。PLT1和PLT2被认为是维持根尖干细胞和QC细胞的胚胎规格所必需的两个功能冗余基因,QC的形成要求PLTI-PLT2和SHR-SCR共同参与。PLT对于促进植物芽再生过程也具有重要作用。研究表明,PLT3、PLT5和PLT7通过复杂的机制调控拟南芥(Arabidopsis thaliana L.)芽再生的过程。具体而言,在芽从头再生途径中,侧器官边界相关基因CUC2发挥着关键作用,而PLT则通过调节CUC2的表达水平来促进芽的再生。另外,PLT还需要与CUC基因相互作用,以参与芽的再生调控过程。豆科植物的根瘤中寄居着固氮根瘤菌,这些根瘤形成在根部表示可能存在着与根相关的因素。研究发现,苜蓿(Medicago truncatula L.)中4个MtPLT基因在根结节发育和维持中起着冗余的功能作用,与拟南芥根部发育中的PLT基因相似。在干细胞的研究中,LT1和PLT2的启动子活性以及蛋白结合物呈梯度状分布,并在干细胞区域达到最大值。PLT活性很大程度上是累加性和剂量依赖性的,干细胞状态受PLT活性水平的调控,高水平的活性有助于保持干细胞的原始状态,较低水平的活性则促进干细胞有丝分裂,更低水平的PLT活性则会促进干细胞向分化状态转变。在根干细胞区域和不定芽生成过程中,可以观察到PLT基因的特异表达。利用农杆菌侵染的方法,在金鱼草(Antirrhinum majus L.)中侵染带有PLT5基因的农杆菌可诱导不定芽的形成。用同样的方法,在番茄(Solanum lyco-persicum L.)和两个甘蓝品种(Brassica rapa,var' Bok choy' and' Pei Tsai')中侵染,都能获得稳定遗传的不定芽,表明应用PLT5可提高植物的转化效率,并且这种转化系统可促进基因组编辑或其他植物生物技术在现代农业中的应用。
花生(Arachis hypogaea L.)作为一种重要的油料和经济作物,在我国具有广泛的适应性和显著的经济效益等优点,总产量在油料作物中名列前茅。PLT基因家族在植物根、叶、花、根瘤等器官的生长发育,以及辅助遗传转化中发挥至关重要的作用。国内对PLT基因家族的研究相对较少,而且在花生领域几乎没有相关报道,因此对花生PLT基因家族进行研究具有重要的意义。花生遗传转化目前仍较困难,研究PLT基因家族将有助于开发新的花生转化系统。本研究通过信息生物学手段对栽培种花生PLT基因家族成员进行全基因组鉴定和表达分析,为花生PLT基因家族功能研究以及花生再生体系建立奠定基础。 1 材料与方法
1.1 花生PLT基因家族成员鉴定和染色体定位分析
借助Pfam数据库(http://pfam. xfam. org/)提供的PLETHORA(PF00847)结构域的隐马尔可夫模型文件,利用TBtools软件对花生数据库进行搜索,接着运用CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)进行保守结构域鉴定和筛选。通过ExPASy(https://web.expasy.org/protparam/)在线网站分析花生PLT成员的氨基酸数量、蛋白质分子量、等电点、亲水性以及亚细胞定位特征,利用SOPMA(https://npsa-prabi.ib-cp.fr/cgi-bin/npsa_automat. pl? page= npsa_sopma.html)在线网站对AhPLTs蛋白的二级结构进行详细分析与结构预测。将PLT基因家族成员的基因名称及其在染色体上的位置上传至在线网站MG2C(http://mg2c.iask.in/mg2c_v2. 11),对花生PLT基因在染色体上的定位进行分析。
1.2 花生PLT基因家族系统发育、基因结构及蛋白保守基序分析
为了研究PLT基因家族的进化关系,从Phy-tozome数据库(https://phytozome-next. jgi. doe.gov/)中下载花生、拟南芥、水稻(Oryza sativa L-)和苜蓿等物种的PLT基因序列和编码蛋白氨基酸序列,利用TBtools软件绘制系统进化树,将输出的nwk. file文件上传至在线网站(https://itol.embl.de/)对进化树进行美化。通过MEME在线网站(https://meme-suite.org/meme/doc/meme.html)对PLT蛋白序列进行保守基序motif分析,将保守基序数目设置为10;之后将花生PLT基因家族的gff3文件、保守基序分析结果(meme.XML)文件和进化树分析结果(nwk.file)文件上传至TBtools软件对基因结构进行可视化分析。使用NCBI数据库BLAST进行同源蛋白序列比对,选取跨物种同源蛋白序列,并将其导人ClustalX进行多序列比对。利用在线工具(https://www.ebi.ac. uk/Tools/hmmer/)分析花生PLT蛋白的保守结构域,将数据上传至TBtools软件进行可视化分析。
1.3 花生PLT基因家族顺式作用元件预测
通过分析AhPLTs起始密码子上游2 000 bp序列,并参考Chao和Wang等的研究方法,预测顺式作用元件。
1.4 花生PLT基因家族在不同组织部位的表达分析
为进一步了解PLT基因在花生不同组织中的表达情况,在Peanut Base(https://peanutbase.org/)数据库中下载参考基因组(arahyTifrunner.GnmI.Ann1. CCJH)的表达谱数据,从中筛选出PLT基因家族成员在19个花生组织中的表达数据,包括叶片、茎尖、根、根瘤、花被、雄蕊、雌蕊、荚果、果针、果壳和种子等。使用联川生物云平台站(https://www.omicstudio.cn/tool/4)对花生PLT基因家族各个组织表达量归一化(数值进行Log2处理),随后构建基因表达热图。
1.5 花生PLT基因家族不同激素处理下表达模式分析
供试花生品种为农大花108,对成熟种子及萌发生长3周左右花生植株的根、茎、茎尖和叶片取样:对萌发生长3周左右的幼苗分别喷施50μmol/L IAA、25 μmol/L 6-BA和100 μmol/L GA3进行激素处理,并在0(处理前)、1、3、6、12、24 h取植株叶片。每个试验3次生物学重复,每个重复取3株的样品混合。将所取材料装入无RNA酶离心管中,液氮速冻后-80℃保存。
通过qRT-PCR检测PLT基因在不同激素处理下的表达情况,引物信息见表1。根据TransS-cript@Green One - Step qRT - PCR SuperMix(TRANS)说明书进行qRT- PCR。反应体系(20μL):Mix 10 μL,ddH20 8.2 μL,cDNA模版1 μL,上、下游引物各0.4 μL。反应程序:95℃5 min,95℃10 s,60℃30 s,40个循环。内参基因使用actin,利用2-△△Ct方法计算AhPLT基因的相对表达水平,然后运用Graphpad Prism软件进行数据可视化呈现,分析花生PLT基因在组织器官和不同激素处理下的表达情况。
2 结果与分析
2.1 花生PLT基因家族成员鉴定和染色体定位
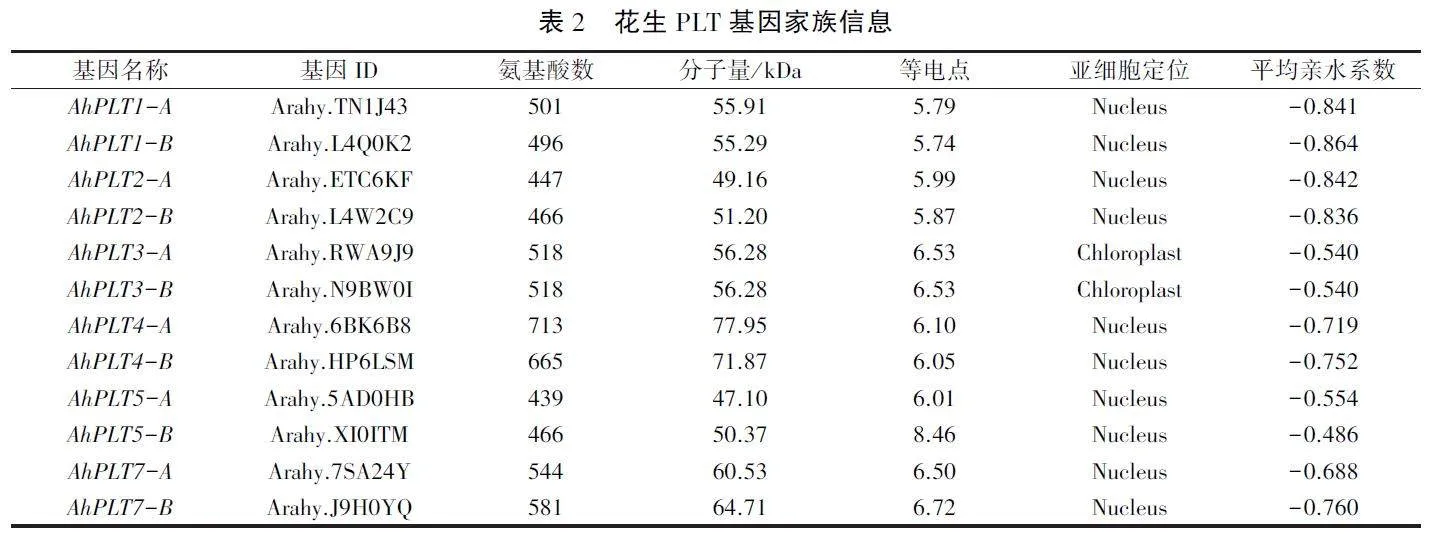
本研究从花生全基因组中共鉴定到12个PLT基因家族成员,分别命名为AhPLTl -A、Ah-PLT1-B、AhPLT2 -A、AhPLT2 -B、AhPLT3 -A、Ah-PLT3-B、AhPLT4 -A、AhPLT4 -B、AhPLT5 -A、Ah-PLT5-B、AhPLT7 -A、AhPLT7 -B。编码439 - 713个氨基酸,分子量在47.10 - 77.95kDa之间,等电点为5.74-8.46(表2)。AhPLT蛋白的亚细胞定位预测结果(表2)显示,除AhPLT3-A和AhPLT3-B两个蛋白定位于叶绿体外,其他蛋白均定位在细胞核中。花生PLT蛋白平均亲水系数均为负值,说明这些蛋白均为亲水性蛋白。
染色体定位结果(图1)显示,AhPLT成员的染色体分布并不均匀,12个AhPLT基因分布在花生的11条染色体上,但大部分染色体上只分布有1个基因,只有8号染色体上分布有2个基因。
2.2 花生PLT基因家族系统进化树构建、基因结构及蛋白保守基序分析
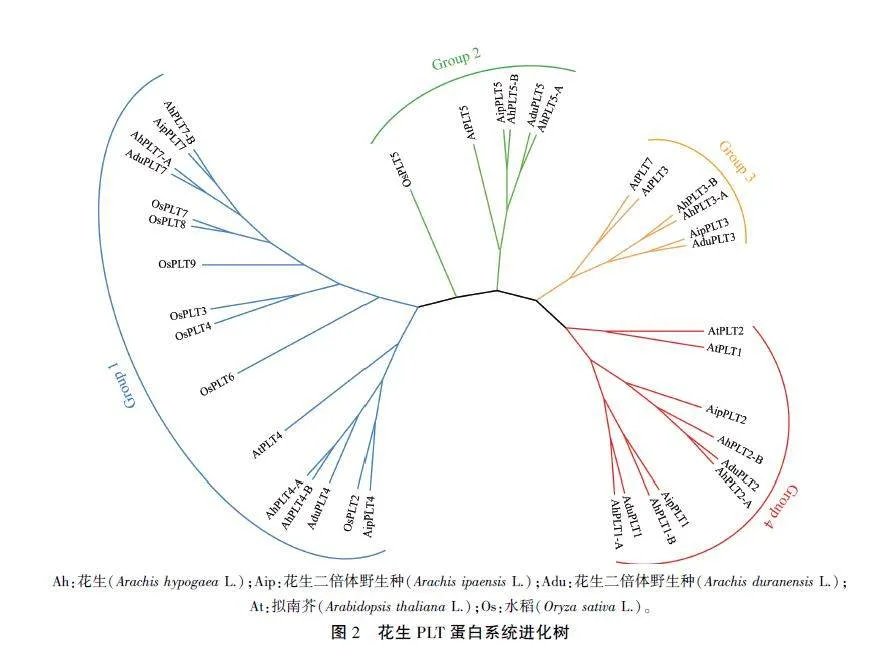
将12个花生PLT蛋白与拟南芥、水稻、二倍体花生中的PLT蛋白进行系统发育分析。结果(图2)表明,12个花生PLT蛋白在进化关系上分为4个分支。其中,AhPLTI-A、AhPLTl-B与At-PLTI/AIL3(AT3G20840.1)和AtPLT2/AIIA(ATIG51190.1)聚类到同一分支,AhPLT3 -A、Ah-PLT3-B与AtPLT3/AIL6(AT5G10510.1)和At-PLT7(AT5G65510.1)聚类到同一分支,AhPLT4 -A、AhPLT4 -B与AtPLT4(AT5G17430.1)聚类到同一分支,AhPLT5 -A、AhPLT5 -B与AtPLT5(AT5 G57390.1)聚类到同一分支,表明它们之间存在密切的亲缘关系。
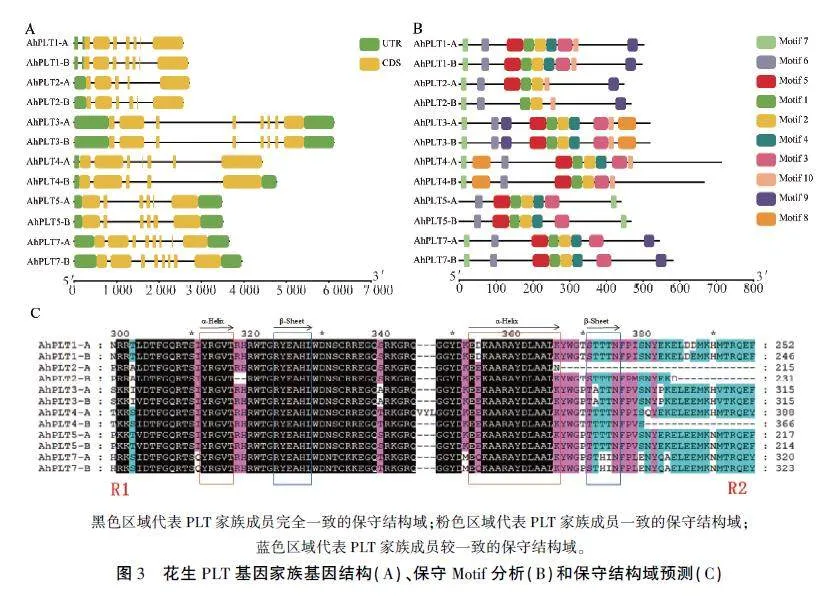
基因结构分析结果(图3A)表明,PLT的结构复杂且具有多样的短外显子结构,外显子数介于5-8个之间。其中,AhPLT3 -A、AhPLT3-B、AhPLT7-A和AhPLT7-B外显子数目最多,各有8个,AhPLT2 -A和AhPLT4-B的外显子数目最少,各有5个。
通过MEME在AhPLT家族中检索到10个保守motif(图3B),其中Motif 1、Motif 2、Motif 6和Motif 7在12个成员中均存在:只有AhPLT2-B缺失Motif 5;AhPLT2-A和AhPLT2-B缺失Motif 3和Motif 4;除AhPLT5 -A、AhPLT5 -B、AhPLT7-A和AhPLT7-B外,其他成员均有Motif 10。值得注意的是Motif 7位于AhPLT5-A和AhPLT5 -B的C端,而在家族其他成员中均位于N端,这是两者区别于家族中其他成员的结构。结果表明,在AhPLT进化过程中家族成员的蛋白序列相对保守。
花生PLT家族成员的蛋白保守结构域分析结果(图3C)显示,AhPLT蛋白含有AP2和AP2superfamily等结构域,其中所有成员均含有AP2结构域,只有AhPLT3-A、AhPLT3-B、AhPLT5 -B、AhPLT7 -A含有AP2 superfamily。多序列比对结果显示,该家族蛋白的结构特点主要表现为包含两个具有高度保守性的AP2结构域,分别命名为R1和R2,其中,AhPLT2 -A、AhPLT2 -B和Ah-PLT4-B蛋白的R2结构域缺失。
2.3 花生PLT基因家族顺式作用元件
通过对AhPLT基因家族成员起始密码子上游2 000 bp序列进行分析,发现其中不仅含有光响应和逆境响应等元件,还拥有植物激素应答相关的元件。PLT基因在调节植物器官的生长发育过程中,通过响应各种植物激素发挥作用。选取生长素(TGA - element,AACGAC)、水杨酸(TCA-element,CCATCTTTTT)、茉莉酸(TGACG - motif,TGACG)、脱落酸(ABRE,ACGTG)、种子特异性(RY-element,CATGCATG)、光响应(MRE,AAC-CTAA)和赤霉素(P-box,CCTTTTG)7类顺式作用元件进行分析,结果(图4)发现,AhPLT基因家族成员启动子区域包含的ABA响应元件最多,仅在AhPLT4-B和AhPLT7-A上未预测到:赤霉素元件在大多数成员中也都可以预测到,而生长素相关元件仅存在于AhPLT1 -A、AhPLT3 -A和Ah-PLT3 -B上。种子特异性元件只有AhPLT1和Ah-PLT5启动子包含,表明这两个基因可能参与花生种子发育的调控过程。
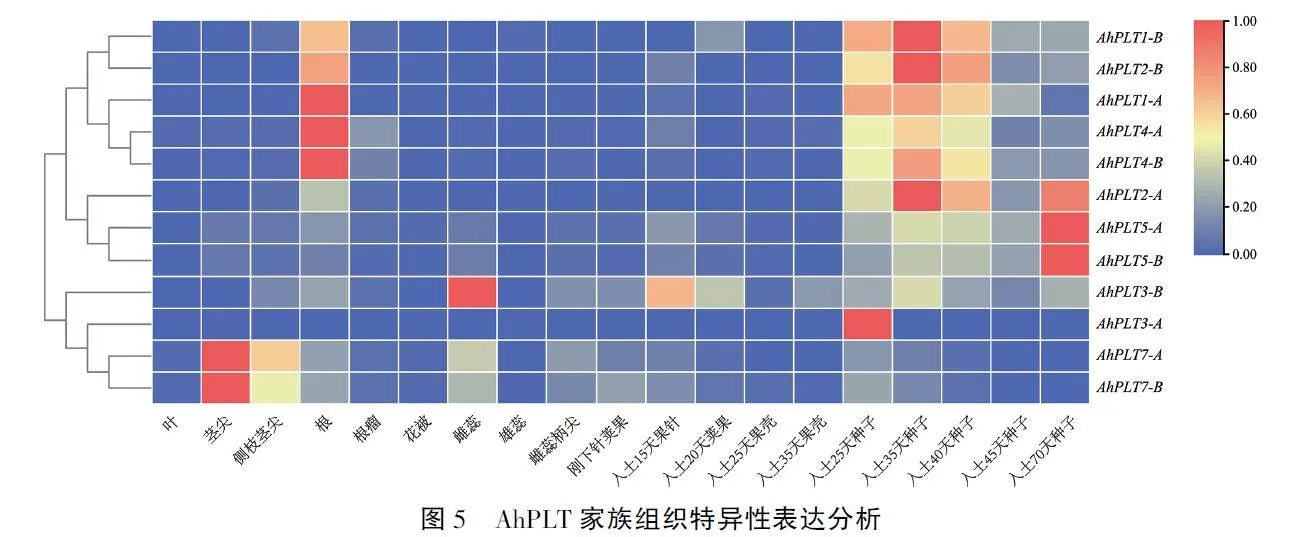
2.4 花生PLT基因家族组织特异性表达分析
为了探究PLT家族基因在花生不同组织部位的表达差异,在花生数据库Peanutbase中下载花生参考基因组表达谱数据,从中筛选出PLT基因家族成员在19个花生组织中的表达谱数据。绘制热图后结果(图5)显示,12个AhPLT基因的表达模式表现出明显的组织特异性。AhPLT1、AhPLT2和AhPLT4在根和种子发育的早期有较高的表达水平,推测它们可能与花生根和种子的发育相关。AhPLT5同样可能参与花生种子的发育过程,但其可能主要在种子发育的后期发挥作用。AhPLT3 -B在雄蕊中的表达量最高,推测其可能参与花生雄蕊或花粉发育的调控过程。Ah-PLT7在主茎茎尖和侧枝茎尖的表达量较高,表明其可能参与花生顶端分生组织活性的调控过程。
2.5 花生PLT1和PLT5基因在不同激素处理下表达模式分析
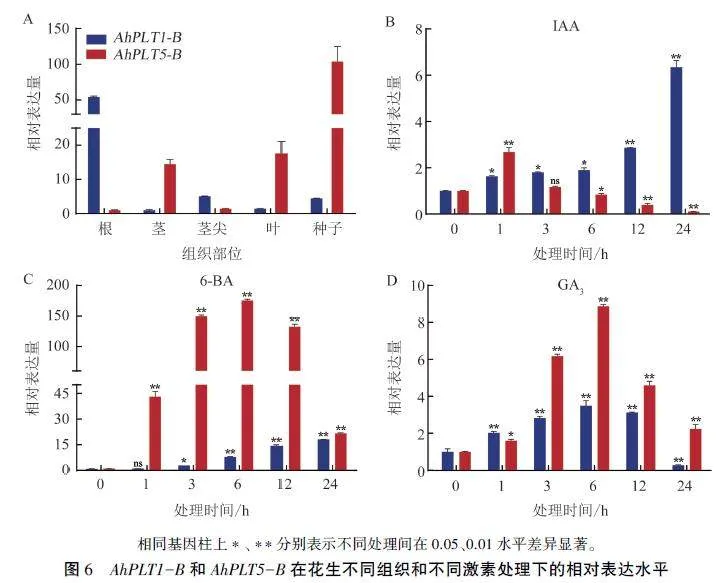
基于PLT1和PLT5基因的重要功能,本研究利用qRT-PCR检测了AhPLTl -B和AhPLT5-B在花生不同组织部位和和3种激素处理下的表达模式。结果表明,AhPLTl -B在茎、茎尖、叶和种子中表达量较低,在根中表达量最高;AhPLT5 -B在种子中的表达量最高(图6A)。在IAA处理下,花生叶片中AhPLT1 -B的表达量随处理时间的延长呈现明显上调趋势,AhPLT5 -B表达量在处理1 h达到最大值,之后逐渐下降(图6B);经过6-BA处理后,AhPLTl -B和AhPLT5-B都有不同程度的上调,从3h开始AhPLT1 -B的表达量升高,24 h达到最高,AhPLT5-B表达量6h达到最大值,之后有所下降(图6C);在GA,处理下,Ah-PLTl -B和AhPLT5-B的表达模式类似,表达量都在6h达到最大值,随处理时间延长,表达量开始出现下降趋势(图6D)。AhPL T1 -B和AhPLT5 -B对IAA、6-BA和GA,都有不同程度的响应,其中对6-BA的响应最明显,表明6-BA在两基因发挥功能时起着重要作用。
3 讨论与结论
AP2/ERF作为一类独特的转录因子,在植物中发挥着重要作用。PLT基因家族作为其中的重要成员,在调控植物胚胎发育、维护分生组织以及促进器官生长等过程中扮演关键角色。在本研究中,通过对栽培种花生全基因组进行深入鉴定,成功筛选出12个PLT基因,同时,结合几种模式植物如拟南芥、水稻等的进化分析,对其进行了命名,揭示PLT基因在植物界的分子进化过程。拟南芥中PLT5参与调控不定芽的生长发育,在番茄中过表达PLT5也可以促进创伤处长出新的愈伤和不定芽。进化分析结果表明,花生AhPLT5-A、AhPLT5 -B与AtPLT5具有较近的亲缘关系,因此推测AhPLT5 -A和AhPLT5-B为AtPLT5的直系同源基因,且也具有促进愈伤组织和不定芽再生的功能。
PLT基因家族含有2个保守的AP2结构域,基因结构比较复杂,外显子数量较多。对花生PLT基因结构进行分析发现,其外显子数量为5-8个,与拟南芥、水稻等模式植物基本一致,说明不同物种中PLT基因家族在进化过程中相对保守。保守结构域分析和AP2结构域多序列比对分析也表现出相似的结果。
PLT基因家族成员参与植物花、芽、结节以及根等组织和器官的发育过程。本研究使用19个花生组织的表达数据对12个AhPLT基因表达模式进行分析,发现大部分基因表达量比较低,个别基因在根、茎尖和种子中的表达量相对较高,推测PLT家族成员在这些花生器官的生长发育过程中发挥重要作用。PLT1和PLT5在愈伤组织和不定芽的生长过程中起着重要的作用,利用PLT5进行遗传转化还能够提高转化效率。AhPLT1和AhPLT5的启动子上包含响应生长素和细胞分裂素等激素的调控元件,说明它们可能通过响应这些激素信号诱导参与调控植物的生长发育过程。已有的研究表明,PLT5在芽原基起始和活性维持中起重要作用,而PLT1作为核心模块调控根尖的发育。组织特异性表达分析的结果表明,这两个基因可能还参与了花生种子的发育调控过程进而影响花生的产量。对花生进行不同激素处理后,两个PLT基因对激素IAA、6-BA和GA。都有不同程度的响应,其中AhPLT1 -B受6-BA调控上调表达最显著,AhPLT5 -B受6-BA、IAA和GA,调控呈先升高后降低的表达模式。同时,研究发现应用PLT1和PLT5可以提高愈伤组织和不定芽的再生效率,具有提高花生遗传转化效率的应用潜力。
综上所述,本研究在栽培种花生中鉴定到了12个AhPLT基因,并对PLT家族成员进行了详细的特征分析与进化分类,结合AhPLT基因表达数据,对基因在不同激素处理下表达模式进行了分析,筛选出AhPLT1和AhPLT5两个与花生再生相关的候选基因,并发现细胞分裂素6-BA可能参与了对它们功能的调控。以上结果表明,AhPLT基因可能在花生的生长发育和再生过程中具有重要作用,但其调控机制仍需进一步探讨。
基金项目:河南省重大科技专项(221100110300);河南省重点研发专项(221111110500);河南省产业技术体系专项(HARS-22-05- G1)
