
斑石鲷irf3 基因鉴定及其在虹彩病毒感染下的表达模式
2024-05-20 05:22:50李开敏巩志宏王清滨杨桂文黄友华陈松林
水产学报 2024年4期
李开敏, 王 磊, 巩志宏, 王清滨, 李 华,杨桂文, 黄友华, 陈松林,3
(1. 中国水产科学研究院黄海水产研究所,海水养殖生物育种与可持续产出全国重点实验室,农业农村部海洋渔业与可持续发展重点实验室,山东 青岛 266071;2. 山东师范大学,山东 济南 250014;3. 崂山实验室海洋渔业科学与食物产出过程功能实验室,山东 青岛 266237;4. 莱州明波水产有限公司,山东 莱州 261418;5. 华南农业大学海洋学院,海洋生物资源保护与利用粤港澳高校联合实验室,广东省水产免疫与健康养殖工程技术研究中心,广东 广州 510642)
干扰素调节因子(IRFs)是一类细胞内转录因子,参与调节多种生物学过程,最开始被认为是调节干扰素(interferon,IFN)表达的转录因子[1];后来研究发现IRFs 还参与Toll 样受体和其他模式识别受体(pattern-recognition receptors,PRRs)引发的基因表达调控,从而证实了它们在宿主抵抗病原体中的核心作用[2-4]。此外IRFs 在造血细胞发育分化方面也发挥重要作用[5]。目前在脊椎动物中发现了11 个IRF 家族成员,哺乳动物中发现9 个成员(irf1~irf9)[6-7],鸟类中鉴定出irf1~irf10[8],在硬骨鱼斑马鱼(Danio rerio)中首次鉴定出irf11,说明鱼类具有完整的干扰素调节机制[9-12]。与哺乳动物相似,鱼类先天免疫系统中也有防御系统和调节机制[13],鱼类利用IFN 诱导系统对抗病毒感染[14-15]。研究表明,病毒感染诱导的鱼类IFNs 可增强抗病毒活性,并加速IFN 刺激基因(interferonstimulated gene,ISG)的表达以抑制病毒复制[16]。在鱼类中IRFs 家族在DNA 和RNA 病毒感染过程中发挥重要作用[17-22]。干扰素调节因子3 (interferon regulatory factors 3,irf3)是IRF 家族成员,在诱导病毒感染细胞中I 型IFN (IFN-α 和IFN-β)的表达上发挥重要作用,在结构上与irf7 同源[7-8],irf3在大多数组织和细胞中均呈组成型表达[23-24],主要存在于细胞质中[25]。病毒感染后,PRRs 启动信号转导并激活丝氨酸/苏氨酸激酶(TANK-binding kinase 1,TBK1),使irf3 的C 端富含丝氨酸区域特定几个丝氨酸和苏氨酸残基磷酸化,诱导IRF3和NF-κB 蛋白复合体从细胞质到细胞核的易位。易位到细胞核后,IRF3 和NF-κB 启动转录,导致经典的JAK-STAT 通路和干扰素刺激基因因子3(ISGF3)的激活[26-27]。研究报道,小鼠(Mus musculus)缺乏irf3 导致体内产生的IFN 急剧减少,因而更容易受到病毒感染[28]。目前,在大黄鱼(Larimichthys crocea)[18]、鲤(Cyprinus carpio)[19]、半 滑舌鳎(Cynoglossus semilaevis)[29]、牙鲆(Paralichthys olivaceus)[30]、尖吻鲈(Lates calcarifer)[31]、虹鳟(Oncorhynchus mykiss)[12]、鳜(Siniperca chuatsi)[32]等鱼类中开展了irf3 基因的克隆鉴定和初步表达分析。
斑石鲷(Oplegnathus punctatus)又名斑鲷、黑金鼓,属于鲈形目(Perciformes)石鲷科(Oplegnathidae)石鲷属(Oplegnathus),主要分布在西太平洋温带水域,具有显著的经济和生态价值[33]。斑石鲷在我国东南沿海均有分布,是我国名贵的海水养殖品种之一。随着国内斑石鲷养殖业集约化水平不断提高,从苗种到养成阶段各种病害频发,严重制约了斑石鲷的健康养殖,其中虹彩病毒病成为制约斑石鲷健康养殖的主要问题[34]。目前关于irf3 基因在斑石鲷抗虹彩病毒感染中的功能研究尚未见报道。
本研究以斑石鲷为对象,初步分析其irf3 基因的序列特征和进化地位,通过实时荧光定量PCR 检测了Opirf3 在斑石鲷各健康组织以及虹彩病毒感染后免疫组织中的表达水平;构建斑石鲷肾细胞系poly I: C 体外刺激模型,在细胞水平上检测Opirf3 的抗病毒免疫应答模式;在斑石鲷肾细胞上对Opirf3 进行siRNA 干扰后,检测Opirf3下游免疫相关基因的表达水平,以期实现对Opirf3 在斑石鲷抗虹彩病毒感染过程中免疫作用初探,旨在为斑石鲷抗病分子育种提供理论依据。
1 材料与方法
1.1 实验样品
正常组织样品 本实验所用的斑石鲷购自山东莱州明波水产有限公司,健康无病,体重为(150±15) g,实验前在水箱暂养1 周。随机选取4尾斑石鲷,麻醉后解剖取肝脏、脾脏、肾脏、头肾、肠道、鳃、皮肤、心脏、胃和脑组织,将组织立即放入盛有RNA 保存液的冻存管中,随后转移至-80 °C 冰箱保存。
虹彩病毒感染样品 上述健康斑石鲷每尾腹腔注射100 μL (109个拷贝数)虹彩病毒进行感染实验,对照组腹腔注射100 μL 磷酸缓冲溶液(PBS),在感染后0、1、4、7 和10 d 这5 个时间点取样,每个时间点感染组和对照组各随机选取5 尾斑石鲷,麻醉后解剖取其肝脏、脾脏和肾脏组织,立即放入盛有RNA 保存液的冻存管,存于-80 °C 冰箱保存。本感染实验所用的虹彩病毒由华南农业大学惠赠[35]。
斑石鲷细胞系 用于免疫刺激实验和siRNA 干扰的斑石鲷肾细胞系均来源于中国水产科学研究院黄海水产研究所青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室。
本研究获得了中国水产科学研究院黄海水产研究所实验动物管理和使用伦理委员会批准,实验过程中操作人员严格遵守中国水产科学研究院黄海水产研究所伦理规范,并按照中国水产科学研究院黄海水产研究所伦理委员会制定的规章制度执行。
1.2 斑石鲷RNA 提取和cDNA 合成
样品使用TRIzol 法提取RNA,实验方法严格参照说明书,用1%的琼脂糖凝胶电泳观察各样品RNA 条带的完整度,并用DNA/Proteins Analyzer P100 测定RNA 的浓度及纯度,选择条带完整、质量良好的RNA,进行cDNA 合成。使用Prime Script RT reagent Kit with gDNA Eraser 反转录试剂盒(TaKaRa,日本)进行cDNA 合成。反转录产物经PCR 反应验证质量,PCR 反应体系:cDNA 1 μL,TaKaRa ExTaq10 μL,正反引物各1 μL,ddH2O 7 μL;反应条件34 个循环:94 °C 30 s,58 °C 30 s,72 °C 30 s,经1%琼脂糖凝胶电泳,观察条带,选择质量好的cDNA 用于后续荧光定量实验。
1.3 Opirf3 序列分析
Opirf3 序列来源于本实验室的斑石鲷基因组数据库[36]。使用DNAman 软件预测Opirf3 基因的氨基酸序列;使用SWISS-PROT (https: //www.expasy.org/)预测Opirf3 基因分子质量和理论等电点;使用SMART 在线网站(https: //smart.embl.de/smart/set_mode.cgi?NORMAL=1)预测同源保守功能结构域。在NCBI 下载不同脊椎动物的Irf3 氨基酸序列,并用BLAST 进行同源比对;使用DNAman进行物种间Irf3 氨基酸序列多重比对;使用MAGE7软件的ClustalW 进行序列比对,采用邻接法(Neighbor-joining,NJ)构建系统发育进化树,设置Bootstrap method 重复1 000 次,来计算各分支的置信度。
1.4 Opirf3 的组织表达分析
使用Primer premier 5.0 软件,根据Opirf3 的CDS 区设计荧光定量引物(表1),以斑石鲷β-actin为内参基因,反转录得到的健康斑石鲷肝脏、脾脏、肾脏、头肾、肠道、鳃、皮肤、心脏、胃和脑组织的cDNA 为模板,使用AG 荧光定量试剂盒、 ABI 7500 Fast Real-time PCR 仪 (Applied Biosystems,美国)测定Opirf3 基因的表达水平。PCR 反应体系:10 μL SYBR mix、0.4 μL ROX、0.8 μL F (10 μmol/L)、0.8 μL R (10 μmol/L )、7 μL ddH2O、1 μL cDNA 模版。反应程序:95 °C 30 s,40 个循环:95 °C 5 s,60 °C 34 s。利用 2-ΔΔcT方法计算Opirf3 基因在各组织中的相对表达量。

表1 本研究所用的引物及其序列Tab. 1 Primers used in this study
1.5 虹彩病毒感染后Opirf3 在免疫组织中的表达分析
以斑石鲷β-actin为内参基因,反转录得到的虹彩病毒感染的肝脏、脾脏和肾脏组织的cDNA为模板,使用上述相同的荧光定量方法检测Opirf3在虹彩病毒感染的免疫组织中的相对表达水平。
1.6 斑石鲷肾细胞体外刺激实验
将单层培养、生长状态良好的斑石鲷肾细胞系接种到12 孔板上,24 h 后细胞覆盖率达到90%左右。将12 孔板的培养基吸弃去,1×PBS 洗涤3 次后换上新鲜的L15 培养基。不同浓度poly I: C 刺激斑石鲷肾细胞实验:向12 孔板的每个孔中分别加入终浓度为0、50、100 和200 μg/mL 的poly I: C,对照组加入等体积的PBS,将上述细胞样品放在24 °C 恒温培养箱中培养6 h 后,分别收集细胞样品,TRIzol 法提取RNA,制备定量模板。荧光定量PCR 检测poly I: C 刺激后斑石鲷肾细胞系中Opirf3 的相对表达水平。
1.7 siRNA 干扰实验
委托生工生物工程(上海)股份有限公司合成Opirf3 的siRNA,根据Opirf3 的ORF 区序列筛选到3 个siRNA 位点,分别为siRNA1、siRNA2 和siRNA3,siRNA 序列和Opirf3 免疫相关基因引物序列见表1。将适量生长状态良好的斑石鲷肾细胞系接种到2 个12 孔板中,在24 °C 恒温培养箱中培养至细胞贴壁且覆盖率达到30%~50%即可进行siRNA 转染实验。使用riboFECTTMCP 转染试剂盒进行Opirf3 的siRNA 转染,siRNA 转染终浓度为50 nmol/L,每孔转染体系:1×CP buffer 60 μL,CP Regent 6 μL,siRNA (siRNA1-3; NC; cy3)2.5 μL。3 个Opirf3-siRNA 位点重复6 个孔,NC重复4 个孔,用于检测荧光转染效率的cy3 有2个孔。转染后24 h 荧光显微镜下观察,判断转染效率;72 h 使用TRIzol 收集细胞,使用TRIzol 法提取RNA,制备定量模板,荧光定量PCR 检测3个位点的敲降效率。以敲降效率最高的位点转染的斑石鲷肾细胞的cDNA 为模板,荧光定量PCR检测irf3 免疫相关基因IFN-α、CD40、CD80、IL-6 和IL-1β的表达水平。
1.8 荧光定量PCR 引物扩增效率检测
荧光定量PCR 产物送青岛睿博兴科生物技术有限公司进行测序,测序成功后,将公司返还的PCR 纯化产物与pEASY®T1 载体[天根生化科技(北京)有限公司] 进行连接转化,严格按照说明书进行操作。挑取白色单克隆接种于含有氨苄(AMP+)的LB 液体培养基中,200 r/min,37 °C 培养15 h。用无内毒素质粒小提中量试剂盒[天根生化科技(北京)有限公司] 提取质粒,测定质粒浓度为100 ng/μL。参考北京擎科生物科技股份有限公司《绝对定量标准曲线制作方法》,将上述提取的质粒梯度稀释10~10 000 倍,作为荧光定量PCR 模板,上机进行荧光定量检测,得到对应的CT 值和拷贝数,从而制作标准曲线,标准曲线的斜率即为引物的扩增效率。
1.9 数据分析
实验数据采用SPSS 软件进行方差分析,组织表达数据采用单因素方差分析,P<0.05 为差异显著。其余实验数据采用独立样本t检验进行方差分析,*代表P<0.05,**代表P<0.01。
2 结果
2.1 Opirf3 序列分析
Opirf3 cDNA 开放阅读框为1 362 bp,编码453 个氨基酸,预测蛋白分子质量为50.0 ku,理论等电点为4.97。SMART 软件预测显示Opirf3有1 个IRF 结构域和1 个IRF-3 结构域,分别位于第1~108 和250~430 位氨基酸(图1)。

图1 斑石鲷与其他物种Irf3 氨基酸序列多重比对黑色区域. 氨基酸位点相似性为100%;粉色区域. 氨基酸位点相似性≥75%;蓝色区域. 氨基酸位点相似性≥50%。Fig. 1 Multiple alignment of the deduced amino acids of Irf3 between O. punctatus and other species Black region. amino acid site similarity is 100%; pink region. amino acid site similarity≥75%; blue area. amino acid site similarity≥50%.
2.2 Opirf3 多序列比对与系统进化分析
经BLAST 比对发现,Opirf3 编码的氨基酸序列与其他硬骨鱼类irf3 编码的氨基酸序列具有较高的相似性,与条石鲷(O.fasciatus)、花鲈(Lateolabrax japonicus)、鳜和条纹狼鲈(Morone saxatilis)的相似性分别为91.90%、84.47%、84.26%和84.01%;与两栖动物非洲爪蟾(Xenopus laevis)的相似性最低,为31.08%;与人(Homo sapiens)、小鼠(Mus musculus)和黑猩猩(Pan troglodytes)等的相似性较低,分别为31.41%、31.47%和31.41%(表2)。
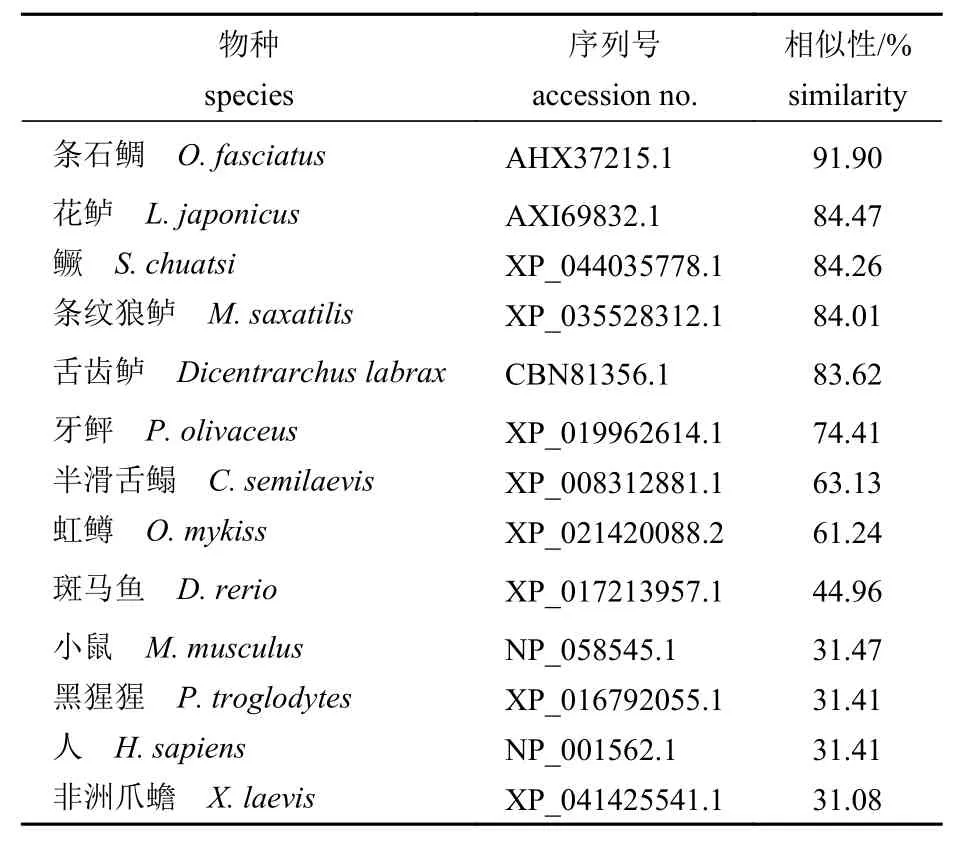
表2 斑石鲷与其他脊椎动物Irf3 的氨基酸序列相似性Tab. 2 Degree of homology between Irf3 of O. punctatus and other vertebrates
使用DNAman 对表1 所述氨基酸序列进行多重比对,发现不同物种间Irf3 氨基酸序列非常保守(图1)。为探究确定斑石鲷Irf3 在动物进化过程中的地位,利用MAGE 7 软件,通过Neighborjoining 法构建了脊椎动物的系统进化树。系统分析结果显示,哺乳动物小鼠、人和黑猩猩的Irf3聚为一支,两栖类非洲爪蟾单独为一支,斑石鲷Irf3 与其他硬骨鱼类聚为另一支。说明在进化上斑石鲷Irf3 与哺乳动物的亲缘关系较远,而与硬骨鱼条石鲷的亲缘关系最近,与牙鲆、半滑舌鳎等的亲缘关系较近(图2)。
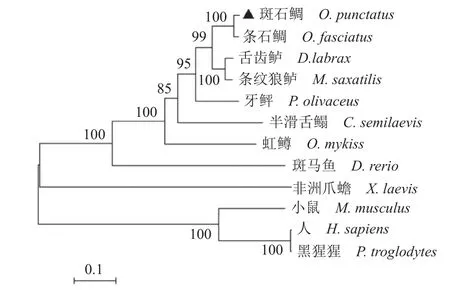
图2 IRF3 氨基酸序列系统进化树分析Fig. 2 Phylogenetic analysis of IRF3 amino acid sequence
2.3 Opirf3 组织表达分析
为了研究Opirf3 在斑石鲷各组织中的表达模式,通过荧光定量PCR 检测健康斑石鲷肝脏、鳃、心脏、皮肤、脾脏、肠道、脑、肾脏、胃和头肾组织中Opirf3 相对表达水平。结果显示,Opirf3在斑石鲷各组织中均有表达,但表达水平有差异(图3),在肝脏中的表达水平显著高于其他组织。
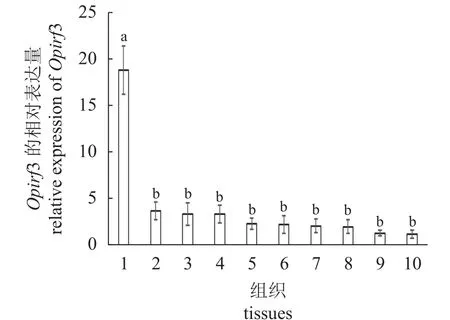
图3 斑石鲷组织中Opirf3 的相对表达水平1. 肝脏,2. 鳃,3. 心脏,4. 皮肤,5. 脾脏,6. 肠道,7. 脑,8. 肾脏,9. 胃,10. 头肾,n=4;数值用平均值±标准差表示,不同字母表示各表达水平之间差异显著(P<0.05),下同。Fig. 3 Relative expression of Opirf3 in O. punctatus tissues 1. liver, 2. gill, 3. heart, 4. skin, 5. spleen, 6. intestine, 7. brain, 8. kidney,9. stomach, 10. head kidney, n=4; values are expressed bymean±SE, and different letters inditace significant differences in expression levels(P<0.05); the same below.
2.4 虹彩病毒感染后Opirf3 在免疫组织中的表达分析
斑石鲷腹腔注射SKIV-SD 后,于不同时间点取肝脏、脾脏和肾脏3 种免疫组织,通过荧光定量PCR 检测Opirf3 的表达水平变化。结果显示,肝脏、脾脏和肾脏组织中Opirf3的表达水平在注射时间为7 和10 d 时均显著升高,但不同组织中Opirf3 的相对表达水平略有差异(图4)。注射时间为7 和10 d 时,肝脏中Opirf3 相对表达水平分别为0 d 的4.6 倍、5.7 倍;脾脏中Opirf3 相对表达水平分别为0 d 的7.3 倍和7.0 倍;肾脏中Opirf3 的相对表达水平分别为对照组的8.9 倍和14.2 倍。

图4 虹彩病毒感染后不同时间点肝脏(a)、脾脏(b)和肾脏(c)中Opirf3 的表达水平PBS 组4 个平行(n=4),SKIV-SD 组5 个平行(n=5);数值用平均值±标准误表示,*. P<0.05,**. P<0.01,***. P<0.001,下同。Fig. 4 Relative expression of Opirf3 in the liver (a),spleen (b) and kidney (c) at difference time points after SKIV-SD infection There were 4 parallels (n=4) in PBS group and 5 parallels (n=5) in SKIVSD group. Values are represented by mean±SE, *. P<0.05, **. P<0.01,***. P<0.001; the same below.
2.5 斑石鲷肾细胞体外刺激实验
为了进一步探讨Opirf3 在斑石鲷抗病毒感染中发挥的作用,本实验设计了斑石鲷肾细胞系的体外刺激实验。分别用不同浓度的poly I: C 刺激斑石鲷肾细胞系,6 h 后收集细胞,提取细胞RNA 制备定量模板,荧光定量PCR 检测poly I: C刺激后肾细胞中Opirf3 的表达水平,PBS 作为对照组。结果显示,不同浓度poly I: C 刺激后,肾细胞中Opirf3 的表达水平均显著升高(图5),在刺激浓度为50、100 和200 μg/mL 时,Opirf3 的表达水平分别是刺激浓度为0 μg/mL 时的80.8、86.8 和60.6 倍。

图5 不同浓度poly I: C 刺激6 h 后斑石鲷肾细胞中Opirf3 的表达水平每组3 个平行,n=3。Fig. 5 Expression level of Opirf3 in kidney cells at 6 h after stimulation by different concentrations of poly I: C There were 3 parallel per group, n=3.
2.6 siRNA 干扰
由转染效率图可知,Opirf3 的siRNA 在肾细胞中的转染效率较高,为90%以上,实验结果可靠(图版)。荧光定量PCR 检测发现,3 个位点均有敲降,其中siRNA-3 对Opirf3 的干扰效果最为明显,较对照组siRNA-NC 显著下降(下调了30%,图6)。因此,实验检测siRNA-3 干扰后Opirf3 免疫相关基因的表达,以期探究Opirf3 对下游免疫相关基因表达的影响。在siRNA 干扰Opirf3 后,分 别 检 测 了 干 扰 素α (interferon alpha,IFN-α)、CD40 抗原(CD40 antigen,CD40)、CD80 抗原(CD80 antigen,CD80)、白细胞介素6 (interleukin 6,IL-6)和白细胞介素1β (interleukin 1 beta,IL-1β)的表达水平,结果显示,IFN-α、CD40、CD80 和IL-1β的表达水平较对照组siRNA-NC 显著下降,IL-6表达水平较对照组显著上调(图7)。

图6 不同靶点siRNA 对Opirf3 的干扰效果1. siRNA-NC,2. siRNA-1,3. siRNA-2,4. siRNA-3,siRNA-NC组4 个 平 行(n=4),siRNA-1、siRNA-2 和siRNA-3 组6 个 平 行(n=6)。Fig. 6 Changes in the expression of three sites after knocking down Opirf3 1. siRNA-NC, 2. siRNA-1, 3. siRNA-2, 4. siRNA-3, there were 4 parallel (n=4) in the siRNA-NC group and 6 parallel (n=6) in the siRNA-1,siRNA-2 and siRNA-3 groups.
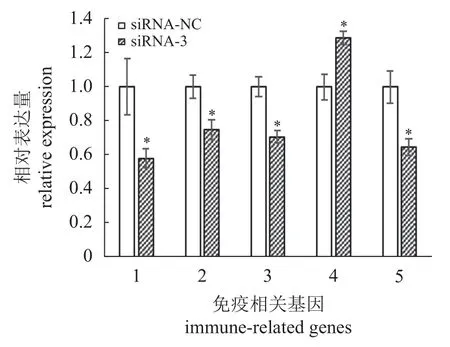
图7 Opirf3 siRNA 干扰后免疫相关基因在斑石鲷肾细胞中的表达水平1. IFN-α,2. CD40,3. CD80,4. IL-6,5. IL-1β,siRNA-NC 作为阴性对照,siRNA-NC 组4 个平行(n=4),siRNA-3 组6 个平行(n=6)。Fig. 7 The relative expression levels of immune-related genes in O. punctatus kidney cells after Opirf3 siRNA treatment 1. IFN-α, 2. CD40, 3. CD80, 4. IL-6, 5. IL-1β, siRNA-NC as a negative control, there were 4 parallel (n=4) in the siRNA-NC group and and 6 parallel (n=6) in the siRNA-3 group.
3 讨论
IRF 家族的功能由特定的蛋白质结构域决定[32]。本研究预测了斑石鲷Irf3 的蛋白结构,发现N 端有一个IRF 结构域,作为DNA 结合域(DBD),这一结构域在硬骨鱼和哺乳动物中略有差别,在哺乳动物中,Irf3 的DBD 有5 个色氨酸重复序列,而硬骨鱼中仅有4 个间隔的色氨酸重复序列[3,9]。该重复序列是Irf3 与启动子元件结合必不可少的,它们可以形成螺旋-转角-螺旋结构而与下游靶基因启动子中的IRF 调节元件(IRFE)和IFN 刺激反应元件(ISRE)共有序列结合[37]。在C 端是IRF3结构域,能够激活双链RNA 激活因子1 并防御病毒入侵[38]。这些结构特征表明Opirf3 在斑石鲷Ⅰ型干扰素激活和抗病毒免疫方面发挥重要作用。
斑石鲷组织特异性表达分析显示,Opirf3 在斑石鲷肝脏、鳃、心脏、皮肤、脾脏、肠道、脑、肾脏、胃和头肾组织中均有表达,在肝脏中的表达水平最高。这种普遍存在的表达模式,与其他已报道的鱼类相似,半滑舌鳎、虹鳟、尖吻鲈和花鲈中irf3 在各组织中均有表达,但不同组织中的表达水平略有差异,尖吻鲈中irf3 在脾脏和肾脏中表达水平最高,花鲈中irf3 在肝脏中的表达水平最高[12,23,29,31]。
poly I: C 类似于双链RNA,是一些病毒的关键病原体成分[39],研究表明病毒和poly I: C 处理诱导硬骨鱼中irf3 转录。花鲈中,赤点石斑鱼神经坏死病毒(RGNNV)和poly I: C 刺激后,细胞和组织中irf3 的表达水平均显著上调;鳜中poly I: C刺激后,显著上调了免疫相关组织中irf3 的表达水平;半滑舌鳎感染细胞肿大虹彩病毒(Megalocytivirus)后,irf3 在免疫组织中的表达水平显著上升;尖吻鲈感染神经坏死病毒RGNNV 后,脑和肝脏、脾脏和肾脏中irf3 的表达水平显著上升[23,29,31-32]。本研究中SKIV-SD 感染斑石鲷7 d 后,Opirf3 在肝脏、脾脏和肾脏中的表达水平显著上调。以上结果提示,irf3 在硬骨鱼抗病毒感染过程中发挥重要作用。
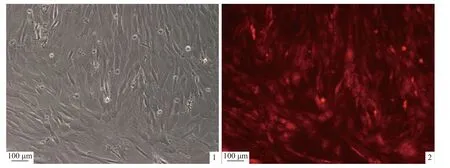
图版 斑石鲷肾细胞siRNA 转染效率图1. 明场观察; 2. 红色荧光观察。Plate Map of siRNA transfection efficiency in kidney cells of O. punctatus 1. brightfield observation; 2. red fluorescence observation.
IRF3 是硬骨鱼中一种重要的转录因子,调节I 型干扰素的表达以抵御病毒感染[9,12]。在花鲈脑细胞中过表达的irf3 显著抑制了RGNNV 复制。鳜中,irf3 的过表达刺激干扰素-β 启动子和含有ISRE 的启动子的活性。牙鲆中,poly I: C 刺激下过表达的irf3 极大提高了Ⅰ型IFN 的转录活性[23,32,40]。这些研究结果表明,硬骨鱼irf3 参与了病毒感染的先天免疫应答,并且诱导产生Ⅰ型干扰素以应对病毒感染[23]。在斑石鲷肾细胞中对Opirf3 进行siRNA 干扰后检测免疫相关基因,发现IFN-α显著下调,证明Opirf3 直接调控Ⅰ型干扰素信号通路。检测其他免疫相关基因发现,CD40、CD80、IL-6 和IL-1β被不同程度的激活或抑制。其中CD40 是表达于多种细胞表面的跨膜糖蛋白,属于肿瘤坏死因子受体超家族,在生发中心应答、T 依赖抗原应答和触发促炎症反应等发面发挥重要作用[41-44]。CD80 是免疫球蛋白超家族成员,调控T 细胞适度活化所必须的膜糖蛋白,若表达失调将导致一系列免疫疾病的发生[45]。IL-6和IL-1β在免疫和癌症等方面发挥着重要的作用[46]。此外研究证明,irf7 也是Ⅰ型IFN 表达的关键调控因子,在硬骨鱼的抗病毒反应中发挥重要作用[327],主要在免疫组织中表达[24]。半滑舌鳎感染细胞肿大虹彩病毒(Megalocytivirus)后,irf7 在免疫组织肝脏、脾脏和肾脏中的表达水平显著上调[29];神经坏死病毒(RGNNV)感染尖吻鲈后,脑和肝脏、脾脏等免疫组织中irf7 的表达量均显著上调[31]。本团队在前期研究中克隆了irf7 序列,并发现irf7 在斑石鲷感染虹彩病毒后表达量升高[47]。斑石鲷irf3 和irf7 的结构存在相似性,活化的irf3 形成同二聚体或与irf7 形成异二聚体,与 NF-κB、转录共刺激分子 CBP /p300 等形成全复合体,进入细胞核启动Ⅰ型 IFN 的 转录[3,47]。本研究对Opirf3 基因进行了鉴定和表征;分析了斑石鲷感染虹彩病毒后免疫组织的表达模式,同时还进行了体外细胞刺激实验和siRNA 实验。以上实验结果初步表明,虹彩病毒感染后,斑石鲷免疫组织和细胞中Opirf3 基因的表达上调,进而激活Ⅰ型IFN 通路来抵抗病毒对宿主细胞的侵袭。但关于斑石鲷Opirf3 介导的Ⅰ型IFN 通路的激活机制仍需要后续研究。本实验为斑石鲷抗虹彩病毒病的研究奠定一定的理论基础,也为斑石鲷的分子抗病育种提供一定的理论参考依据。
(作者声明本文无实际或潜在的利益冲突)
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13 08:59:28
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:06
中成药(2018年12期)2018-12-29 12:25:44
中央民族大学学报(自然科学版)(2018年3期)2018-11-09 01:16:32
中成药(2017年6期)2017-06-13 07:30:35
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:48:47
当代医药论丛(2017年22期)2017-04-12 06:29:43
中国卫生标准管理(2015年15期)2016-01-15 02:58:43
医学研究杂志(2015年4期)2015-06-10 06:42:43
当代畜禽养殖业(2014年11期)2014-02-27 08:00:01
