
轻基质配比对春兰生长和生理生化特性影响的综合评价
2024-05-22 18:05:21刘红马辉魏晓羽孙叶
江苏农业科学 2024年7期
刘红 马辉 魏晓羽 孙叶


(江苏里下河地区农业科学研究所,江苏扬州 225007)
摘要:为解决传统春兰栽培基质保水、保肥性能差及成本高、生产运输不方便等影响春兰产业化生产的问题,探讨将松鳞与泥炭土作为春兰栽培基质利用的可行性。将松鳞与泥炭土按质量比1 ∶0(D1处理)、4 ∶1(D2处理)、3 ∶1(D3处理)、2 ∶1(D4处理)、1 ∶1(D5处理)、1 ∶2(D6处理)、0 ∶1(D7处理)配制成7种轻基质配比,并以传统硬基质配方植金石-仙土(质量比3 ∶1)作为对照(CK),分析轻基质对春兰植株生长指标、光合指标、生理指标和酶活性的影响,并用主成分分析法对各基质配比效果进行评价。结果表明,D2~D5处理的理化性质均属于理想基质范畴,能够保证春兰组培苗的正常生长。D4~D5处理与CK浇水频率以6 d左右为宜,D6~D7处理的浇水频率以8 d左右为宜,D1~D3处理则需要较高的浇水频率。D1、D4、D5、D6处理的新芽数、新芽率均显著高于对照,其中最高的为D4处理,新芽数为3个/盆,新芽率为100%。净光合速率最高的为D4处理,最低的为D6、D7处理,均与对照有显著差异。基质配比影响春兰植株养分的累积和抗氧化活性。不同配比基质的理化性质与春兰的壮苗指标、生理指标及光合指标密切相关。以测定的春兰生长、生理生化指标进行主成分分析,各基质配比的优劣排序为D4处理>D5处理>D3处理>D1处理>D7处理>D6处理>CK>D2处理。综合来看,用D4处理条件栽培春兰具有植株长势好、增殖率高的丰产特性,相比硬基质,其新芽率、根干重、壮苗指数、叶绿素含量、净光合速率、可溶性糖含量、SOD活性分别提高650.19%、49.30%、51.44%、13.93%、47.89%、57.81%、18.16%;同时,该基质成本降低了86.5%,质量降低了60.4%。建议选用松鳞与泥炭土质量比为2 ∶1的基质配比,用于低成本基质的开发及春兰的高品质栽培。研究结果不仅可以降低春兰规模化生产运输的成本,还可以为我国春兰的高品质栽培提供技术指导。
关键词:春兰;松鳞;泥炭土;轻基质;配比;生长特性;生理生化特性;主成分分析;综合评价
中图分类号:S682.310.4 文献标志码:A 文章编号:1002-1302(2024)07-0133-10
春兰(Cymbidium goeringii)是兰科(Orchidaceae)兰属(Cymbidium)的地生兰之一,主要分布于中国、日本与朝鲜半岛南端等地。中国春兰资源丰富,广泛分布于江苏、浙江、江西等16个省份,春兰的自然花期在我国传统节日春节前后,在我国已有数千年栽培鉴赏的历史。春兰的栽培品种一般分为梅瓣型、荷瓣型、水仙瓣型、奇瓣型、素心型、色花型和叶艺型等几个园艺类型[1],其花姿秀美、花香清幽,有非常典型的民族文化特色,是我国特有的珍稀兰花种质资源[2],具有极高的观赏价值和经济价值。栽培基质可以固定植物根系,且基质的物理、化学特性会直接影响基质的蓄水能力、透水性及通气性,对春兰生长有着至关重要的作用[3]。目前,春兰种植户使用的栽培基质大多基于传统经验,以进口硬植料植金石为主,辅以塘基石、仙土、珍珠岩、陶粒和煤渣等硬质颗粒基质,疏水透气性较好,但保水、保肥性能差,同时价格昂贵、质量大,生产运输不方便,无法满足春兰规模化生产和产业化开发的需求[4-5]。经过消毒、腐熟等处理后,很多农林废弃物都能成为优良的育苗基质,具有质轻、持水、保肥、通气性能好等优点,并且成本低,易获取,产品运输携带方便,但在春兰生产栽培上的应用未见报道,而洋兰生产广泛采用树皮、花生壳、椰块、木屑等农林废弃物作为栽培基质,取得了良好的栽培效果[6-8]。目前,蝴蝶兰(Phalaenopsis aphrodite Rchb. f.)、大花蕙兰(Cymbidium faberi×[JP]hybridum)、春石斛兰(Dendrboium)等洋兰栽培技术日益完善,已经实现了規模化的高效生产,代表设施农业发展水平较高[9]。鉴于洋兰产业化开发的经验,轻基质栽培是春兰产业发展的必然趋势,然而目前关于春兰轻基质栽培技术尚没有系统的研究。因此,本研究以腐熟的松树皮、泥炭土为原料,按照不同质量比配制春兰栽培基质,通过比较基质理化性质、植株生长、生理生化等指标,筛选适宜春兰栽培的轻基质配方,以期提高春兰栽培质量、降低春兰规模化生产运输的成本,为春兰的产业化开发提供理论依据。
1 材料与方法
1.1 试验材料
本试验选取生长健壮、无病虫害的2年生春兰大团圆大苗为材料,盆具为直径15 cm、高25 cm的兰花专用黑色硬质塑料盆。
1.2 研究方法
1.2.1 试验设计 根据笔者所在课题组前期的预试验结果,本研究所用基质主要由腐熟松鳞(0.5~1.5 cm)、泥炭土(丹麦品氏,10~30 mm)按质量比组成,试验设置7个配方处理(D1~D7处理),以传统硬基质配方为对照(CK),详见表1。每个处理设20盆,每盆3株春兰,3次重复,共160盆480株大苗。
试验于2021年10月在江苏里下河地区农业科学研究所中国兰种质资源圃温室内进行。培养过程中每隔7~10 d用稀释2 000倍的花宝2号灌根1次,15 d喷1次杀菌剂,多种杀菌剂交替使用,其间进行春兰常见病虫害的防治,同时及时清理病株。
1.3 测定指标与方法
1.3.1 基质物理性质的测定 (1)基质容重、总孔隙度、通气孔隙度、持水孔隙度和气水比的测定。采用环刀法,在朱丽等提供的方法[10]基础上进行优化。取体积为155 cm3的环刀,称重(m1);基质混匀后,随机取自然风干的混合基质,加满环刀,称重(m2);然后将装有基质的环刀用纱布封口,浸泡 24 h(水位线始终超过环刀顶部), 将塑料量杯从水中取出,在饱和水状态下称重(m3),封口纱布不滴水时称重(m4);最后再用湿纱布包住环刀,倒置 8 h,使量杯内水分(重力水)自由沥干,称重(m5)。每个处理测量9份样品。相关物理指标的计算公式如下:
容重(g/cm3)=(m2-m1)/155;(1)
持水能力=(m5-m1-m4)/(m2-m1) ×100%;(2)
总孔隙度=(m3-m2)/155×100%;(3)
通气孔隙度=(m3+m4-m5)/155×100%;(4)
持水孔隙度=总孔隙度-通气孔隙度;(5)
气水比=通气孔隙度/持水孔隙度。(6)
(2)基质pH值、EC值的测定采用饱和浸提法。将自然风干的基质与去离子水按体积比1 ∶5混合,充分搅拌30 min后用保鲜膜封口,静置10 h,取过滤水,用pH计和电导仪进行测定[10],每个处理测量9份样品。
1.3.2 浇水后基质、植株根和叶片含水量的测定 采用喷淋的方式进行同时浇水,浇透即止。在浇水后0、2、4、6、8、10 d,用精讯通畅土壤参数速测平台测定每盆基质的相对含水量,计算各处理20盆样品的平均含水量;取出春兰植株,将根、叶分离,分别称取根、叶的鲜重,然后将鲜样于105 ℃杀青 60 min,再于80 ℃烘至恒重,测定根、叶的干重,并计算根、叶含水量。每个处理随机取9株大苗进行测定,取其平均值。
1.3.3 生长指标的测定 试验6个月后,测量大苗株高、叶长和假鳞茎直径,并记录新芽数,计算存活率。选取1株/盆大苗进行测定,每个处理共计20株,取平均值。取出大苗,将根系的基质清洗干净,用滤纸吸干表面水分后测定根、假鳞茎和叶鲜重及其烘干后的干重(g)[11],计算根、假鳞茎和叶片含水量、根冠比及壮苗指数。每个处理设3个重复,每个重复设3株,共计9株,取平均值。
株高为植株自然状态下最高点距叶片基处的垂直高度;叶长为将叶片拉直后的长度。
新芽率=新芽数/苗数×100%;
存活率=存活苗数/总苗数×100%;
根冠比=根干重/地上部干重;
壮苗指数=(假鳞茎直径/株高+根干重/地上部干重)×全株干重。
1.3.4 生理生化指标的测定
1.3.4.1 光合指标 (1)处理60 d 后,采用托普云农植物营养测定仪(型号:TYS-4N D08)测定叶片叶绿素相对含量(以SPAD值计)。各选1张/盆叶片(叶龄2年)进行测定,每个处理共计测定20张叶片,取其平均值。
(2)在晴朗天气的09:00—11:30用TARGAS-1光合测定仪(汉莎科学仪器有限公司,美国)测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间 CO2 浓度(Ci)。选1张/盆叶片(叶龄2年)进行测定,每个处理共计测定20张叶片,取其平均值。
1.3.4.2 叶绿素、可溶性固形物、抗氧化酶等生理生化指标 试验处理结束后,以叶片为样品,进行生理生化指标的测定[11-12],每个处理随机取3个以上的材料进行混合,再从混合样中称量3份以上。可溶性糖含量用蒽酮比色法测定,可溶性蛋白含量用考马斯亮蓝法(Bradford法)测定,叶绿素含量用分光光度法测定,超氧化物歧化酶(SOD)活性用氮蓝四唑(NBT)法測定,过氧化物酶(POD)活性用愈创木酚法测定,过氧化氢酶(CAT)活性用分光光度法测定。
1.4 数据处理
用Excel 2016进行数据整理及作图,用SPSS 22.0软件对数据进行方差分析,用Duncan's法对数据的显著性进行多重比较,并进行主成分分析。
2 结果与分析
2.1 不同基质配比的理化性质
由表2可知,不同基质处理的理化性质间存在显著差异。D1~D7处理的容重在0.20~0.25 g/cm3之间,均显著低于CK(P<0.05),且与泥炭添加比例成正比。D1~D7处理的总孔隙度在58.64%~69.65%之间,通气孔隙度在12.14%~35.17%之间,持水孔隙度在34.48%~46.49%之间,均高于对照,且除D7处理的通气孔隙度外,均与CK间差异显著(P<0.05);其中,总孔隙度、通气孔隙度与泥炭添加比例成反比,最高的均为D1处理,分别为69.65%、35.17%,而持水孔隙度与泥炭添加比例成正比,以D7处理最高,为46.49%。D1~D7处理的气水比在0.26~1.02之间,与泥炭添加比例成反比,其中D1~D5处理的气水比高于对照,且D1~D3处理的气水比与CK差异显著(P<0.05)。随泥炭添加比例增加,D1~D7处理组的pH值呈下降趋势,但与CK差异均不显著;EC值呈上升趋势,最高的为D7处理,达1.55 mS/cm,除D1处理外,其他处理的EC值均高于CK,其中D5~D7处理的EC值与CK差异显著(P<0.05)。
有研究发现,理想基质的容重为0.1~0.8 g/cm3,总孔隙度为54%~96%[13],通气孔隙度为15%~30%[14],气水比为1 ∶(1.5~4.0)(即 0.25~0.67),H值为6.0~7.5,电导率<2.6 mS/cm。在本研究中,D1处理的通气孔隙度、气水比高于理想值,CK的总孔隙度为40.17%,低于理想值,D6、D7处理和CK的通气孔隙度低于理想值,D2~D5处理的理化性质均属于理想基质范畴,能保证春兰组培苗正常生长。
2.2 基质及根、叶片水分变化情况
由图1-A可知,浇水后,D7处理春兰的吸水性最强,相对含水量达46%左右,而其他处理的相对含水量均在30%以下,其中D1处理的吸水性最差,相对含水量最低,在10%以下。泥炭的保水性最强,随着松鳞添加比例的提高,其保水性逐渐下降,纯松鳞的保水性最差。D6、D7处理的春兰在浇水后8 d的相对含水量下降到15%左右,而D4、D5春兰的相对含水量在浇水后6 d下降到该水平,D1~D3处理春兰的含水量始终低于15%。图1-B可见,不同处理春兰根系的含水量在浇水后持续下降,D1~D3处理及CK春兰植株根的含水量到浇水后 6 d 时下降到90%以下,而D4~D7处理春兰植株根系含水量在浇水后8 d下降到90%左右。图1-C显示,春兰叶片含水量最低的为D1处理,其含水量下降得最快,而其他处理春兰叶片含水量在浇水后4 d才开始快速下降,浇水后4 d时,D2~D3处理及CK比D4~D7处理的叶片含水量低2%左右。结合植株状态,D4~D5与CK处理的浇水频率以6 d左右为宜,D6~D7处理的浇水频率以8 d左右为宜,D1~D3处理则需较高的浇水频率。
2.3 不同基质配比对春兰生长的影响
2.3.1 不同基质配比对春兰形态指标的影响 由表3可知,不同基质配比对春兰株高、叶长、新芽数和新芽率有显著影响。株高最高的为D3处理,为16.90 cm;最低的为D7处理,仅为13.40 cm。叶长最长的是D3处理,其次为D4处理,均高于对照,而D1、D2、D5、D7处理的叶长低于对照,各处理春兰叶长与对照差异不显著。各处理假鳞茎直径与对照差异不显著,其中最高的为D4处理(11.27 mm),最低的为D7处理(10.20 mm)。D1、D4、D5、D6处理的新芽数、新芽率均显著高于对照(P<0.05),其中最高的为D4处理,新芽数为3个/盆,新芽率为100%。
2.3.2 不同基质配比对春兰壮苗指标的影响 由表4可知,根干重最大的为D4处理,为1.06 g,最小的为D7处理,为0.53 g,且均与对照差异显著(P<0.05)。叶干重最大的为D6处理,为1.19 g,最小的为D3处理,为0.39 g,二者相差2.05倍,且均与对照差异显著(P<0.05)。各处理假鳞茎干重均低于对照(0.47 g),其中以D2处理最低,仅为0.27 g。根冠比最高的为D3处理,达2.18,最低的为D6处理,仅为0.47,且均与对照差异显著(P<0.05)。根含水量最高的为D7处理,最低的为D5处理,二者间差异显著,但均与对照无显著差异。叶含水量最高的为D2处理,最低的为D3处理,二者间差异显著(P<0.05)。假鳞茎含水量最高的为D5处理,最低的为D2处理,二者间差异显著(P<0.05)。壮苗指数最高的为D4处理,为3.68,最低的为D7处理,为1.90,且二者均与对照差异显著。
2.4 不同基质配比对春兰光合指标的影响
由表5可知,不同基质处理下春兰的光合指标有显著差异。SPAD值最高的为D4处理,其次为D2处理,均显著高于对照(P<0.05)。净光合速率最高的为D4处理,最低的为D6、D7处理,且均与对照差异显著(P<0.05)。蒸腾速率最高的为D5处理,最低的为D1处理。各处理气孔导度和胞间CO2浓度总体高于对照,气孔导度最高的为D5处理,胞间CO2浓度最高的为D4处理,且均显著高于对照(P<0.05)。
2.5 不同基质配比对春兰养分积累的影响
表6显示,各处理之间春兰叶片叶绿素含量差异显著,其中以D4处理最高,为34.18 pmol/g,D3处理最低,为19.59 pmol/g。在不同基质处理下,春兰叶片的可溶性糖含量均高于对照,其中D3处理最高,为436.46 μg/g,D4处理的可溶性糖含量略低于D3处理,为429.28 μg/g,而D7处理相对最低,为291.43 μg/g。可溶性蛋白含量最高的为D1处理,最低的为D7处理,均与对照差异显著(P<0.05)。上述结果表明,基质配比会影响春兰植株养分的累积。
2.6 不同基质配比对春兰抗氧化酶活性的影响
由表7可知,SOD活性最高的为D5处理,为729.86 U/g,其次为D4处理,为648.73 U/g,且不同处理之间存在显著差异(P<0.05)。CAT活性最高的为D4处理,为92.98 U/mL,而其余处理均显著低于对照(P<0.05)。POD活性最高的为D6处理,为4.19 mU/g,最低的为D1处理,为2.13 mU/g,且均与对照差异显著(P<0.05)。
2.7 基质理化性质与春兰生长的相关性
基质的理化性质与春兰生长、壮苗、生理及光合指标之间的相关性分析结果(表8)表明,基质的容重与春兰的CAT活性呈极显著正相关,与假鳞茎干重、根含水量、假鳞茎含水量及可溶性糖含量呈极显著负相关,与蒸腾速率呈显著负相关;总孔隙度与假鳞茎干重、假鳞茎含水量、可溶性糖含量和蒸腾速率呈极显著正相关,与根含水量呈显著正相关,与CAT活性呈极显著负相关;通气孔隙度与假鳞茎干重、根含水量呈显著正相关,与可溶性糖含量呈极显著正相关,与叶绿素含量、过氧化氢酶活性呈极显著负相关;持水孔隙度与叶重、假鳞茎干重、根含水量、假鳞茎含水量、可溶性糖含量、超氧化物歧化酶活性、净光合速率、蒸腾速率、气孔导度和胞间CO2浓度呈极显著正相关,与新芽率呈显著正相关,与叶含水量呈显著负相关;气水比与假鳞茎干重、SOD活性、净光合速率、蒸腾速率、气孔导度、胞间CO2浓度呈极显著正相关,与可溶性糖含量呈显著正相关,与根冠比呈显著负相关,与叶含水量呈极显著负相关;pH值与假鳞茎干重、可溶性糖含量呈极显著正相关,与净光合速率呈显著正相关,与根冠比呈显著负相关;EC值与叶干重、假鳞茎干重呈显著负相关,与根冠比呈极显著正相关,与可溶性糖含量、可溶性蛋白含量、SOD活性和净光合速率呈极显著负相关。由此可见,基质各理化性质与春兰的壮苗指标、生理指标及光合指标密切相关。
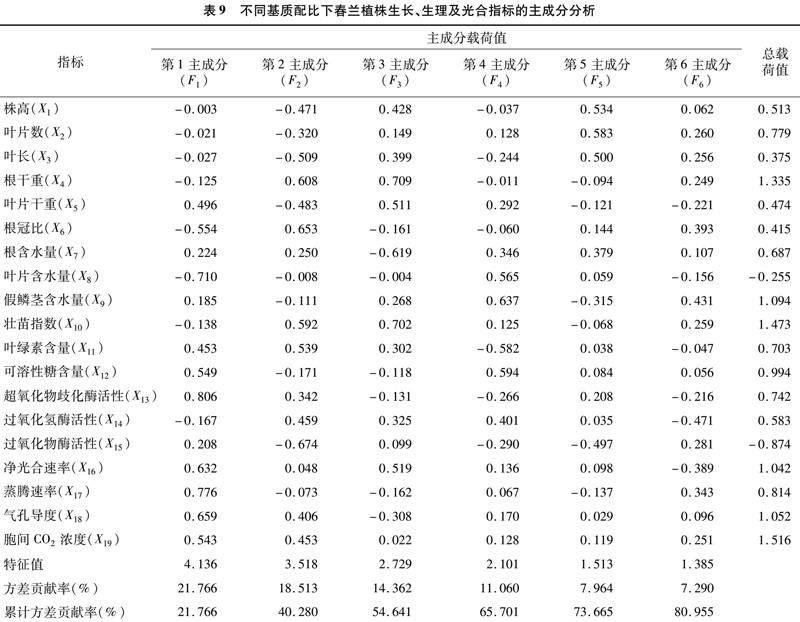
2.8 对春兰各指标主成分分析及综合评价
本研究通过对春兰的19个生长指标、壮苗指标、生理指标及光合指标进行主成分分析,获得6个主成分,其累计贡献率達80.996%,可以保留原始因子中代表的绝大部分信息(表9)。用Yi表示各主成分,得出各主成分线性方程:
Y1=0.396X1+0.110X2+0.244X3+0.091X4+0.027X5+0.223X6-0.010X7-0.001X8-0.082X9+0.381X10-0.349X11+0.324X12+0.311X13+0.267X14+0.102X15-0.272X16-0.013X17-0.062X18-0.068X19;
Y2=0.182X1+0.133X2-0.258X3-0.059X4-0.091X5+0.288X6-0.171X7-0.251X8+0.245X9-0.039X10-0.004X11+0.217X12+0.026X13+0.241X14-0.359X15+0.348X16-0.271X17+0.324X18+0.316X19;
Y3=-0.079X1-0.374X2+0.309X3+0.162X4-0.071X5+0.183X6+0.090X7+0.259X8+0.197X9-0.098X10-0.002X11-0.186X12+0.314X13+0.013X14+0.060X15-0.098X16+0.241X17+0.429X18+0.425X19;
Y4=-0.184X1+0.239X2+0.201X3+0.439X4+0.410X5-0.402X6+0.088X7-0.026X8+0.277X9+0.046X10+0.390X11+0.11712+0.093X13+0.088X14-0.200X15-0.348X16-0.041X17-0.008X18+0.086X19;
Y5=0.169X1+0.308X2-0.098X3-0.256X4+0.068X5+0.031X6+0.474X7+0.434X8+0.029X9-0.111X10+0.048X11+0.023X12+0.079X13+0.097X14-0.404X15+0.117X16+0.406X17-0.077X18-0.055X19;
Y6=-0.183X1+0.091X2-0.188X3-0.366X4+0.048X5-0.040X6+0.221X7+0.052X8-0.400X9+0.292X10-0.132X11+0.082X12-0.331X13+0.213X14+0.239X15+0.334X16+0.218X17+0.212X18+0.220X19。
由表9可知,第1主成分的方差贡献率为21766%,载荷值最大的为SOD活性,第1主成分综合了SOD活性、蒸腾速率、叶片含水量、气孔导度、 净光合速率和胞间CO2浓度等变量信息, 主要
反映春兰植株的抗氧化酶活性和光合作用情况;第2主成分的方差贡献率为18.513%,载荷值最大的为POD活性;第3主成分中,根干重、壮苗指数、根含水量和叶片干重等变量主要反映春兰植株的壮苗情况,方差贡献率为14.362%;第4主成分中,假鳞茎含水量、可溶性糖含量和叶绿素含量主要反映春兰植株的生理指标,方差贡献率为11.060%;第5主成分中,叶片数和株高反映春兰植株的生长指标,方差贡献率为7.964%;第6主成分为CAT活性,方差贡献率为7.290%。
将6个主成分得分用隶属函数归一化处理,根据各主成分的贡献率计算权重系数,得到春兰栽培基质的进行综合得分值:Y=0.217 7Y1+0.185 1Y2+0.143 6Y3+0.110 6Y4+0.079 6Y5+0.072 9Y6。
由表10可以看出,8种基质的综合得分值的变化范围为-0.422~0.848,其中最高的为D4处理,其次为D5处理,最低为D2处理。由此可见,最适合春兰植株的基质为D4处理(松鳞、泥炭的体积比=2 ∶1),最不适宜的基质为D2处理(松鳞、泥炭的体积比4 ∶1)。
3 讨论
3.1 轻基质栽培春兰的可行性及理化性状
栽培基质是春兰生长关键因子之一,基质的肥力、透水性、透气性对春兰的生长起着重要作用[5]。优良的栽培基质应具备稳定良好的理化性质。本研究发现,基质容重会影响春兰植株的假鳞茎干重、根含水量、假鳞茎含水量、可溶性糖含量、蒸腾速率及CAT活性。容重反映的是基质的疏松、紧实程度,是栽培基质最重要的筛选指标之一。基质容重越大, 其疏松程度越差,在同样体积下,容重越大,基质质量越大。本试验中各处理基质的容重均在适宜范围(0.1~0.8 g/cm3)之内,其中CK的容重最大,达0.65 g/cm3,而D1~D7处理的容重仅为CK的30.77%~38.46%,可见轻基质配方较传统硬基质配方质量下降得较多,有利于降低运输成本、节约劳动力。基质的水分特性分为吸水性和保水性,吸水性是指基质一定时间能吸入的水分的量,是提高浇水效率和制定浇水策略的依据之一[15]。基质的孔隙度是影响水分特性的主要因素。在本研究中,总孔隙度及通气孔隙度对春兰植株的假鳞茎干重、假鳞茎含水量、根含水量、叶含水量、可溶性糖含量和光合作用指标等影响较大,在合理范围内,二者数值越大,越有利于植物根系的透水和呼吸,对植物生长越有利。D1处理的容重最小,从通气孔隙度与持水孔隙度来看,其通气性和保水性相近,气水比高于理想孔隙度范围,浇水后基质含水量低于10%,表现为基质贮水保肥能力弱,植株根系和叶片水分下降速度快,植株生长发育受阻。随着泥炭添加比例的增加,基质吸水性和保水性逐渐增强,保水性均大于通气性,贮水保肥能力提高,适宜植株生长发育。CK总孔隙度和通气孔隙度低于各轻基质配方,表示基质的疏松度及透水性相对较差,以其栽培的春兰壮苗指数也相对较低。基质的pH值影响植物生长和基质养分的形态与有效含量[16],在本研究中,各处理基质的pH值均在适宜范围之内,表明基质养分的有效性都较高,植株对养分的吸收能力也较强,对植物生长有利。EC值是指基质中可溶性盐的含量,虽然自然状态下各基质的EC值处于正常水平,但D7处理的EC值达 1.55 mS/cm,为其他处理的3.44~9.12倍,说明纯泥炭土含有较高浓度的可溶性盐,在实施生产中,由于施肥后会增加EC值,因此需要定期對基质EC值进行检测,通过调配基质或调整施肥方案,使 EC 值保持在正常水平。
3.2 不同配比基质对春兰生长的影响
分析春兰的生长情况得知,D3~D5处理的春兰生长性能、壮苗指标均较好,植株生长健壮。植株的株高、假鳞茎直径及干鲜重、壮苗指数是最能直接反映植物生长状态的指标,其中壮苗指数作为一项综合指标,避免了单一指标在秧苗质量评定中的片面性,可以较为全面地衡量植株是否健壮,壮苗指数越高,越有利于加强光合作用[17-18]。植物根系是吸收养分和水分的主要器官,根系的生长发育直接影响植物个体的生长发育[19]。孙燕等推测,基质通过影响植株根系吸收能力及光合能力影响植株生长[20]。用D3~D5处理基质栽培的春兰生长指标优于其他处理及对照,可能因为这些基质理化性质适于春兰根系生长,从而促进植株生长发育。而对于不同栽培基质对春兰植株根系活力的影响仍有待进一步研究。
3.3 不同基质对春兰生理生化指标的影响
叶绿素含量是反映植物营养状态的重要指标,叶绿素含量越高,越有助于植物的光合作用[21]。在一定范围内,叶绿素含量与净光合速率呈正相关[20],其变化趋势直接关系着植株生物量的积累速率[22]。在本研究中,叶绿素相对含量(SPAD值)、叶绿素含量、净光合速率与根干重、壮苗指数最高的均为D4处理(松鳞-泥炭土配比为2 ∶1),说明该基质提高了春兰的光合作用能力,即养分同化吸收能力较强,与上述研究结果较为一致。而CK的叶绿素相对含量、气孔导度最低,其原因可能是硬基质孔隙度最低,使其根系水气交换受到限制,从而影响叶片的光合作用活力。
植物生存所需的最低光照(即耐阴性),是一个至关重要的生活史特征[23]。野生春兰的生境多为湿度较高、空气清新、无烈日和高温的针阔混交林或其他林地,使其生长缓慢。有学者对5种国兰(建兰、春兰、春剑、蕙兰和墨兰)的光合特性进行研究,发现春兰光补偿点低,利用弱光的能力比较强,属于喜阴植物[24-25]。光合作用直接反映作物的生长状况,宋军阳研究发现,秦岭野生春兰的净光合速率在0.33~1.69 μmol/(m2·s)之间[24]。在本研究中,春兰的净光合速率在 0.42~1.05 μmol/(m2·s) 之间,与前人的研究结果相近。叶庆生等对兰属植物的7个种中的10个栽培品种光合途径进行研究发现,这些兰属植物的净光合速率较低,其中春兰的净光合速率为3.33 μmol/(m2·s)[26],而大多数植株的净光合速率在5~50 μmol/(m2·s)[JP]之间,说明春兰的光合作用能力较弱,是其生长缓慢的重要原因。
可溶性糖是植物光合作用的积累产物,是植物主要的能量来源物质,其含量高表明光合作用正常,也是评价容器苗的关键指标[27]。在本研究中,叶片可溶性糖含量较高的为D4处理,同时表现为叶绿素含量、净光合速率高,表明具有较强的光合作用能力。SOD、CAT和POD是植物体内常见的抗氧化酶,当植物受到病害或者是逆境胁迫时,这些酶会被激活或被大量合成,从而起到保护植物的作用[28]。D2、D4、D5处理的SOD活性显著高于对照,D4处理的CAT活性最高,D2、D3、D6处理的POD活性高于对照,表明用这些处理的基质栽培有利于提高春兰的抗逆性。
基质筛选大多根据研究者观测经验确定适宜配方,增加人为主观随意性带来的偏差。本研究采用主成分分析法,利用以上数学模型对8种基质配方的春兰指标进行综合评价,计算综合主成分得分,以各配方的综合得分值排序,能更客观地反映不同基质配方的优劣。由本研究结果可知,D4处理的综合得分最高,为0.848分,而D2处理的综合得分最低,为-0.422分。按综合得分对8种基质配方进行排序,其优劣顺序为D4处理>D5处理>D3处理>D1处理>D7处理>D6处理>CK>D2处理。上述结果表明,D3、D4、D5处理基质适宜春兰栽培,其中最佳基质配方为D4处理。
4 结论
综上可知,D3~D5处理几种基质的理化性质均属于理想范畴,均能保证春兰的正常生长,且其栽培春兰生长指标优于对照。其中,D4处理的假鳞茎直径、新芽数和新芽率、根干重、壮苗指数、叶绿素含量、净光合速率及CAT活性均最高。此外,通过主成分分析得出,D4处理排名第一,为春兰栽培最适的培养基。
综合本研究结果,建议选用D4处理的配方(松鳞 ∶泥炭土=2 ∶1),该基质既能提高春兰生长势,又能改善植株品质,提高光合作用及抗氧化酶活性水平;同时,该配方基质成本约1.55元/盆,质量为377 g/盆,而同等条件下,硬植料成本为11.52元/盆,质量为952.22 g/盆,使用轻基质栽培,基质成本降低了865%,基质质量降低了60.4%,这不但提高了种苗质量与栽培效率,还降低了生产成本,使生产运输更方便。
参考文献:
[1]Wang H Z,Wu Z X,Lu J J,et al. Molecular diversity and relationships among Cymbidium goeringii cultivars based on inter-simple sequence repeat (ISSR) markers[J]. Genetica,2009,136(3):391-399.
[2]陳心启,吉占和. 中国兰花全书[M]. 北京:中国林业出版社,1998.
[3]刘 娟. 喀斯特不同生态修复措施下植被与土壤特征研究[D]. 贵阳:贵州大学,2017:49-64.
[4]严 华,张冬梅,陆 颖. 不同栽培基质和粪肥对下山春兰生长的影响[J]. 福建林业科技,2012,39(4):82-85.
[5]李媛媛,邹 军,范志伟,等. 不同栽培基质在春兰上的应用效果[J]. 江苏农业科学,2014,42(6):199-201.
[6]李 琛. 不同栽培基质对大花蕙兰生长发育的影响[D]. 长沙:湖南农业大学,2012:13-31.
[7]许明修,朱 娇,马 蕾,等. 不同栽培基质对蝴蝶兰矿质营养积累的影响[J]. 热带作物学报,2016,37(7):1261-1265.
[8]孙 燕,杨秀珍,钱 璐,等. 花生壳栽培基质对大花蕙兰‘世界和平生长及生理特性的影响 [J]. 浙江农林大学学报,2015,32(4):566-571.
[9]朱根发,杨凤玺,吕复兵,等. 兰花育种及产业化技术研究进展[J]. 广东农业科学,2020,47(11):218-225.
[10]朱 丽,陈晓东,乔玉山,等. 不同配比基质对草莓母苗生长和抽生匍匐茎的影响[J]. 江西农业大学学报,2021,43(3):547-554.
[11]赵永平,朱 亚,闫敏飞,等. 不同供钾水平对金盏花幼苗生长特性和酶活性的影响[J]. 江西农业学报,2017,29(1):39-42.
[12]欒 添,杨秀珍,李 惠,等. 钾营养水平对大花蕙兰小苗生长及养分吸收的影响[J]. 中国农业大学学报,2014,19(4):103-107.
[13]王旭艳,林夏珍,李 琳,等. 几种农林废弃物复合基质的理化特性及对浙江楠容器育苗的效果 [J]. 浙江农林大学学报,2013,30(5):674-680.
[14]马赞留,蔡红海,戴云新,等. 不同配比基质对番茄幼苗生长的影响[J]. 安徽农学通报,2018,24(15):86-97.
[15]车艳丽,李彦明,杨其长,等. 酒糟沼渣在番茄基质育苗上的应用[J]. 中国农业大学学报,2021,26(1):88-98.
[16]钱又宇,薛 隽. 世界著名观赏树木 美国流苏树·美国香槐[J]. 园林,2009(7):66-77.
[17]文莲莲,李 岩,张聃丘,等. 冬季温室补光时长对番茄幼苗生长、光合特性及碳代谢的影响[J]. 植物生理学报,2018,54(9):1490-1498.
[18]刘小金,徐大平,杨曾奖,等. 脱落酸对檀香幼苗生长、光合及叶片抗氧化酶活性的影响[J]. 南京林业大学学报(自然科学版),2016,40(3):57-62.
[19]丁小涛,姜玉萍,刘 娜,等. 不同栽培基质对夏季温室番茄生长和光合特性的影响[J]. 上海农业学报,2019,35(6):46-53.
[20]孙 燕,赵青云,龙宇宙,等. 不同育苗基质下咖啡种间嫁接苗生长及光合特性[J]. 热带作物学报,2021,42(6):1606-1612.
[21]曹 钰,胡 涛,张鸽香. 基质配比对美国流苏容器苗生长的影响[J]. 东北林业大学学报,2018,46(9):26-30.[HJ2.1mm]
[22]Wu W M,Chen H J,Li J C,et al. Effects of nitrogen fertilization on chlorophyll fluorescence parameters of flag leaf and grain filling in winter wheat suffered waterlogging at booting stage[J]. Acta Agronomica Sinica,2013,38(6):1088-1096.
[23]Fernando V,lo N. Shade tolerance,a key plant feature of complex nature and consequences[J]. Annual Review of Ecology Evolution & Systematics,2008,39(1):237-257.
[24]宋军阳. 秦岭野生春兰光合特性研究[J]. 林业科技通讯,2019(12):59-62.
[25]李鹏民,高辉远,邹 琦,等. 5种国兰(Cymbidium)的光合特性[J]. 园艺学报,2005,32(1):151-154.
[26]叶庆生,潘瑞炽,丘才新. 兰属植物光合途径的研究[J]. 热带亚热带植物学报,1998,6(1):25-29.
[27]Dan D,Long T,Zhang G Y,et al. Quantitative trait loci analysis for chlorophyll content of cucumber (Cucumis sativus L.) seedlings under low-light stress[J]. African Journal of Biotechnology,2010,9(30):4692-4699.
[28]孙跃跃,马军妮,李玉龙,等. 功能性育苗基质中生防菌及腐植酸钾对甜瓜穴盘苗的促生作用 [J]. 西北农林科技大学学报(自然科学版),2016,44(9):174-180.
基金项目:江苏现代农业产业技术体系建设专项基金(编号:JATS[2023]242);江苏省省级农业科技创新与推广补助专项(编号:扬农[2023]73号);江苏省农业种质资源保护与利用平台项目(编号:JSGB2018-1);江苏里下河地区农业科学研究所科研发展专项资金-揭榜挂帅项目(编号:SJ[22]105)。
作者简介:刘 红(1986—),女,黑龙江伊春人,硕士,助理研究员,主要从事花卉遗传育种与产业化生产关键技术研究。E-mail:liuhong_ivy@126.com。
通信作者:孙 叶,硕士,副研究员,主要从事花卉遗传育种研究。E-mail:sunye9999@126.com。
猜你喜欢
快乐作文(1.2年级)(2023年2期)2023-03-30 01:09:26
健康体检与管理(2022年4期)2022-05-13 18:42:17
老年博览·上半月(2021年12期)2021-12-21 02:30:39
森林工程(2018年4期)2018-08-04 03:23:16
时代农机(2018年11期)2018-03-17 06:05:31
西南农业学报(2016年6期)2016-04-16 05:12:58
广西林业科学(2016年4期)2016-03-16 05:45:04
电源技术(2016年9期)2016-02-27 09:05:28
小小说月刊·下半月(2015年2期)2015-05-14 15:23:17
中成药(2014年9期)2014-02-28 22:28:55
