
外源独角金内酯对铅胁迫下番茄幼苗生理特性的缓解效应
2024-05-29 13:56:27史沉鱼韦芳灵黄彩腰邓晰朝
中国瓜菜 2024年5期
关键词:生理指标
史沉鱼 韦芳灵 黄彩腰 邓晰朝
摘 要:为了研究外源独角金内酯(GR24)对重金属铅胁迫下番茄幼苗生长生理的影响,以番茄品种上海红宝石908为材料,在河池学院植物生理材料培养室进行试验,培养室平均温度28 ℃,平均相对湿度为70.85%。选择生长状态较为一致且为3叶1心的番茄幼苗移栽至1/2 Hoagland营养液中,试验共设4个处理,CK(完全营养液)、GR24 (完全营养液+50 nmol·L-1 GR24)、Pb (完全营养液+200 mg·L-1 Pb)、Pb+GR24 (完全营养液+200 mg·L-1 Pb +50 nmol·L-1 GR24)。研究了GR24对正常生长和铅胁迫下番茄幼苗的长势、抗氧化系统和渗透调节物质等的影响。结果表明,与CK相比,200 mg·L-1的Pb胁迫处理番茄幼苗的株高、根长、单叶面积分别降低了21.03%、35.15%、27.27%。与单独Pb胁迫处理相比,施加外源GR24可以明显提高番茄幼苗叶片中叶绿素、类胡萝卜素、可溶性糖、可溶性蛋白含量(SP)和生物积累量;显著提高POD、CAT等抗氧化酶活性和根系活力;降低植株叶片H2O2、[O2[-·]]、脯氨酸和MDA含量。综上,外源施加50 nmol·L-1的GR24能加快铅胁迫下番茄幼苗叶绿素合成速度,提高叶片抗氧化能力,促进番茄苗期生长,缓解铅胁迫对番茄幼苗生长的毒害作用。
关键词:番茄幼苗;铅胁迫;GR24;生理指标;缓解效应
中图分类号:S641.2 文献标志码:A 文章编号:1673-2871(2024)05-115-09
Effects of exogenous on the physiological characteristics of tomato seedlings under Pb stress
SHI Chenyu, WEI Fangling, HUANG Caiyao, DENG Xichao
(Guangxi Key Laboratory of Sericulture Ecology and Applied Intelligent Technology/Guangxi Collaborative Innovation Center of Modern Sericulture and Silk/Guangxi Colleges Universities Key Laboratory of Exploitation and Utilization of Microbial and Botanical Resources/Institute of Chemical and Biological Engineering, Hechi University, Hechi 546300, Guangxi, China)
Abstract:In order to study the effect of exogenous GR24 on the growth physiology of tomato seedlings under heavy metal Pb stress, the tomato variety Shanghai Ruby 908 was used as the experimental material. The experiment was conducted in a plant physiological material culture room, with an average temperature of 28 ℃ and an average relative humidity of 70.85%. Tomato seedlings with consistent growth status and three leaves and one heart were transplanted into 1/2 Hoagland nutrient solution. The experiment included four treatments: CK (complete nutrient solution), GR24 (complete nutrient solution+50 nmol?L-1 GR24), Pb (complete nutrient solution+200 mg?L-1Pb), and Pb+GR24 (complete nutrient solution+200 mg?L-1+50 nmol?L-1 GR24). The results showed that the Pb stress of 200 mg?L-1 reduced the plant height, root length, and single leaf area of tomato seedlings by 21.03%, 35.15%, and 27.27%, respectively. Compared with individual Pb stress treatment, applying exogenous GR24 can significantly increase the content of chlorophyll, carotenoids, soluble sugars, soluble proteins, and bioaccumulation in tomato seedling leaves; significantly increase the activity of antioxidant enzymes such as SOD, POD, CAT, and root activity; reduce the content of H2O2, O2-, proline and MDA in leaf. Therefore, exogenous application of 50 nmol?L-1 GR24 can alleviate the toxic effect of Pb stress on the growth of tomato seedlings.
Key words: Tomato seedlings; Pb stress; GR24; Physiological indicators; Alleviative effect
近年来,我国农业环境受重金属污染的现象屡见不鲜,据报道,中国总耕地面积将近有1/5遭受重金属污染,0.12亿t粮食被重金属毒害[1]。在众多重金属种类中铅是污染面积最大的重金属之一,极易累积在动植物体内,对人体组织器官和植物的生长发育造成损伤[2]。国内外研究发现,在铅胁迫环境下,植物形态会发生变化,同时能引起植物地上部分缓慢生长,根系生物量减少、根系活力降低、细胞膜透性急剧增加、保护酶系统修复能力减弱等[3]。在高浓度铅胁迫下,油菜种子活力降低,幼苗质膜受损,当铅浓度超过一定限度时,POD和SOD活性呈下降的趋势[4]。在铅胁迫下,绿豆种子活力、发芽率、发芽势以及根长、芽长和幼苗鲜质量、幼苗根冠比等指标明显表现为随铅浓度升高而降低[5]。铅胁迫下常春藤生长会受到刺激,生长缓慢;高濃度铅胁迫下,会出现植株徒长、节间距拉长、生长异常等情况[6]。由此可见,研究铅对植物的毒害机制及如何缓解植物的铅毒害效应,保证作物质量安全具有重要意义。
独脚金内酯(SLs)是独脚金醇类化合物及其衍生物的总称,是近年来发现的一类能调控植物发育过程的信号分子,具有促进种子萌发、刺激丛枝菌根有丝分裂和生长、调控植物分枝等多种功能[7]。天然独脚金内酯主要包括独脚金醇(strigol)、列当醇(orobanchol)及其衍生物、高粱内酯(sorgolactone)等。人工合成的独脚金内酯主要是独脚金醇类似物(germination re-leaser,GR),如GR24、GR7、GR6、GR3等,其中GR24活性最高,常被用作独脚金内酯信号通路相关研究中的常规性参照物[8]。GR24能够调控植物的形态结构、生殖发育并且在植物抵御如水涝、干旱、弱光以及重金属镉、高盐等胁迫的过程中也发挥了举足轻重的作用[9-11]。有研究表明,独脚金内酯能在基因水平与ABA相互作用调控乌桕干旱和盐渍胁迫[12]。外源添加独脚金内酯处理后能显著减轻镉对大麦光合作用的抑制[13]。
番茄在蔬菜栽培中占重要地位,是我国种植面积最大、最具象征性的无土栽培作物之一[14]。有研究表明,高浓度的重金属会抑制番茄幼苗的生长,表现为种子萌发率、株高明显降低,幼苗根系明显生长不良等[15]。独角金内酯作为新型植物激素,在调控植物生长生理及抗性生理方面的研究已有相关报道[10-13],但独角金内酯能否缓解铅对番茄的生理毒害还不十分清楚。笔者以重要蔬菜栽培种类和研究模式作物番茄为试验材料,探究了铅胁迫对番茄幼苗生理特性的影响,并初步探索了独角金内酯(GR24)在缓解番茄铅遭受铅胁迫方面的生理效应,以期为实际生产中通过施用外源GR24提高番茄对重金属胁迫的耐受性提供理论指导。
1 材料与方法
1.1 材料
试验材料为上海红宝石908番茄,属早熟品种,生长势强、适应范围、耐黄瓜花叶病毒,是较抗病的番茄品种。独角金内酯购自广西卓一生物技术有限公司。
1.2 试验设计
1.2.1 单独铅胁迫处理 试验于2021年2月至2022年7月在河池学院植物生理材料培养室进行,培养室平均温度28 ℃,平均相对湿度为70.85%。试验材料做如下处理:先用40 ℃温水浸种3 h,捞出擦干后再用1% KMnO4消毒10 min,消毒后用蒸馏水冲洗3~4次,将种子放入底部铺有两层滤纸的干净玻璃培养皿进行催芽,每皿20粒,加入20 mL蒸馏水,种子萌发期间每日补充少量蒸馏水溶液以保持滤纸湿润,待种子萌发后转移到土壤中进行培养。对预培养7 d的番茄幼苗进行胁迫处理,分别用不同质量浓度(0、50、100、200、300、400 mg·L-1)的硝酸铅溶液进行处理,试验采用完全随机设计,共设6个处理,每个处理3次重复,每次18个样本。处理6 d后选取健壮的番茄幼苗洗净,擦干,测量根长、株高、真叶数、单叶面积等形态指标,从6个处理中筛选出合适的铅胁迫浓度进行下一步试验。
1.2.2 外源施加GR24处理 将番茄种子按照上述方法预培养7 d后,选择生长状态较为一致的3叶1心期番茄幼苗移栽至1/2 Hoagland营养液中,进行Pb胁迫和施加外源GR24(浓度依据前期预试验结果)处理,浓度参照相关研究[13]。铅(Pb)以硝酸铅溶液形式加入,试验共设4个处理:CK(完全营养液)、GR24(完全营养液+50 nmol·L-1 GR24)、Pb(完全营养液+200 mg·L-1 Pb)、Pb+GR24 (完全营养液+200 mg·L-1 +50 nmol·L-1 GR24)。每瓶放1株,每个处理设20瓶(玻璃瓶容量350 mL),3次重复。处理20 d后进行番茄幼苗外观形态以及相关生理指标的测定。
1.3 测定指标及方法
采用丙酮-乙醇提取法测定叶绿素和类胡萝卜素含量[16];采用蒽酮比色法测定可溶性糖含量[16];采用考马斯亮蓝法测定可溶性蛋白含量[17];采用磺基水杨酸法测定游离脯氨酸含量[16];采用愈创木酚法测定POD活性[18];采用硫代巴比妥酸比色法测定MDA含量[19];采用氮蓝四唑光化学还原法测定超氧化物歧化酶(SOD)活性[16];采用紫外吸收法测定CAT含量[16]。采用叶片化学染色法,根据染色结果判断超氧阴离子自由基[O2[-·]]积累量[19],采用直接测量法测定主根长度与侧根数量[20],采用TTC法测定根系活力[21],测定株高、单叶面积、单株真叶数、地上地下部干鲜质量[22]。以上生理指标测定每个处理测定15枚叶片,3次重复。
1.4 数据分析
采用Excel(2016)软件作图,同时运用SPSS 18. 0软件进行单因素方差分析,采用Ducans测验在0.05水平上进行显著性分析,对GR24、Pb胁迫以及二者的互作效应进行双因素方差分析。
2 结果与分析
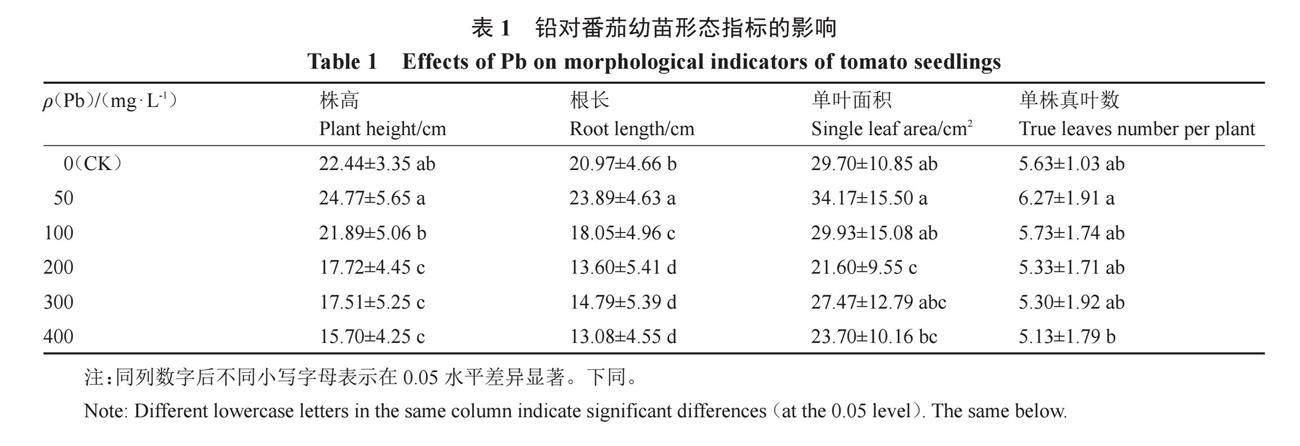
2.1 外源Pb对番茄幼苗形态指标的影响
由表1可知,与对照相比,番茄幼苗铅处理质量浓度≥100 mg·L-1时,株高均表现为下降,铅质量浓度≥200 mg·L-1时各处理株高均与对照存在显著差异;当铅质量浓度为400 mg·L-1时,番茄幼苗株高最小,比对照组降低30.04%。当铅质量浓度为50 mg·L-1时番茄根长显著高于对照,增加了10.38%,此时的铅浓度能够促进番茄幼苗根的伸长;其余浓度铅胁迫下根长与对照组相比均显著下降,分别降低了13.92%、35.15%、29.47%、37.63%,表明高浓度铅处理对番茄幼苗根的伸长表现为抑制作用。仅当铅质量浓度为200 mg·L-1时番茄单叶面积与对照存在显著差异,此浓度单叶面积最小,比对照下降了27.27%;其他各浓度铅处理对单叶面积的影响与对照组相比无显著差异。各浓度铅处理番茄单株真叶数均与对照无显著差异。
综合幼苗的生长情况,后续试验模拟铅胁迫对番茄幼苗的生理响应选取Pb处理的质量浓度为200 mg·L-1。
2.2 外源GR24對铅胁迫下番茄幼苗生理指标的影响
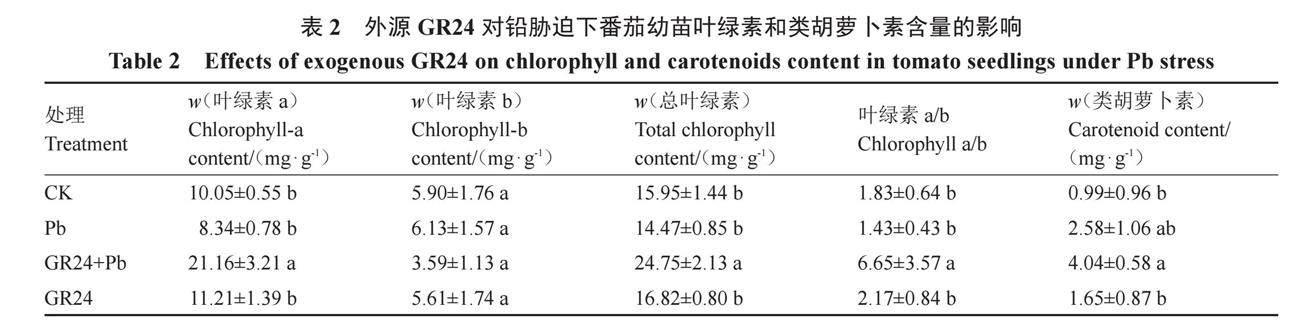
2.2.1 外源GR24对铅胁迫下番茄幼苗叶绿素和类胡萝卜素含量的影响 由表2可知,与CK相比,铅胁迫使番茄叶片中的叶绿素a、总叶绿素含量和叶绿素a/b分别降低了17.01%、9.28%和21.86%,但均与对照无显著差异。与铅胁迫相比,在重金属铅胁迫下添加GR24,叶片中的叶绿素a、总叶绿素含量和叶绿素a/b水平均显著增加,类胡萝卜素含量增加,但二者间差异不显著。在无铅胁迫条件下,GR24处理相较于对照,番茄叶片中的叶绿素a含量、总叶绿素含量、叶绿素a/b、类胡萝卜素含量水平总体上有所增加,但差异并不显著。
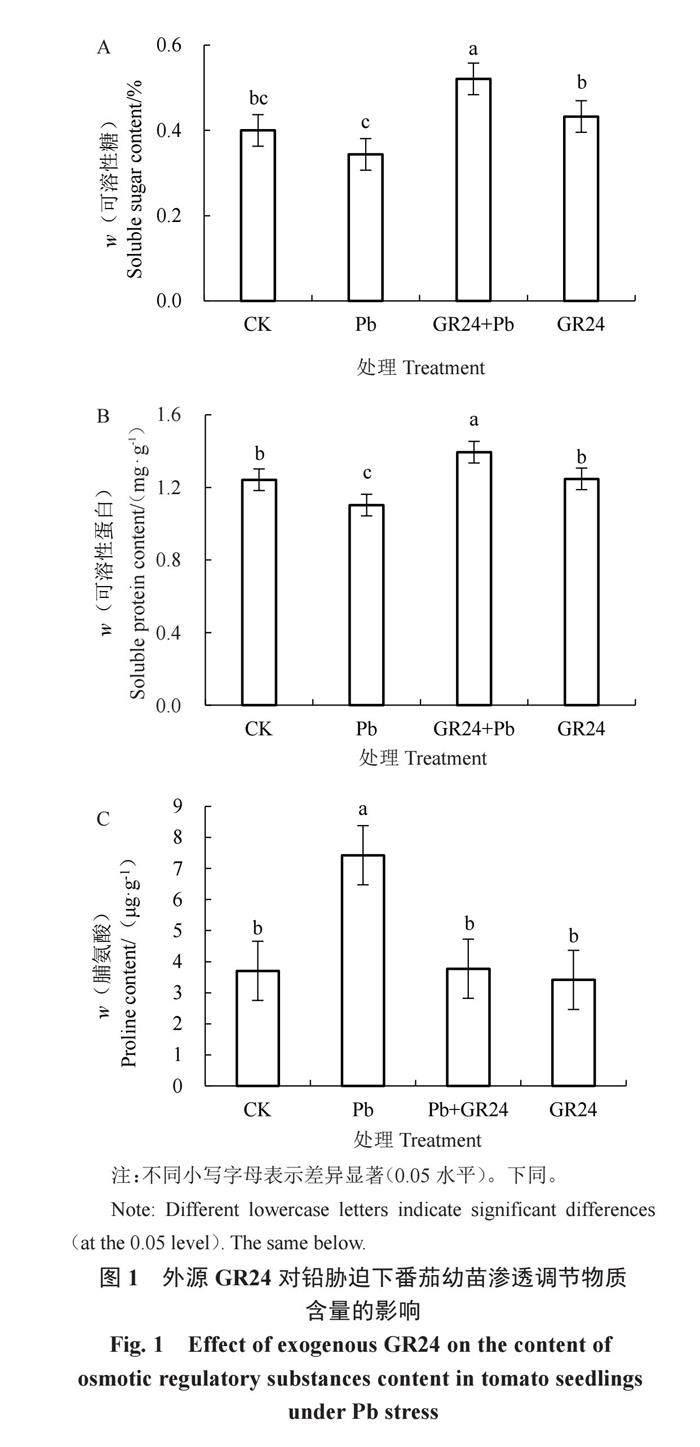
2.2.2 外源GR24对铅胁迫下番茄幼苗渗透调节物质含量的影响 从图1可以看出,铅处理下番茄幼苗可溶性糖含量与对照相比虽没有达到显著差异水平但明显降低。说明铅胁迫可抑制植物体内可溶性糖的合成。添加外源GR24后,可溶性糖含量与对照存在显著差异,增加了30.25%;单独施加GR24番茄幼苗的可溶性糖含量略高于对照,二者间无显著差异。在铅胁迫下添加GR24可促进番茄植株可溶性糖的合成,降低细胞内的渗透势,使番茄幼苗能够表现出更高的耐铅性。铅胁迫处理幼苗的可溶性蛋白含量(SP)显著低于对照;铅胁迫下施加外源GR24处理番茄幼苗SP含量显著高于对照和铅胁迫处理,相比对照提高了12.23%。与对照相比,在铅胁迫下对番茄幼苗施加外源GR24可促进幼苗SP含量提高来适应铅胁迫环境。与对照相比,在铅胁迫下,番茄幼苗叶片中的脯氨酸含量显著增加了1倍;在铅胁迫下施用外源GR24,使幼苗脯氨酸含量比铅胁迫处理下降49.19%,但与对照相比没有达到显著水平。在单独施用GR24时,幼苗脯氨酸含量略低于对照,二者无显著差异。
2.2.3 外源GR24对铅胁迫下番茄幼苗几种抗氧化酶活性的影响 由图2可知,铅胁迫下番茄幼苗细胞内的POD活性与对照相比显著下降29.80%;铅胁迫下施加外源GR24后,POD活性显著升高,且显著高于对照。表明适宜浓度的GR24可以提高番茄幼苗的耐铅性,可以有效缓解番茄幼苗的铅毒害作用。单独施加GR24时番茄叶片内POD活性与对照组无显著差异。
与对照相比,铅处理下番茄细胞内SOD活性显著降低42.60%;对幼苗进行GR24修复处理后发现SOD活性开始回升,与对照无显著差异,相比于铅处理提高了38.00%,这表明外源GR24可以缓解铅胁迫下引起的SOD活性降低现象。无铅处理下对幼苗施加GR24时SOD活性与对照无显著差异。与对照相比,铅胁迫处理番茄幼苗CAT活性显著下降了37.90%。添加外源GR24后番茄幼苗CAT活性显著升高,比对照显著提高43.56%,表明番茄幼苗在铅胁迫下施加外源GR24能够通过提高CAT活性来缓解铅诱发的氧化胁迫。单独施加外源GR24对CAT活性影响不大,与对照无显著差异。
2.2.4 外源GR24对铅胁迫下番茄幼苗MDA和ROS含量的影响 由图3-A可知,与对照相比,铅胁迫下番茄幼苗的MDA含量显著提高30.80%,推测铅胁迫会使番茄幼苗生物膜脂过氧化程度加深,生物膜系统遭到破坏。经过GR24修复后MDA含量下降,与对照无显著差异。单独施加GR24对幼苗MDA含量无显著影响。
由图3-B所示,用3种不同的染色液分别对番茄叶片进行染色时,对照叶片的颜色较淡或仅有几个蓝色小斑点,可以说明CK叶片的细胞膜结构比较完整,H2O2含量很低,[O2[-·]]生成能力也比较弱。单独铅胁迫处理的叶片,用Evans blue、DAB和NBT进行染色后,叶片颜色均变深,斑点数量也有所增加。在重金属铅胁迫下施加外源GR24,其染色叶片颜色明显变浅,斑点减少,但缓解程度不同。而与对照相比,外源GR24单独处理的植株叶片颜色与对照更为相似,表明外源GR24明显减轻了番茄植株的铅胁迫损害。
2.2.5 外源GR24对铅胁迫下番茄根系活力的影响 由图 4-A可知,相比于对照组,铅胁迫下番茄幼苗的主根长与侧根数量均显著降低,分别下降25.58%、27.27%。铅胁迫下同时添加GR24,主根长较Pb处理显著提高了21.06%,此时的侧根数量仍持续减少,达到最小值。单独添加GR24时,番茄幼苗的主根长达到最大值,比对照高出16.50%,而侧根数较对照降低了22.73%,均与对照存在显著差异。
从图4-B可以看出,番茄植株施用GR24处理的根系活力最高,达13.53 μg·g-1·h-1,显著高于对照。而在Pb胁迫下,番茄的根系活力最低,与对照相比,降低了42.14%。对铅胁迫下的番茄幼苗施加GR24根系活力较Pb处理有显著上升趋势,升高了24.35%。
2.2.6 外源GR24对铅胁迫下番茄幼苗生物量的影响 从表3可以看出,在铅胁迫下,番茄幼苗的地上部鲜、干质量和地下部鲜、干质量与对照相比均有显著下降,分别下降了24.63%、25.70%、34.12%、和44.89%。在铅胁迫的同时添加GR24,各部分生物量较Pb处理均有提高。所有处理中,单独施加外源GR24处理的生物量最高,其地上部鲜质量、地上部干质量和地下部鲜质量分别比对照显著提升了27.99%,36.64%和11.32%。
3 讨论与结论
3.1 外源GR24对铅胁迫下番茄幼苗光合色素的影响
植物体内叶绿素与类胡萝卜素等光合色素含量与植株的衰老、营养胁迫以及光合作用强弱都密切相关,是评估植物营养、生长、干物质含量、新陈代谢和酶活性的重要依据[23]。本研究结果表明,铅能抑制光合色素的产生,降低番茄叶片中的总叶绿素含量,提高类胡萝卜素水平。施加外源GR24能有效促进总叶绿素和类胡萝卜素合成,显著减轻铅胁迫对某些叶绿体结构的破坏,增强植物的抗逆性,更有利于光合产物形成和积累,缓解了铅对植物光合作用的抑制效应,这与前人的研究结果一致[13,24]。
3.2 外源GR24对铅胁迫下番茄幼苗渗透调节物质含量的影响
可溶性糖和可溶性蛋白是重要的渗透调节因子,可以调节植物细胞渗透势来抵御逆境伤害,重金属铅的过量积累同样会使植物营养物质的摄取和分配发生紊乱,进而削弱植物体内的氧化过程以及脂肪代谢过程强度[25]。在本试验中与对照相比,番茄幼苗在铅胁迫下可溶性糖含量下降,SP含量显著下降,表明铅胁迫影响植物生理机能,抑制可溶性糖和SP积累,加快分解,致使植物体内可溶性糖和SP含量降低,这与前人的研究结果一致[26-27]。GR24作用后可溶性糖含量显著升高,甚至高于对照,表明GR24能够促进番茄幼苗可溶性糖的积累,增强细胞膜的渗透调节能力,从而起到抗逆性调节作用。游离脯氨酸是植物中最重要的渗透调节物质之一,用于抗氧化、清除自由基、稳定细胞膜结构和保护细胞质中的各种酶,正常情况下,植物体内脯氨酸的水平非常低,当遇到逆境胁迫时,植物会产生大量的脯氨酸,以降低自身所受伤害,从而维持机体的正常生理代谢功能[28]。铅胁迫下番茄幼苗中的脯氨酸含量大幅上升,说明在不利条件下番茄可通过调节体内的渗透物质来保护细胞膜结构,这与在小麦中的研究结果一致[29]。对铅胁迫后的番茄添加GR24,叶片中的脯氨酸含量较单一铅胁迫处理显著下降,说明GR24可以降低铅胁迫下番茄幼苗脯氨酸含量,减轻铅胁迫对植物细胞膜的损伤,减少脯氨酸在植物体内的积累量,进而维持细胞的渗透平衡,从而提高番茄抵御铅胁迫伤害的能力。以上结果表明,铅胁迫下施加外源GR24会增加番茄植株SP含量,进而增强番茄植株的渗透调节作用及膜透性,提高细胞保水能力,维持细胞渗透平衡。
3.3 外源GR24对铅胁迫下番茄幼苗SOD、POD、CAT活性的影响
重金属胁迫能使植物体内产生大量活性极高并且伴有毒性的活性氧自由基,严重损害植物体内蛋白质、脂类、核苷酸等物质。植物体内的保护酶系统可有效清除因环境胁迫而产生的过量的活性氧,其中SOD、POD、CAT是普遍存在的重要的抗氧化酶,可以清除过量活性氧自由基[30]。在本试验中番茄幼苗由于铅胁迫导致SOD、POD、CAT活性均显著下降,施加外源GR24后这3种酶活性均有提高,表明铅处理下添加外源GR24可不同程度地增强番茄幼苗中SOD、POD、CAT活性,清除番茄幼苗组织在铅胁迫下产生的过量活性氧自由基,提高番茄幼苗在铅胁迫环境的生存能力,这与张灿[13]研究独角金内酯能缓解大麦镉毒害结果一致。因此,在受铅胁迫毒害的番茄幼苗中,外源GR24能够促进抵抗活性氧自由基物质的再生,增强幼苗的抗氧化酶活性,缓解铅胁迫对番茄幼苗生长发育的抑制。
3.4 外源GR24對铅胁迫下番茄幼苗MDA和叶片ROS含量的影响
MDA是植物膜脂过氧化的终产物,可以作为植物抗逆境胁迫耐受性检测的指标,当植物受到胁迫时,大量活性氧自由基积累在植物体内致使植物膜脂过氧化程度加深,MDA含量增加[31]。MDA含量与胁迫程度呈正相关,其含量越高代表膜损伤程度越高。植物自身在应对逆境胁迫时会本能地进行生理调节。在笔者的试验中,MDA含量在重金属铅的处理下升高,施加GR24之后,MDA含量开始下降,表明在一定程度上GR24能够使植物叶片中的MDA含量降低。这与王乔健[12]在乌桕上的研究结果一致:GR24能够使MDA含量减少,加速乌柏幼苗清除细胞膜脂过氧化终产物,减轻其对细胞膜的伤害,增强乌柏在逆境胁迫环境中的生存能力。
在正常环境下,活性氧(ROS)在植物中的代谢水平处于平衡状态。然而,在遭受不利环境胁迫下,植物体内会产生和积累大量的[O2[-·]],从而对植物造成伤害[32]。铅胁迫会造成醌类化合物在植物中积累,从而产生大量活性氧物质(ROS),如[O2[-·]]和H2O2。[O2[-·]]具有很强的氧化作用,可损害植物细胞;H2O2是细胞膜发生膜脂过氧化的重要因素之一[33]。在不同处理条件下,H2O2的积累量通过DAB染液染色后,叶片颜色由浅褐色至深褐色的变化情况来评估;NBT染液染色后,叶片上蓝色斑点的数目可以反映出叶片细胞产生[O2[-·]]活性的强弱;而Evans blue染色后,叶片上的颜色由淡蓝色至深蓝色的变化,可用来评价叶片细胞受损伤的程度,斑点的数目愈多,或者颜色愈蓝,则说明植物叶片在逆境中所遭受的损伤越重[19]。在本研究中,单一铅胁迫下,染色使番茄幼苗叶片颜色加深,斑点数量增加,表明重金属铅胁迫破坏了番茄叶片的细胞膜结构,并诱导了大量H2O2和[O2[-·]]产生。施加外源GR24后,其染色叶片颜色明显变浅,斑点数量较单一铅胁迫明显减少,这与董亚茹等[19]在桑树中的研究结果一致,进一步说明外源GR24可通过减少叶片中H2O2和[O2[-·]]的积累量,缓解番茄幼苗在铅胁迫下受到的伤害,提高植株的耐铅性。
3.5 外源GR24对铅胁迫下番茄主根长度、侧根数量及根系活力的影响
根是植物的主要营养器官,很容易受到外界环境因素的影响,对毒性响应的敏感度极高,因此,根系的形态直接反映出其对环境的适应性[20]。在本试验4个处理中,铅胁迫下的番茄幼苗主根长度最短,侧根数也低于对照,表明铅胁迫对番茄主根的伸长和侧根的形成均有明显的抑制作用。施加外源GR24后,主根长度增加,但侧根数继续减少,尤其是单独添加GR24时,主根长度达到最大值,表明GR24能有效促进番茄主根的伸长,并抑制侧根的形成,这一点与庞娟[20]在黄芪中的研究结果相似,这也进一步证实了GR24可以参与调控番茄根系的生长发育,调整作物根部构型和营养吸收状况,提高作物对水和养分的吸收效率。
根是植物的主要营养器官,可以从土壤中吸取水分和养分、合成氨基酸和内源激素等,根的生长与发育情况直接影响着植物对水分和营养的吸
收[34]。植物根系的呼吸速率越高,其活性越强,因此根系中TTC还原量就越多;反之,随着根系呼吸能力减弱,TTC还原量也相应地降低[35]。笔者试验发现,在铅的作用下,番茄幼苗的根系活力明显下降,铅对番茄根系的正常生理机能产生了损害,在一定程度上影响了根系的生长发育,进而降低了根系活力。施加GR24后,较铅胁迫下根系活力整体上升,表明GR24可有效缓解铅胁迫对番茄根系的生长抑制,并能对植株的正常生长起到一定的调节作用,将根系活力维持在一定水平。
3.6 外源GR24对铅胁迫下番茄幼苗生长的影响
过量的铅能使植物根系受损,减缓植物地上部分的生长。有研究表明,女贞苗体内积累过量的Pb会抑制植株生长,严重时甚至导致死亡[36]。另有研究表明,与对照相比,0.05 mmol·L-1 Pb处理对矮牵牛组培苗的根长、茎长、茎鲜质量以及根干质量均无显著影响,但随着Pb浓度的升高,矮牵牛组培苗的株高、根长以及生物量均显著下降[37]。在笔者的试验中,高浓度铅胁迫能够降低番茄幼苗的株高、根长、单叶面积和真叶数,铅胁迫环境下,番茄幼苗的地上部鲜、干质量和地下部鲜、干质量相比对照也有显著下降趋势,在铅胁迫的同时添加GR24,各部分生物量较Pb处理均表现出上升趋势。所有处理组中,单独施加外源GR24处理的生物量最高,其地上部鲜质量、地上部干质量和地下部鲜质量分别比对照显著提升。
本试验结果表明,高于200 mg·L-1的铅会抑制番茄幼苗的生长发育,铅胁迫下施用外源GR24能够显著提高番茄叶片光合色素、SP、可溶性糖含量;还可以通过增强SOD、POD、CAT等抗氧化酶活性来降低番茄植株体内活性氧水平,同时减少叶片MDA含量,有效缓解铅胁迫造成的氧化损伤,减缓植物衰老进程;降低铅胁迫下番茄幼苗的脯氨酸含量,抑制叶片中H2O2和[O2[-·]] 的产生,促进植株生物量的积累,促进主根的伸长,抑制侧根的生成,提升根部的根系活力。可见,外源独脚金内酯能有效缓解铅胁迫对番茄幼苗的损害,緩解铅的毒害效应。由于独角金内酯的作用机制比较复杂,其对番茄的具体影响机制有待研究,在实际生产中的应用需进一步探讨。
参考文献
[1] 韦朝阳,陈同斌.重金属超富集植物及植物修复技术研究进展[J].生态学报,2001,21(7):1196-1203.
[2] SHAHID M,ARSHAD M,KAEMMERER M,et al.Long-term field metal extraction by pelargonium:Phytoextraction efficiency in relation to plant maturity[J].International Joumal of Phytoremediation,2012,14(5):493-505.
[3] ESTRELLA-GOMEZ N,MENDOZA-COZATL D,MORENO-SANCHEZ R,et al.The Pb-hyperaccumulator aquatic fern Salviria minima Baker,responds to Pb2+ by increasing phytochelatins via changes in SmPCs expression and in phytochelatin synthase activity[J].Aquatic Toxicology,2009,91(4):320-328.
[4] 田学军,刘成兴,卢焕仙.铅胁迫对油菜生理的影响[J].西南农业学报,2010,23(2):605-607.
[5] 段代祥,刘俊华.重金属铅胁迫对绿豆种子萌发及幼苗生长的抑制效应[J].种子,2021,40(1):84-87.
[6] 李琬婷,黄轩,程小毛,等.铅胁迫对中华常春藤生长和生理特性的影响[J].云南农业大学学报(自然科学),2019,34(1):103-109.
[7] 郑晨.新型植物生长调节剂独脚金内酯生物学功能及应用[J].湖北农业科学,2020,59(2):9-13.
[8] JIANG L,LIU X,XIONG G S,et al.DWARF 53 acts as a repressor of strigolactone signalling in rice[J].Nature,2013,504(7480):401-405.
[9]马敏艳.独脚金内酯(±)-GR24及其类似物的合成研究[D].西安:陕西师范大学,2016.
[10] 万林,李张开,李素,等.外源独脚金内酯对油菜苗期干旱胁迫的缓解效应[J].中国油料作物学报,2020,42(3):461-471.
[11]胡超,万林,张利艳,等.独脚金内酯缓解油菜渍水胁迫的生理机制[J].中国油料作物学报,2017,39(4):467-475.
[12] 王乔健.独脚金内酯调控乌桕抗旱耐盐的分子机理研究[D].合肥:安徽农业大学,2019.
[13] 张灿.外源独脚金内酯缓解大麦镉毒害的机理研究[D].杭州:浙江大学,2019.
[14] 郭世荣.无土栽培学[M].2版.北京:中国农业出版社,2011.
[15] 李西进.铅胁迫对番茄种子萌发和幼苗生长的影响[J].长江蔬菜,2009(12):21-22.
[16] 王学奎.植物生理生化实验原理和技术[M].2版.北京:高等教育出版社,2006.
[17] 刘家尧,刘新.植物生理学实验教程[M].北京:高等教育出版社,2010.
[18] 刘新,刘洪庆.植物生理学实验[M].北京:高等教育出版社,2018.
[19] 董亚茹,张艳波,赵东晓,等.外源24-表油菜素内酯对NaCl胁迫下桑树幼苗的缓解效应[J].核农学报,2021,35(6):1466-1475.
[20] 庞娟.生长素与独脚金内酯调控黄芪幼苗根系生长发育的机制研究[D].呼和浩特:内蒙古大学,2020.
[21] 刘新,刘洪庆.植物生理学实验[M].北京:高等教育出版社,2018.
[22] 张天莹.两种典型抗生素对小麦幼苗生态毒性效应的影响[D].扬州:扬州大学,2020.
[23] 罗静.毛竹林不同施肥方式对叶片光合特性的影响[D].杭州:浙江农林大学,2015.
[24] 申璐.外源亚精胺对铅胁迫下茶树生长的影响[D].陕西杨凌:西北农林科技大学,2014.
[25] 赵钢,邹亮,彭镰心,等.铅胁迫对苦荞生理特性的影响[J].江苏农业科学,2012,40(7):98-100.
[26] 岳永成.烏苏里风毛菊对铅胁迫的生理抗性研究[D].兰州:西北师范大学,2014.
[27] 王芳,陈晓燕,曹廷俊,等.外源一氧化氮对铅胁迫下玉米幼苗的缓解作用[J].干旱地区农业研究,2014(4):24-29.
[28] 马英超.铅胁迫对构树幼苗光合和生理生化性质的影响[D].南昌:江西农业大学,2016.
[29] 钮力亚,王伟,王伟伟,等.盐胁迫下小麦品种生理指标的变化规律[J].中国农学通报,2019,35(2):1-4.
[30] 徐学华.北方地区7种主要绿化树种对铅镉胁迫的生理生态响应[D].保定:河北农业大学,2010.
[31] 何若韫.植物低温逆境生理[M].北京:中国农业出版社,1995.
[32] 秦建桥,赵华荣,赵鹏.镉胁迫下不同生态型五节芒的膜脂过氧化水平及抗氧化能力比较[J].安徽农业科学,2011,39(12):7418-7421.
[33] 陈晟,施木田,吴宇芬,等.应用组织染色法研究硼胁迫对西瓜叶片的生理伤害[J].中国农学通报,2016,32(7):64-68.
[34] 纠松涛,徐岩,张才喜,等.独脚金内酯及其调控植物根系生长发育的研究进展[J].分子植物育种,2021,19(15):5164-5171.
[35] 朱秀云,梁梦,马玉.根系活力的测定(TTC法)实验综述报告[J].广东化工,2020,47(6):211-212.
[36] ZHOU J,ZHANG Z P,ZHANG Y C,et al.Effects of lead stress on the growth,physiology,and cellular structure of privet seedlings[J].PLoS One,2018,13(3):e0191139.
[37] 杨颖丽,施树倩,李嘉敏,等.铅胁迫对矮牵牛组培苗生长及光合特性的影响[J].兰州大学学报(自然科学版),2023,59(1):47-62.
猜你喜欢
现代农业科技(2017年1期)2017-03-06 01:26:03
中西医结合心血管病电子杂志(2016年22期)2017-03-03 00:42:57
江苏农业科学(2017年1期)2017-02-27 11:11:38
南方农业·下旬(2016年8期)2017-02-23 19:53:16
吉林农业·下半月(2017年1期)2017-02-16 11:31:56
江苏农业科学(2016年8期)2017-02-15 20:29:25
江苏农业科学(2016年8期)2017-02-15 20:14:21
安徽农学通报(2017年1期)2017-02-15 18:23:48
物联网技术(2016年12期)2017-01-21 22:27:52
中国科技博览(2016年1期)2016-04-25 07:47:28
