
烟草镰刀菌根腐病病原生物学特性及其优势种的代谢表型特征
2024-05-22 18:05:21黄玉凤李菲汪汉成蔡刘体陈兴江邱学柏
江苏农业科学 2024年7期
关键词:生物学特性
黄玉凤 李菲 汪汉成 蔡刘体 陈兴江 邱学柏


黄玉凤,李 菲,汪汉成,等. 烟草镰刀菌根腐病病原生物学特性及其优势种的代谢表型特征[J]. 江苏农业科学,2024,52(7):124-132.
doi:10.15889/j.issn.1002-1302.2024.07.017
(1.贵州师范大学生命科学学院/西南喀斯特山地生物多样性保护国家林业和草原局重点实验室,贵州贵阳 550001;2. 贵州省烟草科学研究院,贵州贵阳 550081)
摘要:为明确烟草镰刀菌根腐病(tobacco Fusarium root rot)病原生物學特性和代谢表型特征,采用菌丝生长速率法和活体接种法分别测定了3种烟草镰刀菌根腐病病原[腐皮镰刀菌(Fusarium solani)、木贼镰刀菌(Fusarium solani)、尖孢镰刀菌(Fusarium oxysporum)]在不同温度、光照条件下的生长速率和致病力,同时采用Biolog代谢表型技术测定其优势种腐皮镰刀菌的碳氮源、渗透压和pH值下的代谢表型。结果表明,3种病原菌菌丝生长适宜温度均为25~30 ℃,木贼镰刀菌M417菌丝生长速率最快,可达10.71 mm/d;腐皮镰刀菌F421产孢量最高,可达9.15×10.6 个/cm2;连续黑暗条件均有利于菌丝生长和产孢。尖孢镰刀菌J419和腐皮镰刀菌F503致病力较强,7 d时的病斑面积分别为1.05 cm2和1.35 cm2。代谢表型结果表明,腐皮镰刀菌碳氮源代谢能力强,可高效代谢丙三醇、D-甘露醇、D-松二糖等54种碳源,以及L-鸟氨酸a-氨基-N-戊酸、L-鸟氨酸等大部分氮源;其pH值适应范围为3.5~8.5,最适pH值为6,表现出强脱羧酶活性和弱脱氨酶活性;在1%~5.5% NaCl、5%~10%乙二醇、1%甲酸钠、2%~3%尿素、7%~9%乳酸钠、10~50 mmol/L硫酸铵(pH值=8)、10~100 mmol/L硝酸钠、10~40 mmol/L亚硝酸钠等环境下均可正常生长。研究结果揭示了3种镰刀菌属根腐病病原的生物学特性,发现优势种腐皮镰刀菌的营养需求广、具有较强的渗透压和pH值环境适应能力,为镰刀菌根腐病灾变规律的研究提供了参考。
关键词:烟草根腐病;镰刀菌属;生物学特性;代谢表型
中图分类号:S435.72 文献标志码:A 文章编号:1002-1302(2024)07-0124-09
烟草是我国重要的经济作物,被广泛种植并用于生产香烟、雪茄等烟草制品[1]。然而,烟草种植过程中常常受到多种病害的威胁,其中一种真菌性土传病害——镰刀菌根腐病(tobacco Fusarium root rot)较为常见[2]。该病害由镰刀菌属(Fusarium sp.)真菌侵染引起,被报道的致病菌有尖孢镰刀菌(Fusarium oxysporum)、木贼镰刀菌(Fusarium equiseti)、腐皮镰刀菌(Fusarium solani)等[3-4]。该病主要发生于烟草大田期,典型症状有烟株矮小、底部叶片叶尖黄化蜷缩焦干。近年来,随着气候条件与种植方式的变化,该病害在我国贵州烟区的发生范围及危害程度呈现上升、加重趋势,给烟农造成严重经济损失[5],亟须对其病原的生物学特性加强研究,以期掌握该病害的灾变规律。
温度、光照等环境因子影响着植物病原菌的生长[6]。已有少量烟草根腐病病原菌生物学特性的报道,赵杰研究表明,烟草根腐病病原尖孢镰刀菌和腐皮镰刀菌在10~35 ℃温度范围内均可生长,最适生长温度为30 ℃[7]。陈高航对烟草尖孢镰刀菌研究发现,其致死温度为60 ℃,且环境适应性强[8]。汪汉成等研究发现,烟草腐烂病病原菌九州镰刀菌(F. kyushuense)在完全黑暗、完全光照、光暗交替环境下菌丝生长速率无显著性差异[9]。此外,碳氮源营养、渗透压等环境因子也影响着病原菌的生物活性,病原菌对不同营养因子的代谢表型特征可反映其对环境的适应力。文增叶等研究发现,三七根腐病病原尖孢镰刀菌可有效利用淀粉、乳糖等多种碳源和硝酸铵等氮源[10]。杨静美等研究发现,番木瓜腐皮镰刀菌(Fusarium solani f. sp. glycine)在pH值 3~10范围内均可生长,最适pH值为7~10[11]。相较而言,作为严重危害烟叶生产的根腐病,其病原菌的生物学特性和环境适应力却缺乏了解。
为此,本研究采用菌丝生长速率法分别测定镰刀菌根腐病病原在不同温度、光照条件下的生长速率,利用活体接种法测定不同种的致病力,同时采用Biolog代谢表型技术研究其在不同碳氮源、渗透压和pH值下的代谢表型,旨在明确其生物学特性和环境适应力,为烟草根腐病的灾变规律研究提供参考依据。
1 材料与方法
1.1 试验概况及材料
试验于 2023年6—11月在贵州省贵阳市观山湖区贵州省烟草科学研究院进行。烟草镰刀菌根腐病病原腐皮镰刀菌(F. solani)菌株F421、F506、F503,木贼镰刀菌(F. equiseti)菌株M417、M423、M16,尖孢镰刀菌(F. oxysporum)菌株J419、J3、J210,均由贵州省烟草科学研究院真菌实验室提供。供试烟草品种为云烟87,待烟叶长至7~8叶时备用。
培养基:马铃薯葡萄糖琼脂(PDA)培养基:马铃薯200 g、葡萄糖20 g、琼脂15 g、蒸馏水1 L;燕麦琼脂(OA)培养基:燕麦片30 g、琼脂17 g、蒸馏水 1 L;烷基酯琼脂培养基(AEA)培养基:酵母提取粉5 g、琼脂20 g、丙三醇20 mL、硝酸钠6 g、磷酸氢二钾1.5 g、氯化钾0.5 g、硫酸镁0.25 g;绿豆汤培养基:新鲜绿豆20 g,洗净,放入1 000 mL纯净水中煮沸20 min,过滤去除豆渣,滤液加水补足1 000 mL,加入琼脂粉20 g,搅拌均匀,分装,121 ℃湿热灭菌20 min。
Biolog代谢板FF(货号:94545)、PM3(货号:12121)、PM9(货号:12161)、PM10(货号:12162)和接种液FF-IF(filamentous fungi-inoculating fluid)(货号:72106),均由美国Biolog公司生产;D-葡萄糖、磷酸二氢钾、硫酸钠等,购自美国Sigma公司;酵母氮源,购自美国Difco公司;Omnilog PM高通量微生物细胞表型芯片测定系统,由美国Biolog公司生产。
1.2 方法
1.2.1 烟草镰刀菌根腐病病原的生物学特性 (1)温度对菌丝生长和产孢量的影响:用菌丝生长速率法测定腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌在不同温度范围的菌丝生长速率。用直径6 mm打孔器制取培养7 d的根腐病菌菌碟,置于PDA培养基上,于5、10、15、20、25、30、35 ℃黑暗条件下培养,每处理设3次重复,接种7 d后采用“十字交叉法”测量菌落直径,并计算生长速率。平板测定完后,用10 mL无菌水洗涤平板,制成孢子悬浮液,在显微镜下利用血球计数板测定单位面积(cm2)的产孢量。(2)光照条件对菌丝生长和产孢量的影响:采用菌丝生长速率法测定不同光照条件下菌丝生长速率。使用无菌打孔器在PDA培养基上制取直径为6 mm的培养7 d的根腐病菌菌碟,将其接种到新的PDA培养基中,置于温度均为28 ℃、光照条件分别为连续光照、连续黑暗、光—暗周期为12 h—12 h的生化培养箱,每个处理设置3次重复,接种 7 d 后采用“十字交叉法”测量菌落直径,并计算生长速率。随后用10 mL无菌水洗涤平板,制成孢子悬浮液,在显微镜下利用血球计数板测定单位面积(cm2)的产孢量。(3)菌丝致死温度的测定:随机选取3株菌株(尖孢镰刀菌J419、腐皮镰刀菌F503、木贼镰刀菌M16)进行菌丝致死温度测定。在PDA培养基上制取直径为6 mm的菌碟,将其置于盛有 5 mL 灭菌水的无菌试管中,分别置于35、40、45、50、55 ℃恒温水浴锅中水浴10 min,同时缓慢摇动试管使之受热均匀,取出试管冷却至室温,将处理后的菌饼接种到PDA平板中央,并置于28 ℃培养箱中黑暗培养,6 d后观察菌丝生长情况以确定致死温度范围。每个处理重复3次。(4)不同培养基对菌丝生长的影响:参照上述方法制备腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌直径6 mm 的菌碟,分别置于PDA、OA、AEA、绿豆汤培养基上,接菌后所有平板均置于温度为28 ℃黑暗条件下的培养箱中培养。每处理重复3次,7 d后测量菌落直径。
1.2.2 烟草镰刀菌根腐病病原的致病力测定 随机选取尖孢镰刀菌菌株J419、腐皮镰刀菌菌株F503、木贼镰刀菌菌株M16进行病原菌致病力测定。烟草茎部和根部的致病力测定参照毕武等的方法[12],叶部致病力的测定参照汪汉成等的方法[8]。将6 mm菌饼分别接种到刺伤和无刺伤的烟苗组织上,接种后菌碟和烟株组织采用浸有无菌水的脱脂棉保湿,随后放在28 ℃黑暗条件下的人工气候箱中培养,对照组接种无菌丝的PDA菌碟。接种7 d后,采用“十字交叉法”测量病斑直径,每个菌株重复3次。
1.2.3 代谢表型测定 由“1.2.1”节测定结果可知腐皮镰刀菌F421产孢能力较强,选择此菌株进行测定。使用Biolog系统测定其代谢表型。将烟草根腐病菌接种到PDA培养基上,于28 ℃、[JP2]黑暗条件下培养7 d,至产生大量分生孢子。无菌棉签蘸用FF接种液(FF-IF)浸湿,在菌落表面旋转,接着将棉签浸入20 mL的 FF-IF[JP]接种液中制备1×10.5 个/mL的孢子悬浮液。碳源代谢表型测定:将孢子悬浮液依次加入FF代谢板微孔中(100 μL/孔);氮源代谢表型测定:取10.025 mL孢子悬浮液、D-葡萄糖溶液0.375 mL、酵母氮源溶液1 mL与无菌水0.6 mL均匀混合加入到PM3代谢板中;渗透压和pH值环境下代谢表型测定:取孢子悬浮液20.05 mL、D-葡萄糖溶液0.75 mL、 酵母氮源溶液2 mL、无菌水 1.2 mL 均匀混合,加入PM9和PM10代谢板中;按照Biolog PM微生物细胞表型流程将各测定板放于Biolog 培养箱28 ℃培养,设置 OmniLog 工作软件,每15 min收集1次数据。根据动力学曲线,分析其代谢表型特征。
1.2.4 数据分析 使用Excel 2010和SPSS 24.0软件对试验数据进行单因素方差分析,应用LSD(least significant difference)法进行差异显著性检验。
2 结果与分析
2.1 烟草镰刀菌根腐病病原的生物学特性[HT]
2.1.1 温度对菌丝生长速率的影响 在不同温度下,腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌的菌丝生长速率各不相同,且存在显著性差异。在5~30 ℃ 条件下,所测尖孢镰刀菌、腐皮镰刀菌和木贼镰刀菌菌株均能生长。随着温度的升高,菌丝生长速率均呈现先增加后减小的趋势。在25~30 ℃时,9株菌株的菌丝生长速度均较快。在25 ℃时,木贼镰刀菌M16、木贼镰刀菌M423、腐皮镰刀菌F421、尖孢镰刀菌J210的菌丝生长速率较快,分别为7.07、10.45、8.43、10.00 mm/d;在30 ℃时,木贼镰刀菌M417、腐皮镰刀菌F506、腐皮镰刀菌F503、尖孢镰刀菌J419、尖孢镰刀菌J3的菌丝生长速率最快,分别达到10.71、9.78、8.90、8.64、6.67 mm/d。25 ℃和30 ℃的生长速度显著高于其他温度处理(P<0.05)(图1-A)。9株菌株菌丝生长的适宜温度为25~30 ℃,低于15 ℃或高于30 ℃菌丝生长缓慢。
2.1.2 温度对产孢量的影响 在不同温度下,烟草镰刀菌属根腐病菌产孢量之间存在差异。随着温度升高,腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌的产孢量呈现先增加后减小的趋势。在15~35 ℃温度范围内均可产孢,在25~30 ℃ 时产孢量最多。在25 ℃时,木贼镰刀菌M423、尖孢镰刀菌J419、尖孢镰刀菌J210产孢量最多,分别为7.45×10.6、5.33×10.6、4.95×10.6 个/cm2;在30 ℃时,腐皮镰刀菌F421、腐皮镰刀菌F506产孢量最多,分别为 8.65×106、7.40×10.6個/cm2;而腐皮镰刀菌F503、尖孢镰刀菌J3、木贼镰刀菌M16、木贼镰刀菌M417在这2个温度范围内的产孢量无显著性差异。在35 ℃,产孢量呈下降趋势,尖孢镰刀菌J3、尖孢镰刀菌J419、木贼镰刀菌M16、木贼镰刀菌M417、木贼镰刀菌M423在此温度范围不产孢(图1-B)。总之,9株镰刀菌产孢的适宜温度为25~30 ℃,低于 15 ℃ 或高于30 ℃时产孢量少甚至不产孢。
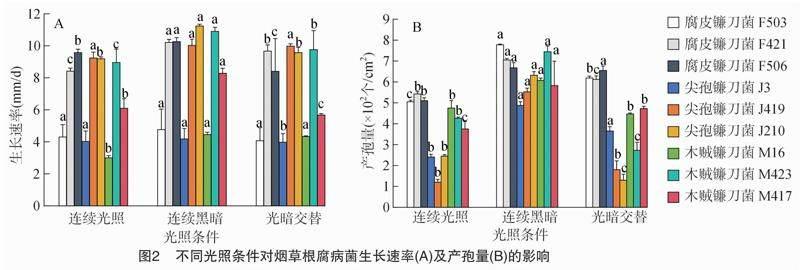
2.1.3 光照对菌丝生长的影响 腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌在连续光照、连续黑暗和 12 h—12 h光暗交替条件下,均可正常生长。在3种光照条件下,木贼镰刀菌M16在连续黑暗条件和12 h—12 h光暗交替条件下的菌丝生长速率分别为4.45、4.33 mm/d,两者之间无显著性差异,但显著高于连续光照条件;尖孢镰刀菌J3在连续黑暗和连续光照条件下的菌丝生长速率分别为4.18、4.02 mm/d,两者之间无显著性差异,但显著高于 12 h—12 h光暗处理条件;尖孢镰刀菌J210在连续光照条件和12 h—12 h光暗交替条件下的菌丝生长速率分别为9.19、9.57 mm/d,两者之间无显著性差异,但显著低于连续黑暗条件。在3种光照测试条件下,腐皮镰刀菌F503的菌丝生长速率范围为4.07~4.76 mm/d,尖孢镰刀菌J419的菌丝生长速率为9.24~10.02 mm/d,相同菌种的不同菌株间均无显著性差异;在黑暗条件下,木贼镰刀菌M417、木贼镰刀菌M423、腐皮镰刀菌F421、腐皮镰刀菌F506的菌丝生长速率均最快,显著高于其他2个光照条件处理(P<0.05,图2-A)。因此,连续黑暗条件为腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌菌丝生长的最适光照条件。
2.1.4 光照对产孢量的影响 在不同光照条件下,所有腐皮镰刀菌、 木贼镰刀菌和尖孢镰刀菌菌株均可产孢,且产孢量在不同光照条件下存在差异性。
由图2-B可知,9株镰刀菌属根腐病菌在黑暗条件下的产孢量最多,例如腐皮镰刀菌F421的产孢量为7.05×10.6个/cm2,尖孢镰刀菌J3的产孢量为 4.88×10.6 个/cm2。连续光照条件下,产孢量均最少。为此,连续黑暗条件有利于腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌9株镰刀菌的产孢。
2.1.5 菌丝的致死温度 腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌在35、40 ℃水浴处理10 min后,重新置于28 ℃黑暗條件下的人工气候箱中培养均可生长,而在45、50、55 ℃条件下水浴处理后不能生长(表1)。为此,3株镰刀菌的致死温度均为45 ℃。
2.1.6 不同培养基对镰刀菌属根腐病病原菌丝生长速率的影响 结果表明,在PDA培养基、OA培养
基、AEA培养基、绿豆汤培养基上,腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌的菌丝生长速率存在差异(表2)。在相同条件下,木贼镰刀菌M16、尖孢镰刀菌J210、腐皮镰刀菌F506在AEA培养基上的菌丝生长速率最大,分别为10.19、11.36、10.07 mm/d,OA培养基和绿豆汤培养基的菌丝生长速率差异不显著,而PDA培养基的菌丝生长速率最慢。尖孢镰刀菌J3和腐皮镰刀菌F503在AEA培养基上的菌丝生长速率最快,分别为9.72、9.95 mm/d,在PDA培养基和绿豆汤培养基的菌丝生长速率无显著性差异。木贼镰刀菌M417在AEA培养基上的菌丝生长速率最大,达到11.09 mm/d,在OA培养基和PDA培养基上的菌丝生长速率无显著性差异。木贼镰刀菌M423在PDA培养基、AEA培养基、绿豆汤培养基上的菌丝生长速率分别4.29、5.24、4.36 mm/d,三者之间无显著性差异。尖孢镰刀菌J419在AEA培养基上和绿豆汤培养基上的菌丝生长速率分别为9.36、8.88 mm/d,两者之间无显著性差异。腐皮镰刀菌F421在AEA培养基上的菌丝生长速率最大,达到10.21 mm/d,而在PDA培养基上的菌丝生长速率最小。为此,3种镰刀菌菌丝生长的最适培养基均为AEA培养基。
2.2 烟草镰刀菌根腐病病原的致病力
致病力试验结果表明,3株腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌菌株刺伤、无刺伤接种,烟草叶片7 d后均出现明显的发病症状,且刺伤接种的尖孢镰刀菌J419、腐皮镰刀菌F506的致病力较强,病斑面积分别为1.35 cm2和1.05 cm2;木贼镰刀菌M16的致病力较弱。3菌种在叶部均形成褐色病斑,病斑周围有黄褐色晕圈,3个种在叶片上的致病力强弱顺序依次为尖孢镰刀菌>腐皮镰刀菌>木贼镰刀菌。腐皮镰刀菌、木贼镰刀菌和尖孢镰刀菌在接种茎部后,茎部均出现白色菌丝体,随后病斑逐渐扩大直至整株死亡,3个种在茎上的致病力强弱顺序依次为尖孢镰刀菌>腐皮镰刀菌>木贼镰刀菌。根部接种3种病原菌后,烟株叶部均发现不同程度的萎蔫黄化,根系破碎,须根减少(图3)。
2.3 腐皮镰刀菌的代谢表型特征
2.3.1 碳源代谢表型 由图4可见,腐皮镰刀菌对Biolog FF板上的所有碳源都能代谢,不同碳源间的代谢强度存在差异。其中可高效代谢的碳源约有54种,包括D-阿拉伯糖醇、丙三醇、D-甘露醇、D-松二糖、L-焦谷氨酸等,但对α-D-乳糖、γ-羟基丁酸等5种碳源的代谢能力相对较弱。
2.3.2 氮源代谢表型 由图4可见,腐皮镰刀菌对Biolog PM3板上的所有氮源都能代谢。其中,能够高效代谢的氮源包括Met-Ala、α-氨基-N-戊酸、L-鸟氨酸等6种氮源;对8种氮源的代谢能力相对较弱,包括羟胺、甲胺、正戊胺、e-氨基-N-己酸、谷胱甘肽等;相较而言,腐皮镰刀菌对谷胱甘肽的利用能力最弱。
2.3.3 渗透压和pH值代谢表型特征 由图5可知,腐皮镰刀菌具有较强的渗透压适应能力,可代谢10~40 mmol/L 亚硝酸钠、1%~ 5.5% 硫酸钠、1%甲酸钠、1%和7%~9%的乳酸钠、5%~10% 乙二醇、2%~3%的尿素、10~100 mmol/L 硝酸钠、10~40 mmol/L 亚硝酸钠,而对6.5%、8%的氯化钠、20~200 mmol/L 苯甲酸钠(pH值5.2)、60~80 mmol/L 亚硝酸钠、7%的尿素的代谢较弱。在6% 氯化钠与不同渗透压物质协同下,腐皮镰刀可代谢渗透压物质,但对甜菜碱、四氢嘧啶、肌酸、γ-氨基-N-丁酸的代谢较弱。腐皮镰刀菌可进行代谢的pH值范围为3.5~8.5,最适pH值为6。当pH值 4.5胁迫与不同氨基酸混合时,腐皮镰刀菌可高效代谢大部分基质,例如L-谷氨酸、L-天冬氨酸、L-丝氨酸、L-丙氨酸、L-组氨酸、L-天门冬酰胺等,但不能代谢邻氨基苯甲酸。当pH值 9.5胁迫与不同氨基酸混合时,腐皮镰刀菌可代谢大部分基质,如L-谷氨酸、L-组氨酸、L-丙氨酸、腐胺、胍基丁胺、酪胺等,而对L-酪氨酸的代谢较弱。此外,还可高效代谢X-β-D-氨基葡萄糖苷、X-β-D-氨基半乳糖苷、X-氨基磷酸酶、X-芳基硫酸酯酶、X-SO4等基质。在pH值分别为4.5和95的不同氨基酸胁迫下,分别测定了B1~D12和E1~G12孔中PM10的脱羧酶和脱氨酶活性。结果反映该病原菌的脱羧酶活性较高,但脱氨酶活性较差。
3 讨论
烟草镰刀菌根腐病是发生在烟草上的真菌性土传病害,在我国各种植烟区均有发生,其危害于烟草根部,对烟草的经济效益造成重大损失[13-14]。温度和光照条件能显著影响病原菌的生长。本研究发现腐皮镰刀菌、木贼镰刀菌、尖孢镰刀菌在 10~35 ℃条件下均可生长,25~30 ℃温度范围内菌丝生长速率最大、产孢量最多,这与赵杰报道的结果[7]基本一致。本研究发现烟草镰刀菌根腐病病原生长的适温范围与同为烟草土传病害的烟草黑胫病菌Phytophthora parasitica(21~35 ℃)[15]类似,这可能是烟草田间病害镰刀菌根腐病与黑胫病常常混发、难以防治的原因之一。汪汉成等报道九州镰刀菌光照处理对菌丝生长没有明显影响,光暗交替更利于其产孢[16];而本研究发现尖孢镰刀菌、腐皮镰刀菌、木贼镰刀菌在黑暗条件下更利于菌丝生长和产孢,其差异可能是不同种镰刀菌的生物学特性不同造成的。本研究发现烟草镰刀菌属根腐病病原尖孢镰刀菌、腐皮镰刀菌、木贼镰刀菌均可致烟草叶部、茎部和根部发病,结果与姚健等研究认为的烟草根腐病病原菌尖孢镰刀菌和腐皮镰刀菌可致烟草发病的结论[17]一致,进一步验证了本研究结果的可靠性。王立国等研究报道,广金钱草根腐病病原腐皮镰刀菌在PDA和OA培养基上的菌丝生长差异不显著[18],而本研究发现腐皮镰刀菌菌丝在OA培养基上比PDA培养基上生长更快,说明同种菌菌株间在不同培养基的生长速度间存在差异。
营养元素是病原菌生长的必要条件,碳源和氮源的利用是其生长最重要的两大营养元素。病原菌对不同碳氮源利用的种类和强度存在差异性,如汪汉成等[19]研究报道烟草赤星病菌可高效代谢的碳源有L-果胶糖、D-半乳糖等34種,氮源有L-谷氨酸、L-赖氨酸等60余种。本研究采用Biolog FF碳源代谢板进行测试,FF板大部分碳源为真菌可代谢碳源,能反映腐皮镰刀菌对大部分碳源的需求。本研究发现腐皮镰刀菌可高效代谢大部分碳源,如D-阿拉伯糖醇、丙三醇等,推测这些碳源有助于根腐病菌的生长。通过Biolog PM3氮源代谢板可知腐皮镰刀菌对大部分氮源的代谢能力一般,且对谷胱甘肽的代谢能力最弱,推测这类氮源可抑制根腐病菌的生长。
渗透压和pH值等环境因子对病原菌的生长与危害具有重要影响[19],病原菌在不同环境中的代谢能力变化,可反映其对环境的耐受性。本研究基于Biolog表型技术测定了烟草镰刀菌属根腐病菌腐皮镰刀菌生长的最适pH值为6。当pH值为4.5时,腐皮镰刀菌可代谢大部分氨基酸;当pH值为9.5时,烟草根腐病菌腐皮镰刀菌对氨基酸代谢活性相对较弱;该结果反映腐皮镰刀菌的脱羧酶活性较高,但脱氨酶活性较差。烟草根腐病菌与烟草其他病原菌相比,其pH值适应范围与烟草青枯病菌Ralstonia solanacearum (pH值 5.0~10) [20]、烟草黑胫病菌 (pH值 3.5~10)[15]相似。病原菌在烟草上的pH值范围不同可能是其脱羧酶活性和脱氨酶活性的差异造成的。病原菌对不同渗透压物质的代谢活性不同,可反映其在不同逆境中的耐受力。烟草腐皮镰刀菌对20~200 mmol/L 苯甲酸钠(pH值 5.2)、60~80 mmol/L亚硝酸钠、甜菜碱、四氢嘧啶等部分物质的代谢较弱,因此可使用不同浓度的渗透压物质来减轻该病害的危害,这需进一步深入研究。本研究获得的烟草腐皮镰刀菌的代谢表型组学信息为镰刀菌属其他病原菌的代谢表型研究提供了借鉴。
参考文献:
[1]刘 茜,李义婷,蔡刘体,等. 烟草根茎部病原真菌的分离与鉴定[J]. 中国烟草科学,2022,43(6):45-52.
[2]刘利佳,李芳芳,何 雷,等. 烟草镰刀菌根腐病病原菌的鉴定及其对5种杀菌剂的敏感性分析[J].河南农业科学,2021,50(7):101-109.
[3]邱 睿,白静科,李成军,等. 河南烟草镰刀菌的分子鉴定及致病性分析[J]. 中国烟草学报,2018,24(2):129-134.
[4]邱 睿,李小杰,李 娟,等. 河南烟区烟草镰刀菌根腐病病原鉴定及侵染烟草根系观察[J]. 烟草科技,2023,56(7):17-24.
[5]盖晓彤,卢灿华,户艳霞,等. 云南省烟草镰刀菌根腐病病原鉴定[J]. 中国烟草学报,2023,29(3):74-83.
[6]胡吉凤,姜于兰. 温度和光照对烟草青枯病菌生长的影响[J]. 山地农业生物学报,2015,34(6):46-49.
[7]赵 杰. 山东省烟草镰刀菌根腐病病原及生物学特性的研究[D]. 北京:中国农业科学院,2013:18-19.
[8]陈高航. 烟草根腐病病原鉴定及其生物学特性观察[D]. 武汉:华中农业大学,2013:19-22.
[9]汪汉成,王进,李文红,等. 烤烟苗期茎部腐烂病病原菌九州镰孢菌的生物学特性[J]. 中国烟草学报,2014,20(1):65-70.
[10]文增叶,李定华,代梦瑶,等. 三七根腐病病原菌尖孢镰刀菌的生物学特性分析[J]. 中药材,2019,42(9):1978-1984.
[11]杨静美,陈 健,罗金棠,等. 番木瓜茄病镰刀菌的生物学特性研究[J]. 中国热带农业,2011,38(1):56-58.
[12]毕 武,陈 娟,焦晓琳,等. 北京地区西洋参根腐病病原菌鉴定及其致病性[J]. 植物保护,2011,37(5):135-138.
[13]孙会忠,王洁琼,郝浩浩,等. 两株异源生防菌的鉴定及其对烟草根腐病的协防增效[J]. 河南大学学报(自然科学版),2023,53(5):575-583.
[14]白茂军,高正锋,张力元,等. 烟草青枯病发病程度与土壤环境间的响应关系[J]. 江苏农业学报,2023,39(6):1294-1302.
[15]刘 畅,向立刚,汪汉成,等. 温度对烟草黑胫病菌致病力及代谢表型的影响[J]. 植物保护学报,2021,48(3):669-678.
[16][JP2]汪汉成,王 进,李文红,等. 烤烟苗期茎部腐烂病病原菌九州镰孢菌的生物学特性[J]. 中国烟草学报,2014,20(1):65-70.[JP]
[17]姚 健,刘玉珍,李建华,等. 许昌烟草根腐病的分子鉴定及致病性分析[J]. 江西农业学报,2020,32(3):99-103.
[18]王利国,王晓英,翁伟华,等. 广金钱草根腐病病原生物学特性研究[J]. 中华中医药学刊,2008,3 (1):147-149.
[19]汪汉成,黄艳飞,陈兴江,等. 烟草赤星病菌嘧菌酯敏感与抗性菌株的代谢表型差异分析[J]. 植物病理学报,2018,48(6):822-832.
[20]Chen X J,Li L C,Wang H C et al. Phenotypic fingerprintsp of Ralstoniaso lanacearum under arious osmolytes and pH environments [J]. Plant Pathology Journal,2016,15(3):102-107.
基金项目:国家自然科学基金(编号:32160522、31960550);贵州省科技基金(编号:黔科合基础-ZK[2021]重點036);中国烟草总公司科技项目[编号:110202101048(LS-08)、110202001035(LS-04)];贵州省“百层次”创新型人才项目(编号:黔科合平台人才-GCC[2022]028-1);贵州省科技创新人才团队项目“茄科叶用经济作物病害绿色防控科技创新人才团队建设”。
作者简介:黄玉凤(1998—),女,贵州遵义人,硕士研究生,主要从事病原微生物学研究工作。E-mail:397521401@qq.com。
通信作者:汪汉成,研究员,主要从事烟草植保及微生态研究,E-mail:xiaobaiyang126@hotmail.com;李 菲,教授,主要从事植物根际微生态研究,E-mail:ifei00987@outlook.com。
猜你喜欢
山东农业科学(2016年12期)2017-01-21 15:02:46
现代园艺(2016年7期)2017-01-09 22:53:28
现代农业科技(2016年19期)2016-12-12 16:15:50
农业与技术(2016年19期)2016-12-12 03:12:12
农业与技术(2016年20期)2016-12-08 19:26:26
现代园艺(2016年13期)2016-09-02 10:29:29
现代园艺(2016年13期)2016-09-02 10:28:33
现代园艺(2016年4期)2016-03-24 11:56:07
