
柏木属的分类地位和物种多样性研究现状与建议
2024-06-16 00:00:00李家亮巫大宇毛康珊
南京林业大学学报(自然科学版) 2024年3期
关键词:生物多样性



摘要:柏木属(Cupressus L.)是柏科的模式属,1753年由卡尔·林奈在其著作《植物种志》中命名,其模式种为地中海柏木(Cupressus sempervirens L.)。但是,随着越来越多的物种得以发现和描述,以及近几十年来系统发育学和群体遗传学研究的开展,柏木属的分类地位和物种数量争议不断。2011年,Christenhusz等提出现存裸子植物新的分类系统时(即克氏裸子植物系统),柏木属 (Cupressus sensu Christenhusz) 的概念可涵盖4个属级类群:狭义柏木属、金柏属、北美金柏属、美洲柏木属;通常广义柏木属 (Cupressus sensu lato) 涵盖狭义柏木属和美洲柏木属物种;而狭义柏木属 (Cupressus sensu stricto) 仅包含广义柏木属中分布在旧世界的物种。基于21世纪以来国内外学者对柏木属及其近缘属的研究进展,以及笔者自身的研究和野外考察经验,笔者简要综述了该属的分类地位和物种多样性。最新的系统发育基因组学证据表明,狭义柏木属为单系群且与刺柏属互为姊妹关系,二者的共同祖先与金柏属、北美金柏属、美洲柏木属的共同祖先互为姊妹关系。对一些具争议的分类处理提出了新的见解,将Cupressus rushforthii Maerki amp; J.Hoch降级为Cupressus austrotibetica Silba的异名,并确认剑阁柏木为大渡河柏木(Cupressus fallax Franco)和柏木(Cupressus funebris Endlicher)的杂交后代,将其拉丁名修订为Cupressus × jiangeensis N.Zhao;同时,将王氏柏木(Cupressus ×wangii J.Hoch, Maerki amp; Rushforth)处理为剑阁柏木的异名;另外,为避免混淆,建议将西藏波密县新近发现的亚洲最高树种Cupressus austrotibetica Silba中文名修改为“西藏高柏”,将主要分布在尼泊尔的“西藏柏木”(Cupressus torulosa D.Don ex Lamb.)中文名恢复为“喜马拉雅柏”。根据目前的研究现状,狭义柏木属应包含14种,其中包括1个杂种。为确保未来柏木属物种划分的准确性和科学性,建议采用整合物种概念,在分类工作中综合考虑形态学、遗传学、生态学和生物地理学等多维度数据。新一代测序技术等前沿技术方法为柏木属系统分类与进化研究提供了强大工具,未来对狭义柏木属物种多样性及其演化历史的探索和理解还将进一步深入。
关键词:柏木属;分类修订;物种概念;生物多样性
中图分类号:S685.99;S718 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)03-0036-10
Taxonomy and diversity of the genus Cupressus: current status and recommendations
LI Jialiang, WU Dayu, MAO Kangshan*
(College of Life Sciences, Key Laboratory of Bio-Resource and Eco-Environment of Ministry of Education, Sichuan University, Chengdu 610065, China)
Abstract:Cupressus L., the type of Cupressaceae family, is typified by Cupressus sempervirens L. Over the years, as many other species have been discovered and described, and with advances in phylogenetics and population genetics, the taxonomy and species diversity within the genus Cupressus have been subjects of continuous debate. Traditionally, the Cupressus sensu lato includes species from true Cypresses sensu stricto and New World cypresses (Hesperocyparis Bartel amp; R.A.Price), while the Cupressus s. s. comprises all Old World species except the Vietnamese golden cypress (Xanthocyparis vietnamensis Farjon amp; T.H.Nguyên) found in Vietnam. Christenhusz et al. defined the broadest interpretation of Cupressus (Cupressus sensu Christenhusz) which encompasses four groups: true cypresses (Cupressus s.s.), Vietnamese golden cypress, Nootka cypress (Callitropsis nootkatensis (D.Don) Oerst), and New World cypresses. These ideas have been challenged by recent phylogenomic studies. Recent studies have shown that Cupressus s.l. is non-monophyletic, Cupressus s.s. is monophyletic and forms a sister group with the genus Juniperus. The common ancestor of these two is a sister group of the common ancestors of the Xanthocyparis, Callitropsis and Hesperocyparis. Based on current research, the true cypresses should encompass 14 species, including one hybrid. This paper offers new perspectives on previous contentious taxonomic treatments, for instance, reclassifying Cupressus rushforthii Maerki amp; J.Hoch as a synonym of Cupressus austrotibetica Silba, and confirming that Cupressus × jiangeensis N. Zhao is a hybrid offspring of the Cupressus fallax Franco and Cupressus funebris Endlicher. Additionally, Cupressus ×wangii J.Hoch, Maerki amp; Rushforth has been reduced to a synonym of Cupressus × jiangeensis. To avoid confusion, it is recommended to rename the recently discovered tallest tree species in Bomi County, Cupressus austrotibetica, to “西藏高柏” (Xizang tall cypress) in Chinese, and the cypress predominantly found in Nepal, Cupressus torulosa D.Don ex Lamb., is referred to as “喜马拉雅柏” (Himalayan cypress) in Chinese, instead of “西藏柏木” (Xizang Cypress). To ensure accuracy and scientific validity in future studies on species delineation within the genus, we recommend integrating multidimensional data from morphology, genetics, ecology and biogeography. This approach will help the reducing of classification confusion and support biodiversity conservation efforts more effectively.
Keywords:Cypressus L.; taxonomic revision; species concept; biodiversity
柏木属(Cupressus L.)隶属柏科(Cupressaceae),为该科的模式属。根据最新的裸子植物分类系统[1],狭义柏木属植物广泛分布于欧亚大陆和非洲北部;柏木属植物在我国分布在秦岭及长江流域以南、横断山区和喜马拉雅山区;在国外则分布在亚洲西部、撒哈拉地区和欧洲南部等地中海沿岸,以及东南亚地区[2]。分布在横断山和喜马拉雅山区的物种,如岷江柏木(C. chengiana S.Y. Hu)、干香柏(C. duclouxiana Hickel)、巨柏(C. gigantea W.C.Cheng amp; L.K.Fu)等,多为干旱/干热河谷等脆弱生境的建群种,对当地生态系统具有不可或缺的作用。在我国南方,柏木(C. funebris Endl.)是一种极为重要的栽培树种,尤其是四川省翠云廊古蜀道所栽种的古柏,自秦汉以来已有逾两千年的栽培历史,承载了深厚的文化价值。2023年,在西藏雅鲁藏布江流域,发现了一株高达102.3 m的柏木属植物(按照最新分类处理,其拉丁名应为Cupressus austrotibetica Silba),这是迄今所知亚洲最高、世界第2高树种[3]。柏木属的模式物种地中海柏木(C. sempervirens L.)栽培广泛且历史悠久,在地中海盆地、西亚、亚细亚、北非和北美地区的文化中具有重要地位[4],法国近代著名画家文森特·梵高在《星空》《麦田中的柏树》等画作中均描绘了地中海柏木蓬勃的生命力。因此柏木属在生态、经济和文化层面上均具有重要价值。
柏木属的分类地位及其物种多样性长期以来一直是针叶树系统分类与进化研究领域争论的焦点之一[5]。尤其是广义柏木属中新世界分布的物种是否应该独立成属,柏木属包含多少种,如何对其进行有效保护等问题,在国际上尚未达成一致意见[6]。广义柏木属中新世界分布的物种与旧世界分布的物种在形态上存在相似之处,但它们之间的分子系统发育学关系却显示出明显的分歧[7]。此外,在种的层面上,不同种之间由于长期的地理隔离和杂交渐渗作用,可能产生了隐存种或杂交种,这导致不同的学者对该属的分类处理持有不同观点。特别是,受人类活动和气候变化的影响,许多柏木属物种正面临灭绝的威胁。例如,巨柏和西藏柏木(C. torulosa D. Don ex Lamb.;按照最新分类学文献,中国境内分布的“西藏柏木”种群应为C. austrotibetica)被列为国家一级保护植物,岷江柏木则是国家二级保护植物;摩洛哥柏木(C. atlantica Gaussen)、撒哈拉柏木(C. dupreziana A. Camus)和剑阁柏木(C. jiangeensis N. Zhao)更是被国际自然保护联盟(IUCN)列为“极危(CR)”等级。因此,准确界定柏木属的分类地位、梳理其物种多样性,对于促进柏木属植物的保护和资源的可持续利用具有至关重要的意义。笔者基于对有关柏木属文献资料的广泛收集和整理,结合四川大学生命科学学院柏木分类课题组对该属物种开展的系列研究,对柏木属的分类地位和物种多样性进行简要综述。
1 柏木属的分类地位和分子系统学研究进展
柏木属最初由卡尔·林奈在其于1753年发表的著作《植物种志》(Species Plantarum)中描述和命名,其模式物种为主要分布在地中海盆地及周边地区的Cupressus sempervirens(地中海柏木)。这个属的典型特征包括:球果第2年成熟,球形或近球形,球果上的木质种鳞超过6片,球果中具有大量小型带翅的种子。基于这些特征,植物分类学者不断描述和发表新种,柏木属逐渐扩展成既有新世界(美洲)物种,也有旧世界(欧洲、非洲和亚洲)物种的大陆间断分布属,即广义柏木属(Cupressus s.l.)。《中国植物志》认为,柏木属共有约20种,其中我国产5种、引种栽培4种[2]。英国邱园(The Royal Botanic Gardens, Kew)的植物分类学家Aljos Farjon在其专著《柏科和金松属专著》(A monograph of Cupressaceae and Sciadopitys)中,提出柏木属包含15种,其中7个来自旧世界,8个来自新世界[8]。在《中国植物志》英文修订版(Flora of China)中,列举了在中国分布和引种栽培的柏木属种9个,其中包括引种自新世界的绿干柏(C. arizonica Greene)、加利福利亚柏木(C. goveniana Gordon)和墨西哥柏木(C. lusitanica Miller)[9]。
近年来,随着分子系统学的不断发展,学者开始认识到广义柏木属实际上并非是一个单系。2006年,美国学者Damon P Little[10]对当时已知的28个广义柏木属现存物种和北美金柏属进行了研究,他使用两套核基因序列(NEEDLY intron 2, nrITS)和3套叶绿体DNA序列(matK、rbcL和 trnL)进行分析,结果表明,旧世界分布的柏木属种与刺柏属(Juniperus L.)种聚在同一分支上;而新世界分布的柏木属种则与北美金柏属(Callitropsis Oerst.)种聚在一起,它们进一步与金柏属(Xanthocyparis Farjon amp; T. H. Nguyên)一起构成一个独立的分支。因此,Little[10]提出柏木属应仅限于旧世界的物种,即狭义柏木属(Cupressus s.s.),而将新世界的类群归到北美金柏属(Callitropsis sensu Little)下(表1)。
2009年,美国学者Robert Adams等[7]运用不同于Little[10]的分子标记,包括3套核基因序列[nrDNA(ITS)、4-coumarate: CoA ligase、abscisic acid-insensitive 3]和一套叶绿体DNA序列(petN-psbM),对旧世界柏木属种、新世界柏木属种、越南黄金柏、北美金柏和刺柏属种之间的系统发育关系进行了深入分析[7]。他们的研究结果与Little[10]的结论相一致,均支持新世界柏木属物种构成了一个独立的谱系。Adams等[7]还指出,新世界柏木属物种在形态上与北美金柏存在显著差异:新世界柏木属的球果含有更多种子(每个球果60~150粒种子),而北美金柏球果中的种子数量通常不超过15粒;此外,新世界柏木属的球果鳞片更加木质化且更大,子叶数量一般为3~5片,而北美金柏子叶仅有2片。鉴于新世界柏木属种独特的形态学特征和系统发育关系,Adams等[7]提出了一个新属——美洲柏木属(Hesperocyparis Bartel amp; R.A.Price),以区分这一独特的进化分支。据此,美洲柏木属、北美金柏属、金柏属形成HCX分支(Hesperocyparis-Callitropsis-Xanthcyparis),刺柏属和狭义柏木属则形成另一个分支。
随后,Mao等[11-12]在全面收集刺柏属、柏木属、美洲柏木属种以及北美金柏和越南黄金柏在内的分子样本的基础上,使用超过10 000个碱基对的叶绿体DNA片段,对这些类群及其近缘类群的系统发育关系进行了深入探究。结果表明,新世界分布的美洲柏木属(Hesperocyparis)、北美金柏属(Callitropsis)和越南分布的金柏属(Xanthocyparis)3个属的种聚在一起,形成了前文提及的HCX分支。此分支与旧世界分布的柏木属[即狭义柏木属(Cupressus s.s.)]种聚为一支,而所有刺柏属种则形成了一个单独的分支。这种与以往研究完全不同的系统发育关系表明这几个类群之间的亲缘关系需要更多的证据来支撑。鉴于柏木属的系统发育位置还未得到完全的解决,Christenhusz等[13]在2011年提出现存裸子植物新的分类系统(即克氏裸子植物系统)时,保守地将狭义柏木属(Cupressus s.s.)、美洲柏木属(Hesperocyparis)、北美金柏属(Callitropsis)、金柏属(Xanthocyparis)4个属归入(克氏)柏木属 (Cupressus sensu Christenhusz)中。
随着高通量测序技术的兴起,研究者们得以在基因组层面上获得大量数据,进而深入探讨柏木属的系统发育地位。2018年,Zhu等[14]利用该技术收集了叶绿体全基因数据,以探究狭义柏木属(Cupressus s.s.)、美洲柏木属(Hesperocyparis)、北美金柏属(Callitropsis)以及金柏属(Xanthocyparis)之间的亲缘关系。研究发现,叶绿体基因组中不同片段在构建系统发育树时显示出显著的异质性。特别是,横跨ycf1和ycf2两个基因的约1.5万碱基长度的序列,强烈支持狭义柏木属与刺柏属形成一个单系分支;而叶绿体基因组的其他基因则强烈支持狭义柏木属与HCX分支是姊妹群,这与Mao等[11-12]在2010年和2012年的研究结果一致。随后,Mao等[5]于2019年进一步使用转录组测序获取了73个单拷贝核基因序列,并应用系统发育基因组学方法分析了这些分支之间的进化关系,进一步探究了导致系统发育冲突的原因。研究表明,不论采用何种建树方法,结果都一致支持狭义柏木属(Cupressus s.s.)和刺柏属(Juniperus)之间互为姊妹分支,二者的共同祖先与HCX分互为姊妹分支[5]。这些类群之间的系统发育冲突可能源于快速谱系分化,一个祖先进化分支在350万年内快速分化成3个后代进化分支,因而导致了明显的不完全谱系分选;除此之外,种间或者进化分支间的杂交渐渗也可能是导致上述冲突的原因之一[5]。
此后,关于柏科和整个裸子植物的系统发育基因组学研究[15-16] 也支持了这一发现。这一系列研究终结了近20年来关于柏木属系统发育位置的争议,明确了柏木属应仅包含旧世界的物种,与刺柏属形成姊妹分支;而原先归类为(广义)柏木属的新世界类群应独立为美洲柏木属,与北美金柏属和金柏属种构成同一分支。这些成果不仅深化了对柏木属及相关属之间关系的理解,也为柏科植物分类提供了明确可靠的分子依据。2022年,南京林业大学杨永教授领衔的国际裸子植物研究团队,提出了裸子植物的新分类体系,进一步明确了柏木属、美洲柏木属、北美金柏属和金柏属的分类地位[1](表1)。
尽管在国际上对柏木属的定义和范围已逐渐形成统一的认识,但许多国际数据库仍沿用旧有的分类体系(表2)。例如,《全球生物物种名录》(Catalogue of Life)依然采用广义柏木属(Cupressus s.l.)的概念,未承认美洲柏木属(Hesperocyparis)。同时,该名录还根据Farjon等[17]在2002年的修订,将北美金柏划归至金柏属下。《世界自然保护联盟受威胁物种红色名录》(The IUCN Red List of Threatened Species)虽承认北美金柏属(Callitropsis)和金柏属(Xanthocyparis)的分类地位,但同样将美洲柏木属的种划归(广义)柏木属。而柏木属保护计划(Cupressus Conservation Project)则采用了Christenhusz等[13]在2011年提出的克氏裸子植物系统,将柏木属、美洲柏木属、北美金柏属、金柏属的物种全部统归于(克氏)柏木属(Cupressus sensu Christenhusz,图1)。这种分类上的差异和延迟更新反映了科学研究与实际应用之间的鸿沟,同时也凸显了生物多样性数据库及时更新和统一的重要性。为了确保科学研究成果能够被广泛理解和接受,有必要在全球范围内加强数据库的更新和分类体系的统一。这不仅有助于提高学术研究的准确性和效率,对于生物多样性的保护和管理亦十分重要。
2 柏木属的物种多样性
2.1 柏木属的物种数量
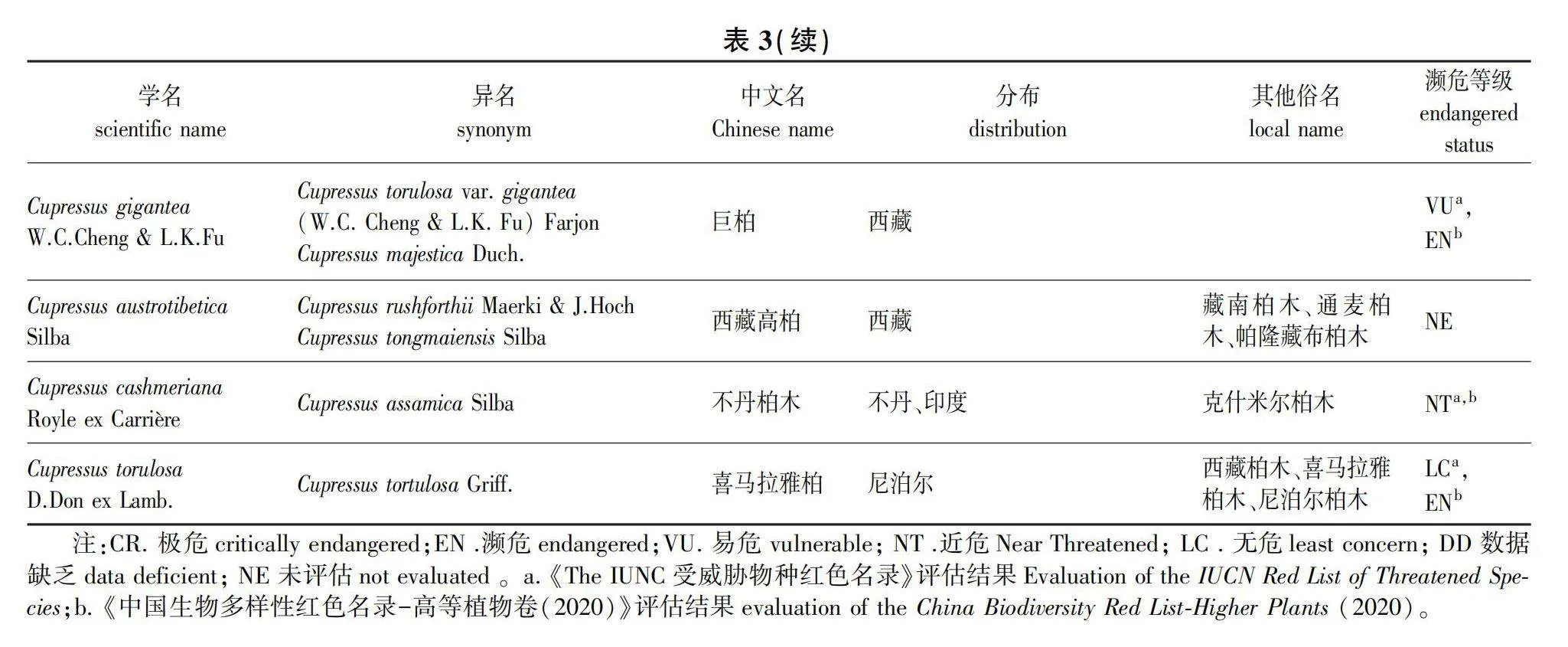
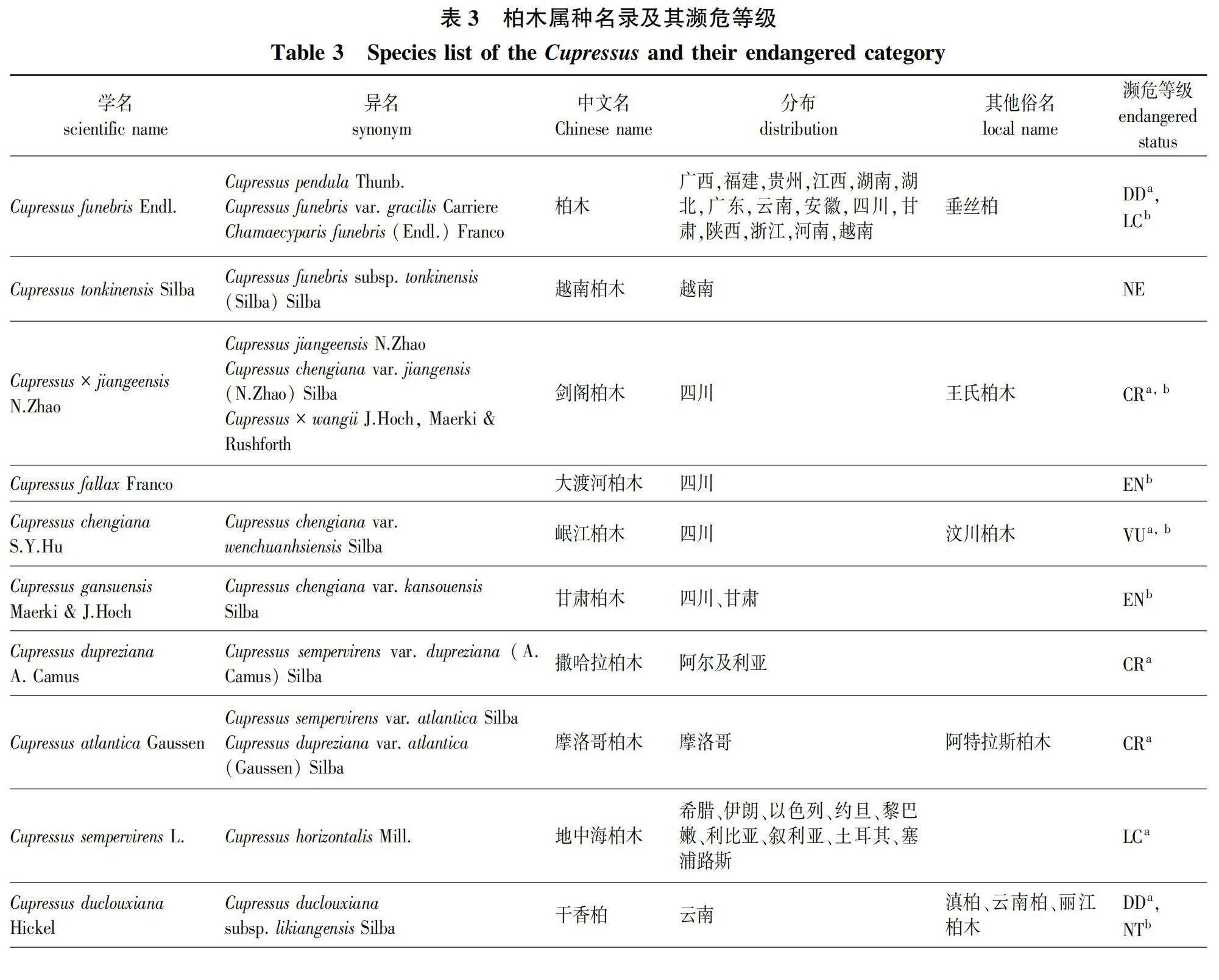
虽然近期的分子系统学研究揭示了柏木属物种主要分布于旧世界,但对该属下确切物种数量的共识尚未形成。据Farjon[8]在其2005年的专著中所述,柏木属共计15种。然而,仅7种来源于旧世界,属于狭义柏木属,包括:不丹柏木(C. cashmeriana Carrière)、岷江柏木(C. chengiana S.Y.Hu)、干香柏(C. duclouxiana Hickel)、撒哈拉柏木(C. dupreziana A.Camus)、柏木(C. funebris Endlicher)、地中海柏木(C. sempervirens L.)及喜马拉雅柏(C. torulosa D.Don ex Lamb.)。此外,剑阁柏木[C. chengiana var. jiangeensis (N. Zhao) Silba]、摩洛哥柏木[C. dupreziana var. atlantica (Gaussen) Silba]和巨柏[C. torulosa var. gigantea (W.C.Cheng amp; L.K.Fu) Farjon]在Farjon[8]的分类系统中均处理为变种,而非独立种。
除了这些被Farjon接受的种和变种之外,还有23个未被接纳的异名存在,尤其是喜马拉雅柏和不丹柏木分别有8个和6个异名,这些异名的存在不仅展示了柏木属植物形态学上的丰富变异,也反映了早期基于形态学特征进行物种鉴定时的复杂性和挑战。例如,美国学者Silba[19-20]在1988—1994年间,基于他在西藏林芝通麦镇帕隆藏布和易贡藏布两岸采集的柏木属样本的独特表型,描述了多个物种,包括C. austrotibetica Silba、C. tongmaiensis Silba及其变种C. tongmaiensis Silba var. ludlowii Silba,而Farjon[8]将上述种和变种均处理为为喜马拉雅柏(C. torulosa)的异名。
2006年,Little[10]在对广义柏木属进行修订时,认定狭义柏木属包含12种,这不仅包括Farjon[8]认定的7种来自旧世界的柏木属物种,还将Farjon(2005)视为变种的巨柏(C. gigantea W.C.Cheng amp; L.K.Fu)、剑阁柏木(C. jiangeensis N.Zhao)和摩洛哥柏木(C. atlantica)提升为种的级别;此外,他还将Silba[20]于1994年在越南发现的越南柏木(C. tonkinensis Silba)和1988年在西藏波密采集的西藏高柏(C. austrotibetica)提升为种的级别,尽管这两个种先前被Farjon[8]视为喜马拉雅柏(C. torulosa)的异名。2018年,Terry等[6]收集了Little[10](2006)认可的狭义柏木属12个种,同时包含了Silba[21](2005)认可的另外3个类群(C. assamica Silba、C. darjeelingensis (Silba) Silba和 C. chengiana var. kansouensis Silba),总计15个类群,重建了柏木属的系统发育关系。研究揭示了C. darjeelingensis 应归入美洲柏木属分支,而其余14个类群则分为3个进化分支。其中,C. assamica被Farjon(2005)[8]和Maerki (2017)[22]当作不丹柏木(C. cashmeriana)的异名处理,该研究结果表明C. assamica和C. cashmeriana聚在一支,支持上述处理方式。关于C. chengiana var. kansouensis,该种在核基因树上同C. chengiana var. chengiana聚为一支,而在叶绿体基因树上,二者并不构成单系群。因此,笔者建议可以根据核基因树的结果将C. chengiana var. kansouensis列为C. chengiana的变种,也可以根据质体树的结果将C. chengiana var. kansouensis独立成一个新种。然而,由于该研究所使用的片段较少,且未完全解决柏木属内各种之间的系统发育关系,因此在获得更有说服力证据之前,暂采用Little[10](2006)提出的关于狭义柏木属12个种的分类体系更为稳妥。
Hoch和Maerki等[23](2020)基于岷江柏木[C. chengiana(S.Y.Hu)]在不同流域间形态学特征的差异,对其进行了重新分类,将其分为3个不同的种。他们将Franco[24]在1969年描述的新种大渡河柏木(C. fallax Franco)重新界定为一个独立的种,这一种主要分布在四川大渡河流域,此前常被视为岷江柏木的一个异名。此外,将岷江柏木的定义缩减,仅包括岷江流域的种群;同时,将分布在甘肃白龙江流域的种群提升为一个新种,即甘肃柏木(C. gansuensis Maerki amp; J.Hoch),而并未采用Silba[19]在1994年提出的将其视为岷江柏木变种甘肃柏(C. chengiana var. kansouensis)的分类处理。笔者此前研究收集的群体基因组学证据表明,岷江柏木可以划分为分布在大渡河、岷江和白龙江流域的3个独立进化单元,其中岷江独立进化单元是大渡河独立进化单元和白龙江独立进化单元杂交产生的;3个独立进化单元的地理分布不重叠,且存在一定程度的生态位分化[25]。
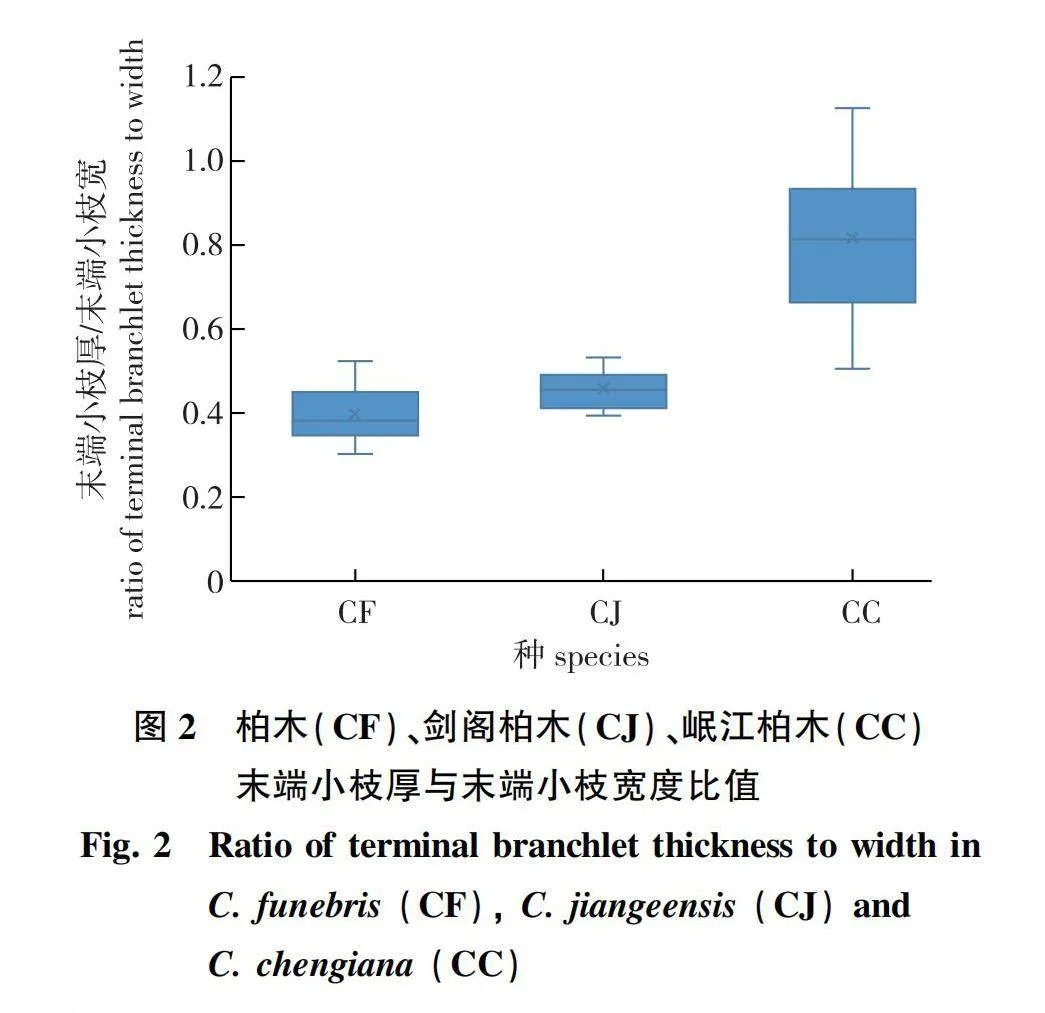
在同一年,Hoch和Maerki[23]根据剑阁柏木(C. jiangeensis)的球果种鳞通常为6对,并结合Xu等[26]在群体水平上的叶绿体基因单倍型分析结果,认为剑阁柏木并不是一个独立种,而是将其视为大渡河柏木(C. fallax Franco)的一个异名。同时,基于从剑阁柏木采集的种子萌发的个体,他们命名了一个新的杂交种,王氏柏木(Cupressus ×wangii J.Hoch, Maerki amp; Rushforth)[23]。然而,根据笔者的野外观察和形态学测量,剑阁柏木同大渡河柏木存在明显的差异:首先,虽然剑阁柏木的球果为椭圆形、具有6对种鳞,更类似于大渡河柏木,但其球果大小却相对较小;其次,剑阁柏木的末端小枝更加扁平,更类似于柏木(图2)。鉴于剑阁柏木在形态上介于柏木和大渡河柏木之间,笔者认为剑阁柏木更可能是前二者的杂交1代个体,应将其命名为Cupressus × jiangeensis N.Zhao。而王氏柏木可能是剑阁柏木与柏木的回交后代,应视为剑阁柏木的异名。
2023年5月,在中国西藏林芝波密县发现了一株高达102.3 m的柏木属植株,不同的媒体在报道时采取了“西藏柏木”和“藏南柏木”两种说法。这引发了之前被忽略的问题:在中国是否有“西藏柏木(C. torulosa)”的分布?Cupressus torulosa 最初于1824年被发现与命名,模式标本采集自尼泊尔。基于形态特征,此后发现于中国西藏东部及南部的柏木属物种被鉴定为C. torulosa。据《中国植物志》记载,该种首次以“喜马拉雅柏”的中文名出现在1937年陈嵘主编的《中国树木分类学》中;1961年《中国树木学》将其中文名定为“西藏柏木”,以反映其在中国西藏分布;1975年《植物分类学报》中,该种被称为“喜马拉雅柏木”;而到了1978年,《中国植物志》第7卷则使用“西藏柏木”作为其中文名[2],此后一直沿用此中文名。1988年,美国学者Silba[18]基于在西藏波密通麦帕隆藏布和易贡藏布沿岸采集柏木属植物标本,将其命名为新种C. austrotibetica。2006年,Little[10]在对柏木属进行系统发育分析时,发现C. austrotibetica与不丹柏木(C. cashmeriana)关系更为接近。2010年,Xu等[26]利用群体数据进行叶绿体单倍型分析,结果显示C. austrotibetica与不丹柏木共享相同单倍型,与Cupressus torulosa在遗传上存在差异。这表明在中国境内实际未分布Cupressus torulosa[27],原本在西藏东部和东南部分布,鉴定为C. torulosa的柏木属植物实为C. austrotibetica。因此,继续使用“西藏柏木”作为C. torulosa的中文名已不再合适,建议恢复其在中国最初的中文名,即“喜马拉雅柏”,以避免分类上的混淆。同时,考虑到“藏南”一词在国内容易与藏南地区混淆,而且该种主要分布在波密县,因此建议将C. austrotibetica的中文名改为可以同时反映其分布地和物种特征的“西藏高柏”。
在2021年,Maerki等[28]基于在内斯植物园中种植的由Rushforth在西藏林芝通麦镇附近不同地点采集的柏木属植物扦插繁殖植株表现出的表型性状变异模式,提出了一个新种的分类建议。他们注意到,从通麦镇以西的易贡藏布河沿岸采集的样本表现出典型的西藏高柏(C. austrotibetica)特征,而从通麦镇以东采集的样本在生境和叶片形态上与已知的西藏高柏类群存在差异。基于这些观察,他们将后者认定为一个新种,命名为Cupressus rushforthii Maerki amp; J.Hoch。虽然该种被中国生物物种名录收录时翻译成通麦柏木,但通麦柏木最早在公开发表的学术文献中指C. tongmaiensis[27]。为避免中文名的混淆,建议C. rushforthii的中文名采用帕隆藏布柏木。
虽然帕隆藏布柏木新种的发表为柏木属分类研究做出了贡献,但这一新种的界定主要依据的是在植物园中栽培的扦插繁殖个体,而未通过野外收集的、群体水平的形态或分子证据进行进一步的验证。此外,在Maerki等[28]利用RAPD技术构建的系统发育树中,新种C. rushforthii与C. austrotibetica聚在同一个单系分支上,显示的差异不足以支持将其上升到种的级别。上述研究引发出一系列问题:Maerki和Hoch所观察到的形态差异是否仅为个体变异?在群体层面上,C. rushforthi是否与C. austrotibetica有明显的形态差异?二者是否存在物种水平的遗传分化?目前这些问题仍然悬而未决。因此,在获取更多充分的证据支持C. rushforthii的种级地位之前,建议将其视为C. austrotibetica的一个异名。
因此,根据目前国内外学者对柏木属的研究现状,综合考虑形态、遗传、生态位分化等,笔者建议的柏木属分类包含14种(表3):柏木(C. funebris)、越南柏木(C. tonkinensis)、剑阁柏木(C.×jiangeensis)、大渡河柏木(C. fallax)、岷江柏木(C. chengiana)、甘肃柏木(C. gansuensis)、撒哈拉柏木(C. dupreziana)、摩洛哥柏木(C. atlantica)、地中海柏木(C. sempervirens)、干香柏(C. duclouxiana)、巨柏(C. gigantea)、西藏高柏(C. austrotibetica)、不丹柏木(C. cashmeriana)以及喜马拉雅柏(C. torulosa)。
2.2 分类处理
2.2.1 西藏高柏 藏南柏木、通麦柏木、帕隆藏布柏木
Cupressus austrotibetica Silba in Phytologia 65: 334 (1988); Cupressus duclouxiana subsp. austrotibetica (Silba) Silba in J. Int. Conifer Preserv. Soc. 12: 62 (2005). Type: China (中国). Xizang (西藏), Pome (波密), Trulung (排龙), more numerous up the river, in mixed forest, 2134 m, 13 Jan. 1947, Ludlow, Sherriff amp; Elliot 12141 (Holotype: BM000546888; Isotype: E00188326).
Cupressus tongmaiensis Silba in J. Int. Conifer Preserv. Soc. 1: 24 (1994); Cupressus gigantea subsp. tongmaiensis (Silba) Silba in J. Int. Conifer Preserv. Soc. 12: 71 (2005); Cupressus austrotibetica subsp. tongmaiensis (Silba) Silba in J. Int. Conifer Preserv. Soc. 20: 28 (2013). Type: China (中国). Xizang (西藏), Pome (波密), Tongmai (通麦), Yiong Tsangpo river valley (易贡河谷), on eastern forest slopes, 2070 m, 23 Jul. 1965, Hong, D.Y amp; Ying T.S (洪德元和应俊生) 650772 (Lectotype: PE00012995; Holotype: PE00012995, PE00012996).
Cupressus austrotibetica subsp. medogensis Silba in J. Int. Conifer Preserv. Soc. 20: 27 (2013). Type: China (中国). Xizang (西藏), Medog (墨脱), Bangxin township (旁辛乡), next to the house, 1560m, 26 Nov. 1982, Cheng S.Z. amp; Li B.S. (程树志和李渤生)3075 (Lectotype: PE00026755: Isolectotype PE00026755).
Cupressus rushforthii Maerki amp; J.Hoch in Bull. Cupressus Conservation Proj. 10(2): 57 (2021). Type: Tree cultivated at Ness Botanic Garden from KR6015 clone B; China (中国), Xizang (西藏), Bome County (波密), Parlung Tsangpo (帕隆藏布), above the confluence at Tongmai, close to Tangdui (30°06′07.16″N, 95°06′15.41″E), 2100 m, T.Baxter s.n. (Holotype: LIV-2005.15.2873)
分布:产于西藏林芝通麦镇附近易贡藏布和帕隆藏布沿岸。
2.2.2 剑阁柏木 王氏柏木
Cupressus× jiangeensis N.Zhao in Acta Phytotax. Sin. 18: 210 (1980); Cupressus chengiana var. jiangeensis (N.Zhao) Silba in Phytologia 49: 394 (1981); Cupressus chengiana subsp. jiangeensis (N.Zhao) Silba in J. Int. Conifer Preserv. Soc. 12: 58 (2005). Type: China (中国), Sichuan (四川), Jiange (剑阁), Hanyang (汉阳), 840 m, 28 Sep. 1978, Cai amp; Min (蔡霖生和闵通知) 101-104(Isotype: PE00002531).
Cupressus × wangii J.Hoch, Maerki amp; Rushforth in Bull. Cupressus Conservation Proj. 9: 20 (2020). Type: USA, California, Santa Cruz Co., cultivated from seeds of the type tree of C. × jiangeensis, 3 Jan. 2003, Silba B-330 (Holotype: NY00658653).
分布:产于四川剑阁汉阳区翠云廊景区内,目前仅发现1棵。
3 展望与建议
柏木属的分类地位近年来引起了较大争议,主要原因有两点。一方面,柏木属分类地位的争议主要在于狭义柏木属和美洲柏木属的形态特征区分度不高,因此基于形态特征开展分类工作的传统分类学者往往倾向于采用广义柏木属的概念。另一方面,虽然基于核基因序列和全证据数据的系统发育分析研究表明,狭义柏木属和刺柏属为姊妹分支[7, 10],但是基于叶绿体基因片段构建的系统发育树支持狭义柏木属、金柏属、美洲金柏属和美洲柏木属形成一个单系类群,为传统分类学者提供了系统发育证据上的支持[11-12]。近年来,基于高通量测序获取的核基因组数据开展的系统发育基因组分析反复证明广义柏木属是并系群,狭义柏木属的姊妹分支是刺柏属,柏木属分类地位的争议才尘埃落定[5, 15-16]。随着高通量测序数据在系统发育分析中的不断推广,相信会有更多类似于柏木属分类地位争议的悬案得以解决。
除此之外,对于狭义柏木属来说,其包含的物种数量近年来也存在争议。究其原因,首先是因为该属物种在形态上极为相似,近缘种难以依据明确的形态学特征进行准确区分。其次,不同学者对种的定义的理解存在差异,导致基于形态学差异描述的许多新种存在争议。再者,某些柏木属物种,如西藏高柏、喜马拉雅柏、摩洛哥柏木等,分布区域非常有限且地理位置偏远,开展野外调查难度大、收集的标本量偏小。此外,一些柏木属种如柏木因长期被广泛栽培,其原始产地和自然分布地难以追溯,缺乏地理分布信息,进一步增加了理解该属内种间关系的复杂度[6]。最后,柏木属内的种间杂交非常频繁,频繁的杂交导致杂交后代呈现出复杂的表型[25,29],有时可能导致杂交后代被错误鉴定为新物种。
正确认识物种是开展生物多样性研究的基础,物种划分的科学性和合理性关系到生物多样性的研究、保护和可持续利用[30]。进入21世纪以来,尽管分类学者们逐渐认识到单凭形态的差异不足以正确认识物种,发表新种时应至少符合两个物种概念的条件,如是否在形态上具有明显间断、是否在遗传上聚为单系、是否具有独特的生态位等[31]。然而,从柏木属的分类学历史来看,很多新种的发表仍然主要依据少量标本来源的形态学证据,忽略对野外种群的调查对比,因此往往很难区分种群内性状变异(包括个体间差异、表型可塑性等)和种间稳定性状差异。因此,在未来对柏木属等进化历史复杂类群的物种进行划分和描述时,除了形态学差异之外,应更多地结合遗传、生态和生物地理等多维度的数据,以确保物种划分的准确性和科学性。同时,建议只有在分子证据和形态学证据等证明具有群体水平上的差异时,才做分类等级调整,这样可以减少不可靠新物种或新类群出现,从而减少分类处理上的混乱。在未来的研究中,随着高通量测序技术为代表的多组学技术的普及和成本的降低,系统分类与进化研究有望获取更全面的基因组学、转录组学、蛋白组学、表型组学数据,从而极大地促进人类对生物类群的起源、演化和系统分类的认知,进而更好地保护和利用生物多样性,促进人与自然和谐共生。
致谢
刘建全、杨亲二、杨永在论文撰写过程中的建设性建议。
参考文献(reference):
[1]YANG Y,FERGUSON D K,LIU B,et al.Recent advances on phylogenomics of gymnosperms and a new classification[J].Plant Divers,2022,44(4):340-350.DOI: 10.1016/j.pld.2022.05.003.
[2]郑万均, 傅立国. 中国植物志 (第七卷) [M]. 北京: 科学出版社, 1978.
[3]REN Y,LI C,CHAU K,et al.Conserving the primary forests in the Yarlung Tsangpo Grand Canyon for people and nature[J].Nat Ecol Evol,2024,8(5):837-839.DOI: 10.1038/s41559-024-02383-y.
[4]FARAHMAND H. The genus Cupressus L [J]. Horticultural Reviews, 2020: 213-287.
[5]MAO K S,RUHSAM M,MA Y Z,et al. A transcriptome-based resolution for a key taxonomic controversy in Cupressaceae[J].Ann Bot,2019,123(1):153-167.DOI: 10.1093/aob/mcy152.
[6]TERRY R G,SCHWARZBACH A E,BARTEL J A. A molecular phylogeny of the Old World cypresses (Cupressus:Cupressaceae):evidence from nuclear and chloroplast DNA sequences[J].Plant Syst Evol,2018,304(10):1181-1197.DOI: 10.1007/s00606-018-1540-1.
[7]ADAMS R P,BARTEL J A,PRICE R A,et al.A new genus,Hesperocyparis,for the cypresses of the western hemisphere (Cupressaceae)[J].Phytologia,2009,91(1):160-185.
[8]FARJON A.A monograph of Cupressaceae and Sciadopitys[M].Richmond,Surrey,UK:Royal Botanic Gardens,Kew,2005.
[9]FU L, YU Y F, ADAMS R P, et al. Cupressaceae [J]. Flora of China, 1999, 4: 62-77.
[10]LITTLE D P.Evolution and circumscription of the true cypresses (Cupressaceae:Cupressus)[J].Syst Bot,2006,31(3):461-480.DOI: 10.1600/036364406778388638.
[11]MAO K S,MILNE R I,ZHANG L B,et al. Distribution of living Cupressaceae reflects the breakup of Pangea[J].Proc Natl Acad Sci USA,2012,109(20):7793-7798.DOI: 10.1073/pnas.1114319109.
[12]MAO K S,HAO G,LIU J Q,et al. Diversification and biogeography of Juniperus (Cupressaceae):variable diversification rates and multiple intercontinental dispersals[J].New Phytol,2010,188(1):254-272.DOI: 10.1111/j.1469-8137.2010.03351.x.
[13]CHRISTENHUSZ M J M,REVEAL J L,FARJON A,et al. A new classification and linear sequence of extant gymnosperms[J].Phytotaxa,2011,19(1):55.DOI: 10.11646/phytotaxa.19.1.3.
[14]ZHU A D,FAN W S,ADAMS R P,et al. Phylogenomic evidence for ancient recombination between plastid genomes of the Cupressus-Juniperus-Xanthocyparis complex (Cupressaceae)[J].BMC Evol Biol,2018,18(1):137.DOI: 10.1186/s12862-018-1258-2.
[15]LIU X Q,XIA X M,CHEN L,et al. Phylogeny and evolution of Cupressaceae:updates on intergeneric relationships and new insights on ancient intergeneric hybridization[J].Mol Phylogenet Evol,2022,177:107606.DOI: 10.1016/j.ympev.2022.107606.
[16]STULL G W,QU X J,PARINS-FUKUCHI C,et al. Gene duplications and phylogenomic conflict underlie major pulses of phenotypic evolution in gymnosperms[J].Nat Plants,2021,7(8):1015-1025.DOI: 10.1038/s41477-021-00964-4.
[17]FARJON A,HIEP N T,HARDER D K,et al. A new genus and species in Cupressaceae (Coniferales) from northern Vietnam,Xanthocyparis vietnamensis[J].Novon,2002,12(2):179.DOI: 10.2307/3392953.
[18]LITTLE D P,SCHWARZBACH A E,ADAMS R P,et al. The circumscription and phylogenetic relationships of Callitropsis and the newly described genus Xanthocyparis (Cupressaceae)[J].Am J Bot,2004,91(11):1872-1881.DOI: 10.3732/ajb.91.11.1872.
[19]SILBA J. A new species of Cupressus L. from Tibet (Cupressaceae) [J]. Phytologia,1988, 65(5): 333-336.
[20]SILBA J. The transpacific relationship of Cupressus in India and North America [J]. J Int Conifer Preserv. Soc, 1994(1): 1-28.
[21]SILBA J. A monograph of the genus Cupressus L. in the twenty-first century[J]. J Int Conifer Preserv Soc, 2005(12): 30-103.
[22]MAERKI D. Note on Cupressus assamica Silba [J]. Bull Cupressus Conserv Proj, 2017, 6: 74-6.
[23]HOCH J, MAERKI D. About Cupressus jiangeensis N.Zhao [J]. Bull of Cupressus Conserv Proj, 2020, 9(2): 15-22.
[24]FRANCO J A. On Himalayan-Chinese cypresses[J]. Portugaliae, Acta Biol Ser B, 1969(9):183-185.
[25]LI J L,MILNE R I,RU D F,et al. Allopatric divergence and hybridization within Cupressus chengiana (Cupressaceae),a threatened conifer in the northern Hengduan Mountains of Western China[J].Mol Ecol,2020,29(7):1250-1266.DOI: 10.1111/mec.15407.
[26]XU T T,ABBOTT R J,MILNE R I,et al. Phylogeography and allopatric divergence of cypress species (Cupressus L.) in the Qinghai-Tibetan Plateau and adjacent regions[J].BMC Evol Biol,2010,10:194.DOI: 10.1186/1471-2148-10-194.
[27]杨永,檀超,杨智.从《国家重点保护野生植物名录》看我国裸子植物保护[J].生物多样性,2021,29(12):1591-1598.YANG Y,TAN C,YANG Z.Conservation of gymnosperms in China:perspectives from the list of national key protected wild plants[J].Biodivers Sci,2021,29(12):1591-1598.DOI: 10.17520/biods.2021403.
[28]MAERKI D, HOCH J.Cupressus rushforthii, a new cypress species in Xizang, China, with an introduction on the Chinese cypresses and a survey of C. austrotibetica [J]. Bulletin of the Cupressus Conservation Project, 2021, 10(2): 55-89.
[29]MA Y Z,WANG J,HU Q J,et al. Ancient introgression drives adaptation to cooler and drier mountain habitats in a cypress species complex[J].Commun Biol,2019,2:213.DOI: 10.1038/s42003-019-0445-z.
[30]洪德元. 生物多样性事业需要科学、可操作的物种概念[J].生物多样性,2016,24(9):979-999.HONG D Y.Biodiversity pursuits need a scientific and operative species concept[J].Biodivers Sci,2016,24(9):979-999.DOI: 10.17520/biods.2016203.
[31]刘建全. “整合物种概念” 和“分化路上的物种”[J].生物多样性,2016,24(9):1004-1008.LIU J Q.“The integrative species concept” and “species on the speciation way”[J].Biodivers Sci,2016,24(9):1004-1008.DOI: 10.17520/biods.2016222.
(责任编辑 吴祝华)
猜你喜欢
环球人文地理·评论版(2016年5期)2017-01-03 03:19:00
农家科技下旬刊(2016年9期)2016-12-15 13:55:30
农业与技术(2016年20期)2016-12-08 21:26:55
农业与技术(2016年15期)2016-11-09 16:57:47
河北渔业(2016年9期)2016-11-05 10:22:38
现代交际(2016年14期)2016-08-15 15:23:21
科技视界(2016年4期)2016-02-22 08:57:24
南方农业·下旬(2015年4期)2015-06-24 19:55:19
建材发展导向(2015年2期)2015-04-16 09:37:33
