
矩镰荚苜蓿地下器官贮藏组织中淀粉粒变化规律
2024-04-24 09:25:10李永强杨小霞张京萍杨艳丽方强恩
草业科学 2024年3期
李永强,杨小霞,张京萍,杨艳丽,方强恩
(甘肃农业大学草业学院 / 草业生态系统教育部重点实验室 / 甘肃省草业工程实验室 /中-美草地畜牧业可持续发展研究中心, 甘肃 兰州 730070)
淀粉是植物体内具有储存功能的糖类物质,在寒冷胁迫下,植物体内水解酶活性大幅提升,促使大量的淀粉加速分解成可溶性糖[1]。可溶性糖作为渗透保护物质,可以通过提高细胞液的浓度、减缓水分散失和降低细胞质的冰点等方式保护植物器官[2]。因此,淀粉不仅为植物贮能,也是植物应对寒冷逆境的调节物质。赵凤军和张艳红[3]对多种杜鹃花属(Rhododendron)植物研究发现,细胞内淀粉粒在越冬休眠前积累丰富,越冬休眠期充分水解,休眠解除时又会重新积累。董丽等[4]对多种常绿植物越冬特性的研究也同样发现,在秋季植物细胞中都有较多的淀粉粒分布,冬季观测时淀粉粒消失,到了春季又重新开始积累。大量研究数据均表明,储藏组织中淀粉粒的季节性动态变化与植物抗寒能力及越冬适应节律具有高度一致性[5-6]。这为从组织化学角度研究植物越冬抗寒机制提供了依据。
苜蓿(Medicago)是奶牛等草食动物的重要优质饲草。发展苜蓿产业,对于调整种植业结构、提升草产品和畜产品市场竞争力具有重要意义。西北是我国苜蓿的主产区,但是西北大部分地区,尤其是青藏高原区,因海拔高、气温低等因素导致苜蓿产量降低,越冬率下降,严重影响了苜蓿种植的生态和经济效益[7-8]。因此,苜蓿的安全越冬问题仍然是目前制约我国北方苜蓿草地建植、生产与可持续利用的关键问题[9]。研究高寒区野生苜蓿属植物抗寒机理,挖掘适应高寒气候条件的优异种质,对苜蓿抗寒品种选育和高寒区栽培技术研究具有重要意义。
矩镰荚苜蓿(M.archiducis-nicolai)是豆科苜蓿属多年生野生牧草,别名青海苜蓿、青藏扁蓿豆等[10],广泛分布在我国青海、甘肃、四川、西藏等海拔2 500~3 900 m 的高寒地区[11]。矩镰荚苜蓿营养价值高,在青藏高原高寒气候条件下自然生长,能在极端低温条件下安全越冬返青[12],具有极强的抗寒能力。目前,关于矩镰荚苜蓿抗寒能力的相关研究较少,其抗寒机理至今还没有完全揭示。基于此,本研究通过观测矩镰荚苜蓿地下器官越冬期间的解剖结构与淀粉变化特征,探究其越冬期抗寒能力的变化规律,旨在进一步开展矩镰荚苜蓿越冬抗寒机理研究提供参考。
1 材料与方法
1.1 试验地概况
试验地位于青藏高原东北缘的武威市天祝藏族自治县抓喜秀龙镇(102°79′11″ E, 37°19′67″ N),平均海拔高度3 022 m, 属于温带大陆性高原季风气候区。试验地年平均气温-0.1 ℃,≥ 0 ℃年积温1 300℃·d,年日照时间2 500 h,雨季集中在7 月-9 月,年平均降水量400 mm,年平均蒸发量1 590 mm。早霜9 月下旬,晚霜4 月下旬,无霜期155 d。土壤主要为高山黑钙土,土壤pH 7.0~8.2,有机质含量10%~16%,土层厚度40~80 cm[13]。
1.2 试验材料
材料为甘肃农业大学天祝高山草原生态试验站内自然生长的矩镰荚苜蓿。根据采样地的气温变化(图1)及土壤封冻状况将矩镰荚苜蓿的越冬过程划分为3 个时期(表1),分别为越冬前期(9 月-11 月)、越冬中期(11 月-3 月)和越冬后期(3 月-5 月)。
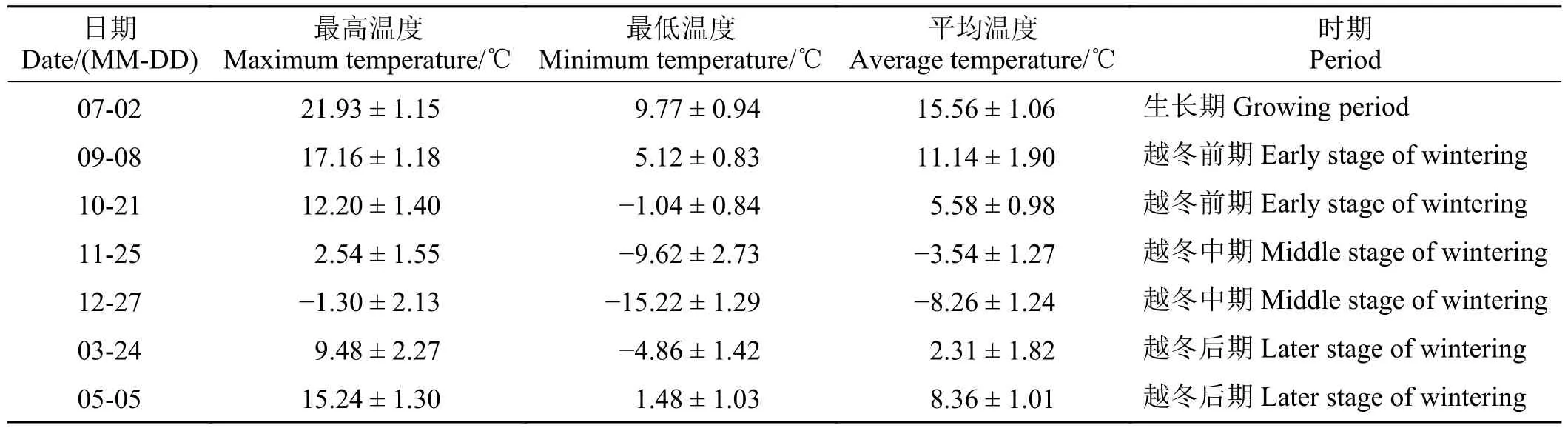
表1 不同取样时期气温Table 1 Ambient temperatures during the different sampling periods

图1 试验地气温变化Figure 1 Changes in ambient temperature at the test site
低温变化是影响植物休眠态转变的关键,因此,在矩镰荚苜蓿越冬前期设置2 次取样(分别为9 月8 日:日均温度10 ℃以上取样一次;10 月21 日:温度0~5 ℃,植物受到冷胁迫时取样一次)、越冬中期设置2 次取样(分别为11 月25 日:样地日均温度降低至0 ℃以下,土壤开始封冻时取样一次;12 月27 日:最低气温降到-10 ℃以下,植株受到冻胁迫时取样一次)、越冬后期设置2 次取样(3 月24 日气温回升到0 ℃以上时取样一次;5 月5 日日均温度5 ℃以上,植物开始返青时取样一次)。对试验地与材料进行调查与鉴定后,在矩镰荚苜蓿株丛随机选取生长状况良好的植株,以单株矩镰荚苜蓿为中心,在50 cm × 50 cm 的范围内,由外向内逐渐剖开、向下直到矩镰荚苜蓿地下器官分布的最底层,再向一侧跟踪性挖取地下器官。重复此操作,挖取10 株完整的矩镰荚苜蓿。另外,在矩镰荚苜蓿生长期(7 月2 日)取样一次作为对照。
1.3 试验方法及分析
参考杨小霞等[14]的试验方法,将挖取的材料用蒸馏水冲洗干净,用滤纸吸干地下部分水分后,分开地下器官的根、根颈和地下茎,分别切取5 mm 左右长的段各10 段放入FAA 溶液(70%乙醇溶液 ∶ 冰乙酸 ∶ 福尔马林 = 18 ∶ 1 ∶ 1)中固定,抽真空使试验材料下沉瓶底,4 ℃下保存备用。
将材料固定48 h 后,采用常规的石蜡制片法制片。经乙醇梯度脱水、透明、浸蜡、包埋等过程后,使用自动切片机进行切片,厚度范围8~12 μm,粘片后,恒温37 ℃烘片36 h。使用番红-固绿法对矩镰荚苜蓿地下器官的不同部分组织染色,使用高碘酸-席夫反应法(PAS 法)对地下器官组织淀粉粒进行染色,淀粉颗粒在碱性品红染色下呈紫红色,最后使用中性树胶封片。在Motic Panthera U 生物显微镜下进行观察,辨别矩镰荚苜蓿的地下器官淀粉储存部位,每个部位随机选取5 个视野,每个视野选取3 个细胞,统计其淀粉颗粒数量,记录并拍照。利用SPSS 26.0 对统计数据计算处理,Origin 2018 制图。
2 结果与分析
2.1 矩镰荚苜蓿地下器官的解剖结构比较
矩镰荚苜蓿越冬根的解剖结构(图2A)显示,根由外向内主要由周皮和维管柱构成。其中,周皮较厚,木栓层极发达。栓内层由6~7 层薄壁细胞构成,细胞较薄壁细胞大且密集。木质部占据维管柱大部分,导管极多,木质部极发达。导管排列紧密,中部导管直径较大,木射线明显。
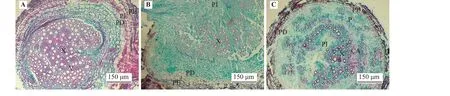
图2 矩镰荚苜蓿根、根颈和地下茎的解剖结构Figure 2 Anatomical structure of the root, crown, and rhizome tissues of Medicago archiducis-nicolai
矩镰荚苜蓿越冬根颈解剖结构(图2B)在组成上与根相似,由周皮和维管柱组成,但根颈在维管柱中央具有完整的髓。与根周皮相比,根颈周皮外部的木栓层更加发达,且具有栓内层薄壁组织。栓内层位于周皮和维管柱之间,由多层体积较大的薄壁细胞组成,整体较厚,细胞排列疏松,有明显的细胞间隙。根颈维管柱内木质部导管排列较规则,呈辐射状向外延伸,导管较小,且数量少于根木质部。木质部薄壁细胞分布在木质部导管之间,数量较多。
矩镰荚苜蓿越冬地下茎的解剖结构(图2C)与根颈相同。周皮较厚,木栓层细胞呈砖形,排列紧密。栓内层由3~4 层薄壁细胞构成,细胞排列稀疏,有胞间隙。与根颈不同的是,地下茎木质部导管散乱分布,导管大小极不规则,维管柱内髓狭长,有裂隙。
2.2 矩镰荚苜蓿越冬时期根组织结构中的淀粉粒变化
矩镰荚苜蓿根组织中淀粉粒主要分布在栓内层与木质部薄壁细胞中,淀粉粒数均在9 月上旬积累至最高,其中栓内层细胞淀粉粒数较多,且对低温变化较敏感(图3)。11 月下旬,在越冬期开始时,根栓内层淀粉数骤降至较低水平,并在12 月下旬降至最低(图3)。木质部薄壁细胞淀粉粒数也在越冬期开始时下降,越冬中期降至最低(图3)。越冬后期,根淀粉粒又逐渐开始积累(图3)。不同于根颈与地下茎的是(图3),根木质部薄壁细胞可以积累更多的淀粉粒数,且对温度的变化更为敏感(图3)。此外,矩镰荚苜蓿地下器官细胞内淀粉多为4~8 粒淀粉颗粒聚集在一起,整体集中呈团状(图4、图5、图6)。切片显示(图4),根越冬前期细胞内淀粉粒较大,分布密集,积累成较大的团状。越冬中期前后,淀粉粒较小,分布散乱。

图3 矩镰荚苜蓿根、根颈和地下茎组织结构不同部位细胞淀粉数量的变化Figure 3 Changes in the number of cell starches in different parts of Medicago archiducis-nicolai root, crown, and rhizome tissue structures
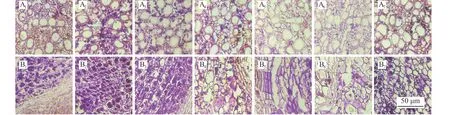
图4 矩镰荚苜蓿根组织淀粉积累变化Figure 4 Changes in starch accumulation in Medicago archiducis-nicolai root tissues

图5 矩镰荚苜蓿根颈组织淀粉积累变化Figure 5 Changes in starch accumulation in Medicago archiducis-nicolai crown tissues
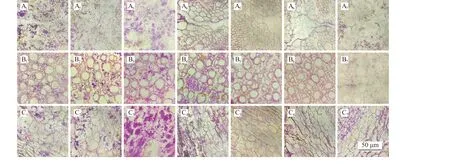
图6 矩镰荚苜蓿地下茎组织淀粉积累变化Figure 6 Changes in starch accumulation in Medicago archiducis-nicolai rhizome tissues
2.3 矩镰荚苜蓿越冬时期根颈组织结构中的淀粉粒变化
矩镰荚苜蓿根颈组织内淀粉主要分布于髓与栓内层薄壁细胞中,少量分布于木质部薄壁细胞中(图3)。越冬前期,髓与栓内层的淀粉粒积累量逐渐下降,在越冬中期达到最低水平。其中,栓内层淀粉粒数量下降较快,在12 月下旬,先于髓、木质部到达最低点;相比于髓细胞,栓内层对低温的变化更敏感。木薄壁细胞在越冬前期与越冬中期开始时,对温度变化不敏感,淀粉粒数量无明显变化。在3 月下旬,越冬中期结束时,淀粉粒数下降至最低水平。根颈髓、木质部、栓内层均在越冬后期开始积累淀粉粒,淀粉粒数在5 月上旬显著增加。切片显示(图5),各地下器官在生长期、越冬前期及越冬后期结束时,均贮存有较大的淀粉颗粒,且分布密集。越冬后期开始时,淀粉粒数量总体最少,淀粉颗粒最小。
2.4 矩镰荚苜蓿越冬时期地下茎组织结构中的淀粉粒变化
矩镰荚苜蓿地下茎内淀粉粒主要分布在栓内层与髓薄壁细胞中,部分分布于木质部薄壁细胞中,且其数量在生长期维持在较低水平(图3)。地下茎的淀粉在越冬前期持续积累,越冬前期开始时,淀粉数量明显提升,10 月下旬达到最高。越冬中期开始时,淀粉粒数骤降至最低点,淀粉几乎消失,在整个越冬中期的较长时间段内,保持在最低水平,持续至越冬期完全结束,直到越冬后期,开始少量积累淀粉。与根颈在越冬前期各组织淀粉数量较多,积累量大,数量随温度变化而变化的状况不同,地下茎在气温未达到冰点的越冬前期持续积累淀粉,越冬中期开始之前达到最高。切片显示(图6),在越冬前期,矩镰荚苜蓿地下茎内淀粉粒最大,生长期淀粉粒相对较小,越冬后期细胞内淀粉粒最小,越冬中期几乎无明显淀粉粒。
3 讨论
3.1 矩镰荚苜蓿根颈和地下茎的解剖结构与功能
目前,关于苜蓿根颈的定义及其属性的说法较多且不一致[15]。通常认为,苜蓿近地面上的根冠部称为根颈,由真叶以下的胚轴发育而来[16]。张宝田等[17]认为紫花苜蓿根颈是联系地上部分和根系的关键部位,可能属于根与茎的过渡区。杜宝红[18]研究了花苜蓿(M.ruthenica)根颈形态解剖特征,认为花苜蓿的根颈就是根的最上端部分。本研究通过解剖观察发现,矩镰荚苜蓿根颈组织中,内部具有发达的髓,外部具有周皮,与根结构不完全相同,应该是变态的地下茎。通常情况下,植物淀粉粒主要贮存于髓、皮层等薄壁组织中[19]。发达的髓组织使根颈产生了重要的养分贮备能力。
本研究中,矩镰荚苜蓿具有发达的地下茎,其解剖结构组成与根颈基本相同。前期研究发现,地下茎是矩镰荚苜蓿不同于其近缘种的重要的营养繁殖器官,具有极强的地下扩展能力[20]。为了应对寒冷和干旱环境,矩镰荚苜蓿地下茎可以产生内、外周皮,在极端寒旱环境下,地下茎会通过外周皮内卷或内周皮外拓发生分裂[21]。在本研究中,矩镰荚苜蓿地下茎和根颈相似,与根相比除了具有栓内层薄壁组织外,还具有发达的髓(图2C)。在越冬过程,髓与栓内层薄壁组织会贮存大量淀粉(图5、图6),这表明地下茎是矩镰荚苜蓿兼具扩展、繁殖、贮藏等功能的特殊器官。
3.2 矩镰荚苜蓿越冬时期地下器官淀粉变化与抗寒性分析
低温环境下,植物组织中的可溶性糖含量升高是一个普遍现象[22]。越冬前后植物细胞内淀粉数量会随温度变化而变化[23],淀粉的水解不仅可以为地下器官提供能量和营养,且有利于可溶性糖、蛋白质等抗寒相关物质的合成,提高植物的抗寒能力[24]。可溶性糖与淀粉互相转化,二者存在动态平衡[25]。本研究结果也表现出相同的规律。在本研究中,矩镰荚苜蓿根、根颈与地下茎在越冬前后的淀粉数均呈现出先减少后增加变化趋势,其中地下茎淀粉数在越冬前期仍有一定增加,但各器官淀粉数量在同一时期始终维持较为相近的水平,与温度变化趋势基本一致,且不同组织部位淀粉数的变化趋势也基本一致。这表明矩镰荚苜蓿在寒冷环境中可以通过转化地下器官各组织中的淀粉来提高自身的抗寒能力,从而安全越冬。
北温带多年生植物在冬季越冬过程中通常会休眠,这是植物为抵抗寒冷等逆境而产生的一种生理现象[26]。前人研究发现,植物组织中淀粉的变化与休眠状态的发生有关[3]。植物在秋季感应到温度降低、日照时长变短信号后,会停止地上部分的生长,进入休眠准备阶段。对草本植物而言,此时会将地上养分向地下转移,以淀粉的形式储存在地下器官中,为其越冬提供能量储备[27-28]。随着休眠状态的不断加深,储藏器官中的大量淀粉会发生水解,植物的越冬抗寒能力不断增强,当植物进入深度休眠后,淀粉贮存量也达到最低,大部分淀粉分解为可溶性糖,以积极应对严冬下的寒冷胁迫[24]。从休眠态变化的角度分析本研究结果,发现矩镰荚苜蓿地下器官根、根颈、地下茎在越冬期都会发生休眠,但是不同器官进入深度休眠的时间存在差别(图3)。地下茎在最低气温降至0 ℃时(图1)淀粉分解才开始启动,但在之后短短的一个月内淀粉便全部分解,在严冬到来之前很快进入深度休眠,而且相比较于根和根颈,地下茎深度休眠持续时间较长,休眠解除非常缓慢。根和根颈薄壁组织内储藏的淀粉都是在严冬期才全部分解完,表明二者在冬季温度最低时进入深度休眠,不同的是,根颈早在7 月之后气温下降之初便开始分解淀粉,而根在最低气温降至5 ℃时(图1、图3)淀粉才开始分解。三者深度休眠起始时间的差异表明,地下茎对冬季低温胁迫最敏感,抗寒力获得时间最早且最持久。地下茎不仅是矩镰荚苜蓿重要的养分储藏器官,地下茎中产生的数量丰富的根茎芽是越冬后再生苗的主要来源[14],所以地下茎持久的抗寒力是矩镰荚苜蓿成功越冬和翌年返青更新生长的重要保障。此外,由于青藏高原海拔高,气候严酷多变,昼夜温差大,矩镰荚苜蓿地下器官抗寒力形成的不同步性,可能也是该植物应对高寒环境的适应性策略。
4 结论
矩镰荚苜蓿地下器官的淀粉主要分布在栓内层与髓薄壁细胞中,根颈与根内淀粉粒分布较为密集;越冬前期,淀粉粒在各地下器官均有明显的积累,低温变化下,淀粉发生水解,逐渐减少至消失。气温回升后,淀粉数量开始增加。各地下器官中的淀粉数量在越冬前后的变化与气温变化趋势基本一致,总体呈现出先减少后增加的变化规律;矩镰荚苜蓿地下器官越冬期抗寒力的形成具有不同步性,且根颈对温度变化响应最快,地下茎对低温变化最为敏感;矩镰荚苜蓿地下茎是兼具扩展、繁殖、贮藏与抗寒等能力的特殊器官。
猜你喜欢
建筑与预算(2023年12期)2024-01-18 06:45:50
建筑与预算(2023年11期)2023-12-25 12:38:44
建筑与预算(2023年9期)2023-10-21 10:14:20
建筑与预算(2023年2期)2023-03-10 13:13:20
西北农林科技大学学报(自然科学版)(2023年1期)2023-02-20 03:57:48
今日农业(2021年20期)2021-11-26 01:23:56
草地学报(2021年10期)2021-11-09 02:00:46
河北果树(2020年4期)2020-11-26 06:04:28
草地学报(2018年1期)2018-07-30 03:29:08
草原与草坪(2016年5期)2016-11-28 07:57:43
