
不同光强下琼岛杨幼苗生长和光合特性
2024-05-11 00:00:00饶丹丹韩豫吴二焕陈彧
山西农业大学学报(自然科学版) 2024年1期






摘要:[目的]本文旨在探究不同光照强度对琼岛杨(Populus qiongdaoensis)幼苗生长和光合特性的影响。[方法]以1. 5 年生琼岛杨幼苗为试验材料,设置全光100%(CK)、全光70%(RI70%)、全光40%(RI40%)和全光10%(RI10%),处理12 个月后,分析不同光强对琼岛杨幼苗生长和光合特性的影响。[结果]成活率、地径增长量、叶片数、叶长、叶宽、叶面积和单叶质量均随着光强的减弱而降低,株高增长量和叶片数增长量呈先增后减趋势,RI70% 和RI40% 处理的琼岛杨幼苗的成活率、株高、地径和叶片数均显著高于RI10% 处理;RI70% 处理的根生物量、根生物量比和根冠比显著高于RI40% 和RI10% 处理,而RI70% 和RI40% 处理的根、茎、叶生物量和总生物量均显著高于RI10%;随着光照强度的降低,叶片叶绿素a、叶绿素b 和总叶绿素含量逐渐增大,类胡萝卜素含量随之减小;净光合速率(Pn)随光照强度的减小而显著下降,在RI70% 光强下琼岛杨幼苗叶片的Pn 最大,分别是RI40% 和RI10% 的1. 72倍和2. 16 倍;叶片数增长量与株高增长量、地径增长量呈显著正相关关系,净光合速率与类胡萝卜素含量(Car)和蒸腾速率(Tr)呈显著正相关,气孔限制值(Ls)与胞间CO2浓度(Ci )和气孔导度(GS)呈显著负相关。[结论]琼岛杨幼苗能忍耐一定程度遮阴,适度遮阴可促进幼苗地上和地下部分的生长并提高其气体交换能力,但在弱光胁迫下幼苗难以成活和生长,相对光强70% 下琼岛杨幼苗的适应性最强。
关键词:琼岛杨; 幼苗; 光照强度; 光合特性
中图分类号:S718.43 文献标识码:A 文章编号:1671-8151(2024)01-0061-09
光照是影响植物光合作用和生长发育的重要因素,光环境对木本植物林下幼苗的生长发育、形态特征、光合特性以及生物量等方面有重要影响[1]。当光环境发生变化,植物可以通过改变植株自身形态性状、叶片形态、生物量累积和光合指标等来适应外部生境变化[2-3],表现出表型可塑性[4]。通过遮光处理对红砂幼苗生物量积累和光合特性影响的研究发现,15% 遮光度可促进红砂幼苗生长和光合作用[1];金丝李幼苗通过在形态及光合生理上调节来适应光环境的变化[4];风吹楠幼苗在不同光强下叶片形态和光合参数表现出显著差异[5]。植物可通过调节光合作用来适应光照强度的变化[6],同时光合作用是植物生长和生物量积累的重要途径,生物量反映了植物的生长状况,光照过弱或过强都会导致供光合作用的能量不足,不利于生物量积累[7]。林下幼苗受到生境变化的影响表现更为敏感[8],林下幼苗对光环境的适应能力决定了它的分布地区和物种丰度[7]。遮阴是通过改变环境因子中光照强度,降低到达地表的太阳总辐射强度,相应的改变地表温度和相对湿度[8]。大量研究表明,适度遮阴有利于木本植物幼苗的生长发育、形态特征、光合特性及生物量累积等[1,3-10],因此,选择适宜的光照强度对苗木培育以及人工林的管理和立地选择等都具有重要指导意义[11-13]。
琼岛杨(Populus qiongdaoensis)为杨柳科(Salicaceae)杨属(Populus)热带速生杨树,海南省重点保护植物,其木材可制作家具、纸张等,苗木可作为生态修复树种应用于海南热带雨林国家公园生态修复建设,经济价值和生态价值极高。近年来,由于其受自然更新能力差和自然环境变化等原因的影响,琼岛杨天然居群个体急剧减少,集中分布于海南热带雨林国家公园霸王岭分局和鹦哥岭分局等高海拔原始林和次生林中,成年植株分布于森林冠层的顶端,对光照的需求大,而幼苗零散生长于成树林下散射光中,林下幼树极少[14]。鉴于原生境的光照强度对幼苗的天然更新有较强的影响[12],因此探究琼岛杨幼苗在生长发育过程对光照强度的需求和光强适应性具有重要的理论和现实意义。目前,关于琼岛杨的研究集中于迁地培育[15]、高压繁殖[16]、嫩枝扦插[17]、种子苗期节律[18]、愈伤组织培养[19]和基础生物学[20-22]等方面,而有关琼岛杨幼苗光合特性的研究尚未见报道。本研究通过测定不同强度光照环境下琼岛杨幼苗生长特性、叶片功能性状、光合色素和光合特性,探究不同光照强度对琼岛杨幼苗生长和光合特性的影响,以期为琼岛杨种质资源保护和利用、濒危原因探究、回归种植和生态修复等提供理论依据。
1 材料与方法
1. 1 试验地概况
试验地点设在海南省林业科学研究院(海南省红树林研究院)科研基地苗圃内,该基地位于海南省海口市(19°52′21′′ N、110°28′59′′ E),平均海拔30. 4 m,热带季风气候,年均气温23. 8 ℃,年均日照1750~2000 h,年均降雨量1 697. 8 mm,降雨多集中在雨季(6-10 月)[17]。
1. 2 试验材料与设计
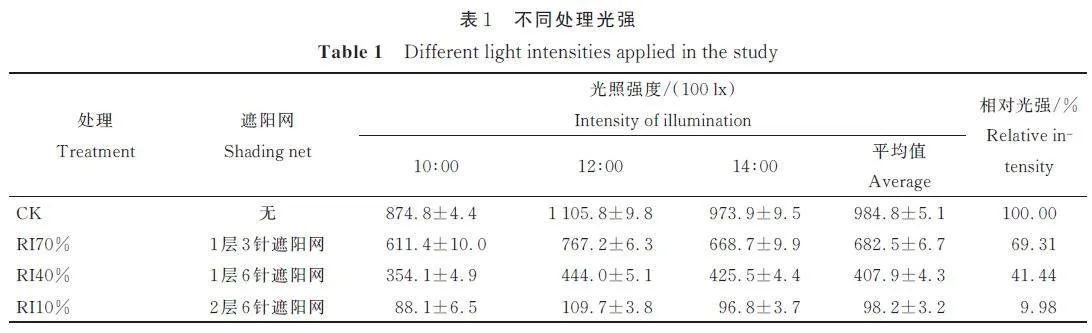
选择生长健壮、长势基本一致、同一家系琼岛杨1. 5 年生实生苗(苗高12. 3 cm、地径1. 62 mm),于2021 年5 月,移栽于上口径15 cm,盆高14 cm 的花盆中,以红心土+泥炭土+珍珠岩+椰糠(体积比2∶1∶1∶1)为栽培基质,每盆栽植1 株幼苗,浇透定根水,置于RI70% 下缓苗1 月后置于遮雨棚中进行遮光处理,苗木管护措施保持一致,遮光处理12 个月;以全光100% 为对照(CK),设置3 种光照强度处理(具体见表1),经照度计实际测量,3 种处理相对光照强度约为全光70%(RI70%)、全光40%(RI40%)和全光10%(RI10%),采用规格为3针和6 针的黑色遮阳网搭设遮荫棚,每个处理15盆,试验重复3 次。
1. 3 指标测定方法
3 种光强处理12 个月后测定生长指标、叶片指标和光合指标,生长指标采用直尺、游标卡尺和叶面积仪测定,生物量采取电热恒温鼓风干燥箱和分析天平(0. 001 g),光合指标采用LI-6800 便携式光合作用测量系统(LI-COR,美国)进行测定,光合色素采用紫外分光光度计测定。
1. 3. 1 成活率、生长特征和叶片特征测定
采用直尺和游标卡尺测定植株的株高(Height,H)和地径(Diameter,D),统计叶片数(Leaf number,LN)和分支数(Branch number,BN),并计算各指标增长量(增长量=实验结束时苗木指标−实验前苗木指标)。
各处理随机选6 株平均木,分别从每株幼苗上剪取功能叶片3 片(取中上部完全展开的成熟叶),采用手持式叶面积仪(YMJ-B,浙江托普云农科技服份有限公司,中国)测定叶长(Leaf long,LL)、叶宽(Leaf width,LW)、叶面积(Leaf area,LA)和叶片长宽比(Aspect ratio,LB)。
采用分析天平测定称量叶片鲜重(Freshweight,FW),置于烘箱100 ℃杀青1 h,再于85 ℃烘干12 h 至恒重,称量单叶片干重(Dry weight,Dw),并计算叶片含水量(Leaf water content,LWC)和比叶面积(叶面积/叶干质量;Specificleaf area,SLA),公式如下:
1. 3. 2 生物量测定
每个处理随机选择6 株平均木作为生物量测定对象,整株带回实验室,置于烘箱中100 ℃杀青1 h,再把温度调到85 ℃,烘干12 h 直至恒重,分别称量根茎叶各部分生物量和总生物量(Total biomass,TB),并计算根生物量比(根生物量/总生物量;Root mass ratio,RMR)和根冠比(根生物量/地上部分生物量;Root to shoot ratio,R/S)。
1. 3. 3 气体交换参数测定
于2023 年6 月的晴天,在上午的8:30-11:30进行,每个处理选取3 株琼岛杨幼苗中上端完全展开的3 片成熟叶,采用LI-6800 便携式光合测定系统(LI-COR,美国)测定气体交换参数,记录叶片净光合速率(Net photosynthetic rate,Pn)、蒸腾速率(Transpiration rate,Tr)、气孔导度(StomatalConductance,Gs)和胞间CO2 浓度(IntercellularCO2 concentration,Ci)等参数,并计算气孔限制值(Ca/Ci;Stomatal limitation value,Ls),水分利用效率(Pn/Tr;Water use efficiency,WUE)[4]。
1. 3. 4 光合色素测定
每处理随机选取3 株琼岛杨成熟叶片,低温避光保存带回实验室用于叶片含水量及光合色素含量测定。光合色素含量测定方法采用乙醇提取法[23],用紫外分光光度计在470、649、665 nm 波段测量吸光值(A),叶绿素a(Chla),叶绿素b(Chlb),叶绿素(Chl)和胡萝卜素(Car)计算公式如下:
Chla 含量/(mg·g-1)=( 13.95 × A665 - 6.88 ×A649 )× V × n/( m × 1000 ) (2)
Chlb 含量/(mg·g-1)=( 24.96 × A649 - 7.32 ×A665 )× V × n/( m × 1000 ) (3)
Chl 含量/(mg·g-1)=Chla+Chlb (4)
Car 含量/(mg·g-1)=[ 1000 × A470 - 2.05 ×( 13.95 × A665 - 6.88 × A649 )- 114.8 ×( 24.96 ×A649 - 7.32 × A665 ) ]×V× n/( m× 1000 ) ( 5)
式中,V 为提取液体积,n 为稀释倍数,m 为样品质量(g)。
1. 4 数据处理
采用Excel 2010 和SPSS 22. 0 软件进行统计分析,运用单因素方差分析(ANOVA)中的最小显著性差异(LSD)法进行显著性差异分析,运用皮尔逊(Pearson)法进行相关性分析,各参数以平均值±标准误(Mean±SE)表示。
2 结果与分析
2. 1 不同光强对琼岛杨幼苗生长影响
由表2 可见,光照强度对琼岛杨幼苗成活率、株高、地径和叶片数有显著影响。琼岛杨幼苗各处理间的成活率以及株高、地径和叶片数的增长量呈显著差异,分支数增长量无显著差异,同时,随着光照强度的降低,成活率显著降低,RI10% 处理的幼苗出现较多的死苗,RI100% 处理时,植株全部死亡。幼苗株高、地径和叶片数的增长量随光强的降低呈先增大后减小的趋势;RI40% 处理的株高增长量和叶片数增长量最大,分别为RI10% 的4. 07 倍和3. 61 倍,RI70% 的成活率和地径增长量最大且显著高于RI10%,分别较RI10%增加了0. 51 倍和5. 87 倍。
2. 2 不同光强对琼岛杨幼苗叶片形态影响
3 种光强处理之间琼岛杨幼苗叶片的叶长、叶宽、单叶面积、叶片数、单叶质量、比叶面积均差异显著,叶片长宽比和含水量差异不显著,随着光照强度的降低,叶长、叶宽、单叶面积和单叶质量均显著降低(表3)。遮阴胁迫的RI10% 处理下叶长、叶宽、叶面积、单叶质量均显著低于RI70% 和RI40% 处理,而RI40% 处理的叶长、叶宽、单叶质量低于RI70% 处理,但无显著差异。琼岛杨幼苗叶片形态的变化体现了对光照强度的适应性,光强对其叶片长度、宽度、叶面积和单叶质量的影响明显,适度遮阴下可促进幼苗叶片生长,强遮阴处理对幼苗叶片生长有一定程度的胁迫作用。
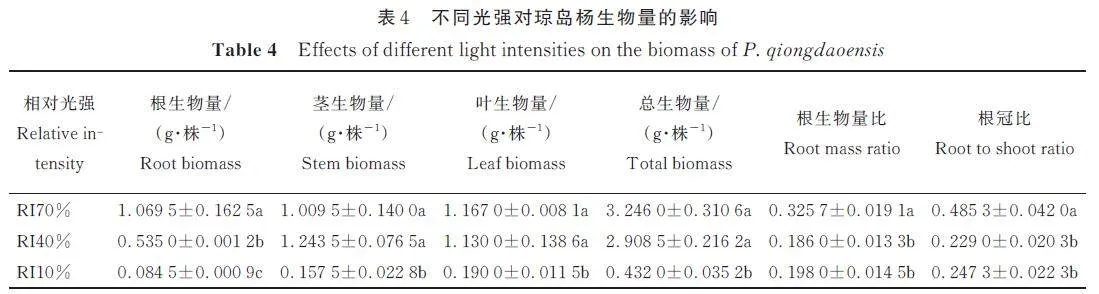
2. 3 光照强度对琼岛杨幼苗生物量分配影响
由表4 可见,3 种光强处理下,琼岛杨根、茎、叶以及总生物量,根生物量比和根冠比均呈显著差异,表明光照强度对琼岛杨幼苗生物量有明显影响。随着遮阴的增加,幼苗的根、叶、总生物量均呈减小的趋势,茎生物量呈先增大后减少的趋势。RI70% 处理的根生物量、根生物量比和根冠比最大,显著高于RI40% 和RI10% 处理,根生物量分别是RI40% 和RI10% 的2. 0 倍和12. 7 倍,这说明较高强度光照有利于琼岛杨幼苗根部生长;RI70% 和RI40% 处理的根、茎、叶生物量和总生物量均显著高于RI10%,表明强遮阴胁迫不利于琼岛杨幼苗根茎叶的生长,适度遮阴一定程度上能促进幼苗地上和地下部分的生长。
2. 4 不同光强对琼岛杨幼苗叶片光合色素影响
由表5 可见,琼岛杨幼苗在不同光照强度下生长1 年后,其成熟叶片的Chla、Chlb、Chl 和Car 含量,以及Chla/Chlb 和Car/Chl 差异显著,因此,光照强度对琼岛杨幼苗叶片光合色素产生了明显影响。随着光照强度的降低,叶片的Chla、Chlb 含量和Chl 含量逐渐增加,这表明,弱光条件下琼岛杨幼苗叶片能通过增加Chl 含量来应对弱光胁迫,增加对光能的利用;相反地,Chla/Chlb 随光强的降低而下降,RI70% 处理的Chla/Chlb 显著高于其它处理。Car 含量随光强的降低而降低,Car/Chl 逐渐降低,其中RI70% 处理的Car/Chl 显著高于RI40% 和RI10%。
2. 5 不同光强对琼岛杨幼苗叶片气体交换参数影响
由表6 可见,光照强度对琼岛杨幼苗叶片气体交换参数有显著影响。Pn 随遮光程度的增加而显著减小,在RI70% 处理下琼岛杨幼苗叶片的Pn 最大,分别是RI40% 和RI10% 的1. 72 倍和2. 16 倍。Tr 随遮光强度的增加,先大幅度降低,后缓慢降低,RI70% 光强下叶片Tr最大,且显著高于RI40%和RI10%。Ci 和Gs 与Pn 呈相同趋势。WUE 随遮光程度的增加逐渐降低,Ls 随遮光程度的增加而显著降低,在强遮阴胁迫RI10% 下的WUE 和Ls最大,且显著高于RI70% 和RI40%。
2. 6 不同指标相关性分析
琼岛杨幼苗生长和光合特性随光照环境而变化,各指标间表现出一定程度的相关性(表7)。生长性状中D 与TB、LA 呈显著正相关关系,与DW呈极显著正相关关系,DW 与TB、LA 呈显著正相关关系;光合特性中,Pn 与生长指标呈正相关,与Car、Tr、Ci 和Gs 均呈正相关,与Chl、WUE、Ls 呈负相关,并与Car 和Tr 呈显著正相关关系,Ci 与Gs 呈显著正相关,WUE 和Chla 呈显著正相关关系,Ls与Ci 和Gs分别呈极显著负相关和显著负相关。
3 讨论
适宜的光强有利于植物幼苗的生长发育,植物的光合作用能力强,能够迅速合成养分,促进有机物合成和生物量累积,从而促进植物的生长发育。喜阳植物更适应较强的光照环境,如长序榆[24]、乐昌含笑[25]等幼苗在较高光强下生长和光合更强;喜阴植物能耐较高的遮阴度,如谢君魔芋[26]、景东翅子树幼苗[27];中性植物如棱角山矾幼苗在相对光强50% 下生长最佳[26]。表型可塑性是植物通过生长发育和形态特征调节来适应环境的变化[28]。一般来说,适度遮阴有利于植物的生长发育[25]。光照强度对琼岛杨幼苗生长发育有显著影响,光照强度不同,琼岛杨幼苗的株高、地径、叶片数、叶长、叶面积等形态特征参数随之发生显著变化,表明琼岛杨幼苗具有表型可塑性。随着光强的降低,苗木的地径、叶片数、叶长、叶宽、叶面积和单叶质量逐渐降低,在一定遮阴条件下,植物幼苗通过伸长枝条、增大叶片面积和质量来获取更多的光能,以满足其生长需求。因此,本试验发现,较高光强环境下,琼岛杨幼苗地径生长量、叶片面积明显增大,相对光强40% 时幼苗则偏向于株高生长,相对光强10% 时幼苗株高增长量、地径增长量和叶片均明显减小,这表明,70% 光照强度有利于琼岛杨幼苗植株和叶片的生长发育,但幼苗对强遮阴胁迫的适应性较差。这与不同光照强度下红砂[1]、火力楠[28]幼苗形态生长对光强的响应具有一定的相似性。
光合作用是植物合成有机物和能量的根本途径,是累积生物量和生理代谢的基础,植物在适宜的光照强度下光合作用更强,根茎叶各部分生物量和总生物量累积较多[11,13,29]。琼岛杨幼苗根茎叶各部分生物量以及总生物量等参数均随光强变化以适应光环境的变化,与弱光胁迫相比,适宜光强下幼苗的根生物量、叶生物量、总生物量、根生物量比和根冠比显著增大,表明较高的光强有利于其根茎叶的生长,随着光强的降低,光合作用累积生物量逐渐减少,幼苗通过增加茎的生长适应光环境的改变,因此分配至茎的生物量先增大后降低,分配至根部和叶部的生物量逐渐降低,相对光强为70% 有利于琼岛杨幼苗地上地下部分的生长和生物量的累积。
植物叶片的光合色素含量为响应光强的变化,不同光强下各叶绿体色素指标差异明显[4,31],本研究中琼岛杨幼苗叶片Chla、Chlb 和Chl 含量随光照强度降低而增加,这与代大川等[31]对光强对桢楠幼苗的影响研究结果基本一致。遮阴有利于光合色素的积累,这可能是因为光合色素积累会影响叶片对光能捕获能力,植物可通过增加叶片Chl 含量来提高光能捕获,Chlb 含量的增加可提高植物对蓝紫光的利用效率,增加光能的吸收利用,来响应弱光环境。同时,Car 具有光破坏防御的功能[30],琼岛杨幼苗随着光照强度的增加,Car 含量逐渐增大,在相对光强70% 时达到最大,这可能是为防御高光照对叶片的伤害。
光照强度对木本植物幼苗的光合气体交换有显著影响[28],本研究表明,琼岛杨幼苗叶片的Pn、Tr、Ci 和GS 随光照强度的降低而显著降低,而WUE 和Ls 随之增大,这与乐昌含笑[25]等阳性树种的变化趋势一致,相对光强70% 时幼苗叶片的Pn、Tr、Ci 和GS 均最大,显著高于相对光强40% 和10%,WUE 和Ls 则显著低于相对光强40% 和10%,这说明相对光强70% 下的琼岛杨幼苗气体交换能力强于另外2 个光强,表现出更强的光合作用,植株生长迅速,这表明适度遮阴有利于其幼苗的光合作用和气体交换。
分析各指标相关性发现,Pn 与生长指标之间呈正相关,Chla 含量与形态生长指标呈负相关,蒸腾速率和净光合速率呈显著正相关,这是因为蒸腾速率的提高不仅能加快水分和物质传输,还可促进CO2通过气孔进入植物体内,进而提高光合速率[12],胞间CO2 浓度与气孔导度呈显著正相关,气孔限制值与胞间CO2 浓度和气孔导度呈极显著负相关和显著负相关,这是因为气孔限制开度影响外界CO2进入细胞内,从而影响胞间CO2浓度和气孔导度。诸多报道表明,适当弱光环境对植物苗木生长有利,本研究结果也支持了这一观点。
结合本文研究和课题组前期调查研究推测,随着琼岛杨幼苗不断生长发育,其对光照需求越来越多,而在乔灌木高度郁闭的林下,虽然种子大量萌发,但幼苗保存率低,底层幼苗和幼树获得的光能不足,生长发育被抑制,导致种群自然更新能力受到制约,因此,过高的森林郁闭度是限制琼岛杨自然更新、导致天然个体急剧减少的原因之一。
4 结论
适宜的光照条件对琼岛杨幼苗的生长发育至关重要,琼岛杨为强阳性树种,其幼苗对弱光的适应性较差,光照过弱不利于其生长发育,在相对光强70% 时,调节了植株表型特征和光合色素,有利于其生长、生物量累积和光合作用,促进其生长发育,说明琼岛杨幼苗在相对光强70% 下较其他光照强度具有更强的适应性。
参考文献
[1]顾雪丹,吕东,赵祜,等. 遮阴对红砂幼苗生长及光合特性的影响[J]. 干旱区资源与环境,2023,37(8):145-152.
Gu X D,Lv D,Zhao H,et al. Influence of shading on growthand photosynthetic characteristics of Reaumuria soongoricaseedlings[J]. Journal of Arid Land Resources and Environment,2023,37(8):145-152.
[2]Pearcy R W, Sims D A.Photosynthetic acclimation to changinglight environments:scaling from the leaf to the whole plant[M]//Exploitation of Environmental Heterogeneity by Plants.Amsterdam:Elsevier,1994:145-174.
[3]Deguchi R,Koyama K. Photosynthetic and morphologicalacclimation to high and low light environments in Petasitesjaponicus subsp. giganteus[J].Forests,2020,11(12):1365.
[4]张俊杰,刘青,韦霄,等. 光强对金丝李幼苗生长及光合特性的影响[J]. 林业科学,2022,58(5):53-64.
Zhang J J,Liu Q,Wei X,et al. Influence of light intensity ongrowth and photosynthetic characteristics of Garciniapaucinervis seedlings[J]. Scientia Silvae Sinicae,2022,58(5):53-64.
[5]李小琴,张凤良,杨湉,等. 遮阴对濒危植物风吹楠幼苗叶形态和光合参数的影响[J]. 植物生理学报,2019,55(1):80-90.
Li X Q,Zhang F L,Yang T,et al. Effect of shading on leafmorphology and photosynthetic parameters in endangeredHorsfieldia glabra seedlings[J]. Plant Physiology Journal,2019,55(1):80-90.
[6]杨霖,黄郑雯,毛开泽,等. 望天树幼苗光合特性与功能性状对光照和施肥的响应[J]. 西部林业科学,2021,50(6):46-52.
Yang L, Huang Z W, Mao K Z, et al. Response ofphotosynthetic characteristics and functional traits of Parashoreachinensis’ seedlings to light and fertilization[J]. Journal of WestChina Forestry Science,2021,50(6):46-52.
[7]徐清,闭鸿雁,崔光帅,等. 珍稀濒危植物毛果木莲幼苗光合特性及对遮阴处理的响应[J]. 南京林业大学学报(自然科学版),2019,43(6):46-52.
Xu Q,Bi H Y,Cui G S,et al. Response of photosyntheticfluorescence of the endangered plant Manglietia ventii seedlingsto shade treatment[J]. Journal of Nanjing Forestry University(Natural Sciences Edition),2019,43(6):46-52.
[8]冯玉龙,曹坤芳,冯志立,等. 四种热带雨林树种幼苗比叶重,光合特性和暗呼吸对生长光环境的适应[J]. 生态学报,2002,22(6):901-910.
Feng Y L,Cao K F,Feng Z L,et al. Acclimation of Laminamass per unit area,photosynthetic characteristics and darkrespiration to growth light regimes in four tropical rainforestspecies[J]. Acta Ecologica Sinica,2002,22(6):901-910.
[9]舒文将,柴胜丰,黎兆海,等. 不同光强对棱角山矾幼苗生长及光合特性的影响[J]. 植物研究,2017,37(4):556-562.
Shu W J,Chai S F,Li Z H,et al. Effects of different lightintensities on the growth and photosynthesis of Symplocostetragona seedlings[J]. Bulletin of Botanical Research,2017,37(4):556-562.
[10]Sefcik L T,Zak D R,Ellsworth D S. Photosynthetic responsesto understory shade and elevated carbon dioxide concentrationin four northern hardwood tree species[J]. Tree Physiology,2006,26(12):1589-1599.
[11]陈菊艳,龙海燕,邓伦秀. 遮光对野鸦椿幼苗生长和光合生理特性的影响[J]. 西部林业科学,2021,50(2):19-27.
Chen J Y,Long H Y,Deng L X. Effects of shading on growthand photosynthetic characteristics of Euscaphis japonicaseedlings[J]. Journal of West China Forestry Science,2021,50(2):19-27.
[12]王满莲,韦霄,唐辉,等. 光强对三种喀斯特植物幼苗生长和光合特性的影响[J]. 生态学杂志,2015,34(3):604-610.
Wang M L,Wei X,Tang H,et al. Effects of light intensity ongrowth and photosynthesis of three Karst plant seedlings[J].Chinese Journal of Ecology,2015,34(3):604-610.
[13]王琳,王满莲,梁惠凌,等.遮阴对地枫皮幼苗生长和生物量分配的影响[J/OL].热带亚热带植物学报,2022:1-9[. 2023-10-09].https://kns.cnki.net/kcms/detail/44.1374.Q.20221208.1656.001.html.
Wang L,Wang M L,Liang H L,et al. Effect of shading ongrowth and biomass allocation of Illicium difengpi seedlings[J/OL]. Journal of Tropical and Subtropical Botany, 2022:1-9.[2023-10-09]. https://kns. cnki. net/kcms/detail/44.1374.Q.20221208.1656.001.html.
[14]梁居红,方发之,陈彧,等. 琼岛杨野生资源的调查[J]. 热带林业,2012,40(1):32-34.
Liang J H,Fang F Z,Chen Y,et al. Survey on wild Populusqiongdaoensis T. Hong et P. Luo resource[J]. TropicalForestry,2012,40(1):32-34.
[15]梁居红,方发之,陈彧,等. 高山琼岛杨引种迁地培育技术研究[J]. 热带林业,2013,41(1):28-29,20.
Liang J H,Fang F Z,Chen Y,et al. The Qiong Dao YangPoplar introduction and ex situ cultivation technology research[J]. Tropical Forestry,2013,41(1):28-29,20.
[16]陈彧,黄小谦,徐建辉,等. 琼岛杨高枝压条繁殖技术研究[J].热带林业,2019,47(4):22-25.
Chen Y,Huang X Q,Xu J H,et al. Propagation of Populusqiongdaoensis by air layering[J]. Tropical Forestry, 2019,47(4):22-25.
[17]饶丹丹,刘雯雯,陈彧,等. 琼岛杨嫩枝扦插影响因素探究[J/OL]. 分子植物育种,2023:1-14.[2023-10-09]. https://kns.cnki.net/kcms/detail/46.1068.S.20230612.1144.002.html.
Rao D D,Liu W W,Chen Y,et al. Study on the factorsinfluencing twig cuttage of Populus qiongdaoensis[J/OL].Molecular Plant Breeding,2023:1-14.[2023-10-09]. https://kns.cnki.net/kcms/detail/46.1068.S.20230612.1144.002.html.
[18]陈彧,李小成,韩豫,等. 琼岛杨一年生播种苗生长节律研究[J]. 浙江林业科技,2021,41(1):88-92.
Chen Y,Li X C,Han Y,et al. Experiment on growth of 1-yearPopulus qiongdaoensis seedlings [J]. Journal of ZhejiangForestry Science and Technology,2021,41(1):88-92.
[19]周双清,梁居红. 琼岛杨愈伤组织培养的初步研究[J]. 热带林业,2014,42(2):15-17.
Zhou S Q,Liang J H. A preliminary study of Populusqiongdaoensis T. Hong et P. Luo callus culture[J]. TropicalForestry,2014,42(2):15-17.
[20]徐家洪,曾晴,叶富余,等. 基于全长转录组序列、核基因与叶绿体基因分析琼岛杨在杨属的亲缘关系[J]. 北京林业大学学报,2021,43(10):28-37.
Xu J H,Zeng Q,Ye F Y,et al. Genetic relationship of Populusqiongdaoensis in Populus based on full-length transcriptomesequences, nuclear genes and chloroplast genes[J]. Journal ofBeijing Forestry University,2021, 43(10):28-37.
[21]Xu J H,Fang M,Li Z H,et al. Third-generation sequencingreveals LncRNA-regulated HSP genes in the Populus xcanadensis moench heat stress response [J]. Frontiers inGenetics,2020,11:249.
[22]Xu J H,Du R Y,Meng X X,et al. Third-generationsequencing indicated that LncRNA could regulate eIF2D toenhance protein translation under heat stress in Populus simonii[J].Plant Molecular Biology Reporter,2021,39:240-250.
[23]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:134-137.
Li H S. Principles and techniques of plant physiological andbiochemical Experiment[M]. Beijing:Higher Education Press.2000:134-137.
[24]罗光宇,陈超,李月灵,等. 光照强度对濒危植物长序榆光合特性的影响[J]. 生态学杂志,2021,40(4):980-988.
Luo G Y,Chen C,Li Y L,et al. Effects of light intensity on thephotosynthetic characteristics of Ulmus elongata[J]. ChineseJournal of Ecology,2021,40(4):980-988.
[25]周欢,韦如萍,李吉跃,等.光照强度对乐昌含笑幼苗生长及光合特性的影响[J/OL].生态学杂志,2023:1-9[. 2023-10-09].https://kns.cnki.net/kcms/detail/21.1148.Q.20230328.1344.004.html.
Zhou H,Wei R P,Li J Y,et al. Effects of light intensity ongrowth and photosynthetic characteristics of Michelia chapensisDandy seedlings[J/OL]. Chinese Journal of Ecology,2023:1-9.[2023-10-09]. https://kns. cnki. net/kcms/detail/21.1148.Q.20230328.1344.004.html.
[26]李珍,谢世清,徐文果,等. 间作和净作条件下喜阴植物谢君魔芋的光合作用及光合诱导特征研究[J]. 热带亚热带植物学报,2017,25(1):26-34.
Li Z,Xie S Q,Xu W G,et al. Photosynthesis and photosyntheticinduction of shade-tolerant species Amorphophallus xiei underintercropping and monoculture planting patterns[J]. Journal ofTropical and Subtropical Botany,2017,25(1):26-34.
[27]张珊珊,袁春明. 土壤水分和光强对景东翅子树幼苗生长及光合特性的影响[J]. 植物研究,2022,42(3):502-511.
Zhang S S,Yuan C M. Effects of soil moisture and lightintensity on growth and photosynthetic characteristics ofPterospermum kingtungense seedlings[J]. Bulletin of BotanicalResearch,2022,42(3):502-511.
[28]韦中绵,覃德文,吴敏,等. 不同光照强度对火力楠幼苗生长及生理特性的影响[J]. 西部林业科学,2018,47(2):48-53.
Wei Z M,Qin D W,Wu M,et al. Growth and physiologicalcharacteristics of Michelia macclurei in different light intensities[J]. Journal of West China Forestry Science,2018,47(2):48-53.
[29]姬拉拉,刘筱,王锐洁,等. 遮阴和施氮对金荞麦生长和光合荧光特性的影响[J]. 山西农业大学学报(自然科学版),2019, 39(5): 23-31.
Ji L L,Liu X,Wang R J,et al. Effects of shading levels andnitrogen application on the growth and photosyntheticfiuorescence characteristics of Fagopyrum dibotrys[J]. Journalof Shanxi Agricultural University (Natural Science Edition),2019,39(5):23-31.
[30]安静,韦小丽,姚瑶,等. 闽楠幼苗适应弱光环境的可塑性研究[J/OL]. 广西植物,2023:1-11.[2023-10-09].https://kns.cnki.net/kcms/detail/45.1134.Q.20230524.1636.002.html.
An J,Wei X L,Yao Y,et al. Plasticity of Phoebe bourneiseedlings adapting to low light environment[J/OL]. Guihaia,2023:1-11.[2023-10-09]. https://kns. cnki. net/kcms/detail/45.1134.Q.20230524.1636.002.html.
[31]代大川,胡红玲,陈洪,等. 遮阴对桢楠幼苗生长和光合生理特性的影响[J]. 西北农林科技大学学报(自然科学版),2020,48(4):56-64,74.
Dai D C,Hu H L,Chen H,et al. Effects of shading on growthand photosynthetic characteristics of Phoebe zhennan seedlings[J]. Journal of Northwest Aamp;F University (Natural ScienceEdition),2020,48(4):56-64,74.
(编辑:郭玥微)
猜你喜欢
今日农业(2021年10期)2021-11-27 09:45:24
中学生物学(2017年2期)2017-03-20 07:30:12
现代农业科技(2017年1期)2017-03-06 12:38:31
山东农业科学(2016年12期)2017-01-21 14:41:54
山东农业科学(2016年12期)2017-01-21 14:39:43
山东农业科学(2016年11期)2016-12-17 21:16:16
吉林农业·下半月(2016年10期)2016-12-16 23:45:53
农业与技术(2016年20期)2016-12-08 21:57:43
湖南师范大学学报·自然科学版(2016年3期)2016-06-25 06:22:53
中国火炬(2015年12期)2015-07-31 17:38:35
