
模拟氮沉降对刺萼龙葵幼苗生长与生物量的影响
2024-04-11 02:48王雪丽王鹤松王竹天周立业
内蒙古民族大学学报(自然科学版) 2024年2期
张 佳,王雪丽,王鹤松,王 鹏,王竹天,周立业
(1.内蒙古民族大学草业学院,内蒙古 通辽 028043;2.通辽市科尔沁区草原工作站,内蒙古 通辽 028000)
氮素是植物生长发育不可或缺的营养元素之一,调节植物的各个生命活动过程,如促进植物营养生长,提高光合能力,保证其正常生长发育。氮素水平的高低也将直接影响土壤肥力水平和土壤结构的好坏[1-2]。自工业革命以来,燃烧化石燃料、使用化学氮肥等人类活动使含氮化合物的排放量大幅度增加,从而导致全球氮沉降明显增加[3]。氮沉降可导致土壤酸化,引起生态系统的退化,甚至促进和加速某些物种对新生境的入侵[4]。大气氮沉降主要包括湿沉降和干沉降[5],沉降中的氮主要是铵态氮、硝态氮和有机氮等3种形态。张颖等[6]研究结果表明,在农田生态系统中以铵态氮的沉降为主,而城市生态系统中以硝态氮的沉降为主。相关研究表明,与本地植物相比,部分入侵植物具有较强的氮吸收速率,常表现为较快的生长速度和较强的适应能力,入侵性常随氮素含量的升高而增强[7]。氮沉降导致的营养资源水平的改变可能对本地种和入侵种造成不同的影响。尤其是氮沉降可提升入侵植物的资源利用率,使其具有更高的生存竞争优势,并通过抑制本地植物的生长,进而降低本地植物群落的生物多样性[8]。因此,氮沉降加剧可能对植物入侵具有促进作用。
茄科茄属一年生草本植物刺萼龙葵(Solanum rostratumDunal.)是入侵性非常强的一种植物,原产于北美洲,具有极耐干旱特性,有极快的蔓延传播速度,竞争能力强,能耐比较恶劣的生存环境,常形成大面积单一群落,可严重抑制其他植物生长,破坏当地生物多样性[9-10]。刺萼龙葵的入侵极大地危害了当地自然环境和生态系统。目前,氮沉降对刺萼龙葵的入侵机制影响研究鲜见报道。因此,以科尔沁沙地常见的恶性入侵植物刺萼龙葵为对象,通过氮添加试验模拟氮沉降,研究氮沉降对刺萼龙葵幼苗生长的影响和氮沉降对刺萼龙葵生物量分配格局的影响。
1 材料与方法
1.1 试验地概况
试验地位于内蒙古通辽市科尔沁区丰田镇内蒙古民族大学科技园区(43°64′N,122°07′E),为温带半干旱大陆性季风气候,温度适中,年均气温6.1 ℃,年均降水量385.1 mm,降水多集中于7—9月,土壤类型多为风沙土。试验用盆栽土壤基本养分情况如下:有机质7.28 mg/kg,全氮素1.87 g/kg,碱解氮素11.24 mg/kg,速效钾95.12 mg/kg,速效磷10.59 mg/kg。
1.2 试验设计
试验于2023年5月8日开始,根据前人研究结果[11-12],设置2种氮沉降方式(铵态氮((NH4)2SO4),记为A;硝态氮(NaNO3),记为B)和4 种氮沉降水平(0、4、8、10 g/(m2·a),记为1、2、3、4)共16 个处理,即A1B1(CK)、A1B2、A1B3、A1B4、A2B1、A2B2、A2B3、A2B4、A3B1、A3B2、A3B3、A3B4、A4B1、A4B2、A4B3和A4B4,依次记为处理1~16。每种氮沉降水平重复3次,共计48盆,随机放置。植物种植前,将刺萼龙葵种子用蒸馏水浸泡12 h,土壤过2 mm筛后混匀,每个花盆(上口直径29.5 cm,高度20 cm)装入等量的试验土壤并浇足水分。刺萼龙葵每盆播种20粒种子,覆土2 cm并压实,出苗后每盆定植10株。株高2 cm左右时开始施氮,幼苗期分2次施入,处理时将氮肥溶于水中施入土壤,对照施入等量的水,正常浇水管理。2023年6月9日取样测定各生长指标。
1.3 测定指标与方法
试验结束后,将所有植株整株挖出用水冲洗干净并除去根部土壤,用滤纸和吸水纸将植株表面水渍擦干。叶面积采用YMJ-CH智能叶面积扫描仪进行测定。根粗、茎粗采用电子游标卡尺测量,植株鲜质量采用电子天平进行测量,测量后用剪刀将根、茎、叶3部分分开,每个处理取5株分别称取根、茎、叶鲜质量,随后将植物的根、茎、叶分别装于牛皮纸袋放入烘箱中,105 ℃杀青15 min后75 ℃烘干至恒质量,再分别称量各部分的干质量。
1.4 相关指标计算方法
比叶面积=叶面积/叶干质量;
总生物量=所有生物量之和;
根生物量比=根生物量/总生物量;
茎生物量比=茎生物量/总生物量;
叶生物量比=叶生物量/总生物量;
根冠比=地下生物量/地上生物量。
1.5 数据分析
使用Microsoft Excel 2019 软件整理数据及作图。使用SPSS 26.0 软件进行数据统计分析,应用双因素方差分析(Two-way ANOVA)检验各生长指标在氮沉降处理下的差异显著性(P<0.05)。
2 结果与分析
2.1 模拟氮沉降对刺萼龙葵幼苗生长的影响
刺萼龙葵幼苗株高在处理12水平下达到最大值,为4.92 cm,在处理1水平达到最小值,为4.15 cm;铵态氮浓度为8和10 g/(m2·a)时各水平间的株高随硝态氮的浓度增大而升高。处理1(即CK)水平下的根粗与其他水平均有显著差异(P<0.05),且值最大,为1.59 cm,铵态氮浓度为4和10 g/(m2·a)时各水平间的根粗逐渐增大,处理5、处理6 与处理8 水平的根粗差异显著(P<0.05),处理16 的根粗显著大于处理13、处理14 和处理15(P<0.05);铵态氮、硝态氮及其交互处理对根粗均产生极显著影响(P<0.01)。铵态氮浓度为4 g/(m2·a)时各水平间的茎粗随硝态氮浓度增大而变粗,处理8与处理5之间存在显著差异(P<0.05),处理1水平值最大,为2.20 cm;铵态氮、铵态氮与硝态氮交互对茎粗的影响均存在极显著差异(P<0.01)。处理8和处理10与其他14个水平的比叶面积均存在显著差异(P<0.05),且在处理10水平达到最大值,为23 748.41 mm2/g,处理1水平的比叶面积最小,为2 505.35 mm2/g,所有氮处理对刺萼龙葵幼苗的比叶面积均产生极显著差异(P<0.01)。见表1、表2。

表1 模拟氮沉降对刺萼龙葵幼苗生长的影响Tab.1 Effect of simulated nitrogen deposition on growth of Solanum rostratum Dunal.seedlings

表2 铵态氮、硝态氮及其交互作用对刺萼龙葵幼苗形态指标影响的方差分析Tab.2 Analysis of variance of ammonium nitrogen,nitrate nitrogen and their interactions on morphological indices of Solanum rostratum Dunal.seedlings
2.2 模拟氮沉降对刺萼龙葵幼苗生物量积累的影响
除处理9和处理15外,处理8的根生物量与其他各水平均存在显著差异(P<0.05);在处理14水平下根生物量最少,为0.01 g,处理8水平下根生物量最多,为0.06 g,处理8的根生物量是处理14的6倍。除处理1、处理2、处理4和处理15外,处理9与其他11个处理的茎生物量均有显著差异(P<0.05),且值最大,为0.09 g,处理13的茎生物量最少,为0.02 g,处理9的茎生物量是处理13的4.5倍。除处理2、处理4、处理5、处理8和处理9外,处理15与其他水平的叶生物量间均有显著差异(P<0.05),且值最大,为0.16 g,处理9和处理1、6、7、12、13、14、16间的叶生物量存在显著差异(P<0.05)。除处理8外,处理9和处理15的总生物量与其他各水平均存在显著差异(P<0.05)。见图1~图4。

图1 模拟氮沉降下刺萼龙葵幼苗的根生物量Fig.1 Root biomass of Solanum rostratum Dunal.seedlings under simulated nitrogen deposition
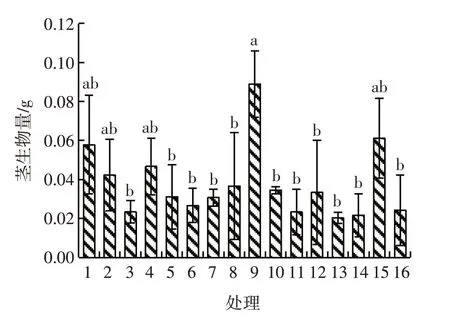
图2 模拟氮沉降下刺萼龙葵幼苗的茎生物量Fig.2 Stem biomass of Solanum rostratum Dunal.seedlings under simulated nitrogen deposition
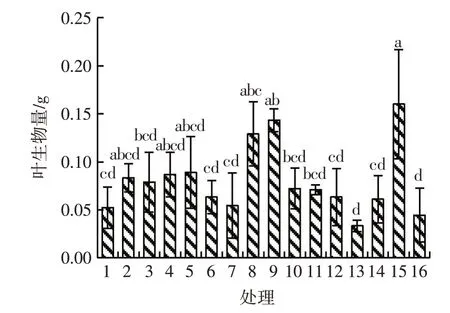
图3 模拟氮沉降下刺萼龙葵幼苗的叶生物量Fig.3 Leaf biomass of Solanum rostratum Dunal.seedlings under simulated nitrogen deposition
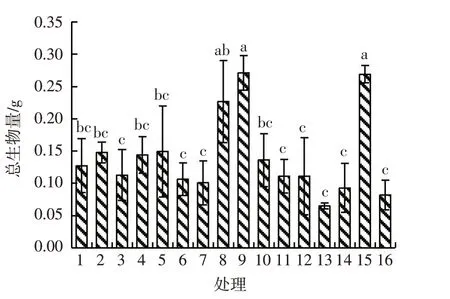
图4 模拟氮沉降下刺萼龙葵幼苗的总生物量Fig.4 Total biomass of Solanum rostratum Dunal.seedlings under simulated nitrogen deposition
铵态氮、铵态氮与硝态氮交互对刺萼龙葵幼苗根生物量的影响均存在极显著差异(P<0.01)。铵态氮和硝态氮交互对刺萼龙葵幼苗茎生物量的影响有极显著差异(P<0.01)。铵态氮、铵态氮与硝态氮交互对刺萼龙葵幼苗叶生物量的影响均产生极显著差异(P<0.01)。硝态氮和铵态氮交互对总生物量的影响存在极显著差异(P<0.01)。见表3。
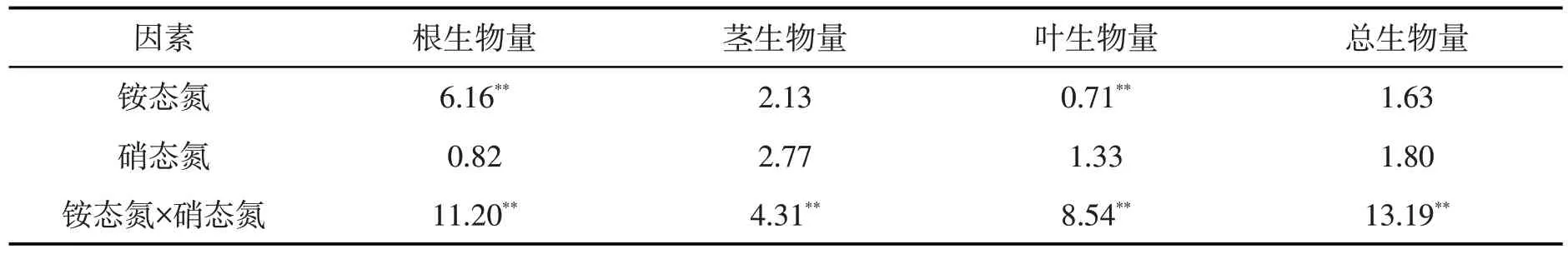
表3 铵态氮、硝态氮及其交互作用对刺萼龙葵幼苗生物量积累影响的方差分析Tab.3 Analysis of variance of ammonium nitrogen,nitrate nitrogen and their interactions on biomass accumulation of Solanum rostratum Dunal.seedlings
2.3 模拟氮沉降对刺萼龙葵幼苗生物量分配的影响
处理8中刺萼龙葵幼苗的根生物量比最高,为30.00%,处理4的最低,为8.50%;处理8的根生物量比是处理4的3.5倍。刺萼龙葵幼苗的茎生物量比只有处理9高于处理1,其他处理都降低了刺萼龙葵幼苗的茎生物量比,且处理3最低,为21.00%。除处理9外,其他处理均增大了刺萼龙葵幼苗的叶生物量比。见图5。除处理12外,处理8的刺萼龙葵幼苗根冠比显著高于其他处理(P<0.05),为1.84%;处理16的根冠比最小,为0.17%。见图6。
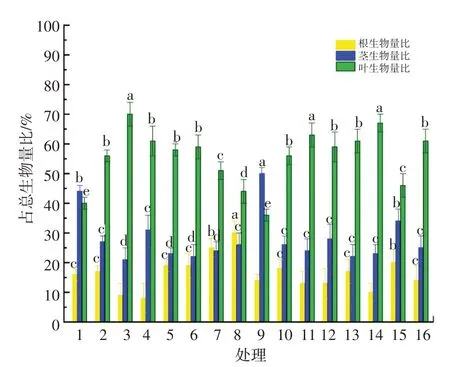
图5 模拟氮沉降下刺萼龙葵幼苗根茎叶占总生物量比重Fig.5 The proportion of rhizomes and leaves in total biomass of Solanum rostratum Dunal.seedlings under stimulated nitrogen deposition

图6 模拟氮沉降下刺萼龙葵幼苗根冠比Fig.6 Root-shoot ratio of Solanum rostratum Dunal.seedlings under stimulated nitrogen deposition
铵态氮对刺萼龙葵幼苗根生物量比有显著影响(P<0.05),铵态氮与硝态氮交互对刺萼龙葵幼苗根生物量比的影响存在极显著差异(P<0.01)。硝态氮、铵态氮与硝态氮交互对刺萼龙葵幼苗茎生物量比的影响均存在显著差异(P<0.05)。铵态氮与硝态氮交互对刺萼龙葵幼苗叶生物量比的影响存在极显著差异(P<0.01)。铵态氮和硝态氮对刺萼龙葵幼苗根冠比的影响显著(P<0.05),铵态氮与硝态氮交互对刺萼龙葵幼苗根冠比的影响存在极显著差异(P<0.01)。见表4。
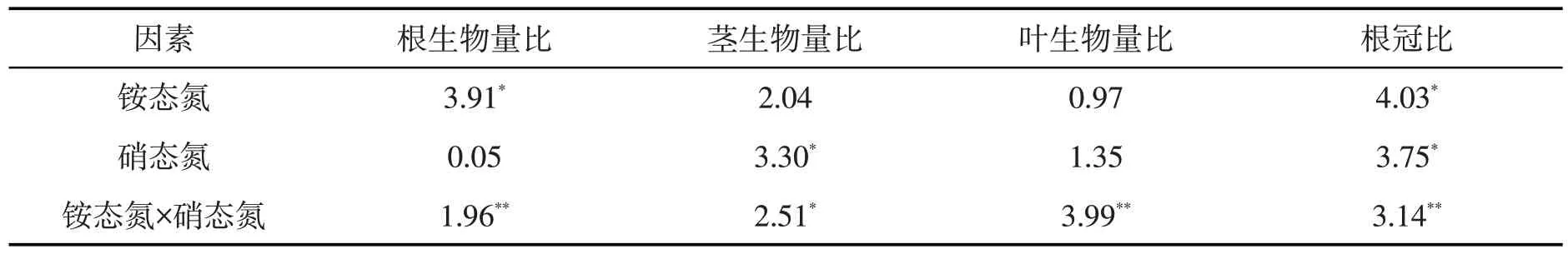
表4 铵态氮、硝态氮及其交互作用对刺萼龙葵幼苗生物量分配影响的方差分析Tab.4 Analysis of variance of ammonium nitrogen,nitrate nitrogen and their interactions on biomass allocation of Solanum rostratum Dunal.seedlings
3 讨论
3.1 模拟氮沉降对刺萼龙葵幼苗生长特性的影响
植物株高和叶面积是衡量植物生长的重要指标,是植物获取光合作用空间和更多光照的保证[13]。本研究结果表明,所有氮沉降处理能不同程度地增大刺萼龙葵幼苗的株高,同时,所有氮沉降处理均显著增大了刺萼龙葵幼苗的比叶面积,可能的原因是刺萼龙葵为适应氮沉降影响,增加其植株株高和比叶面积水平以提高光合速率,影响其生物量合成[14]。在环境较好时增加比叶面积有利于提高获取资源的能力,使其能充分利用环境资源,加快植株的生长发育,进而增强自身的入侵竞争能力[15-16]。本试验结果显示,氮沉降处理对刺萼龙葵幼苗的根粗和茎粗影响效果不明显,甚至降低了根粗和茎粗的大小,这说明氮沉降在一定程度上减少干物质向根部分配的比例,可能的原因是刺萼龙葵幼苗将更多的资源分配到地上部分,施氮更有利于促进植物光合器官的发育,又在一定程度上促进了地上部分的生长,以获得更好的空间资源和光照资源,有利于在地上的竞争中获得优势[17-18]。
3.2 模拟氮沉降对刺萼龙葵幼苗生物量积累的影响
在不同氮沉降水平下,生物量的积累很大程度上反映了植物的生长发育情况[19]。本研究结果显示,无氮处理的刺萼龙葵幼苗茎生物量的积累大于叶生物量的积累,而其他氮沉降水平幼苗的叶生物量的积累多于茎生物量的积累,这说明氮沉降改变了刺萼龙葵幼苗各器官生物量的积累,把更多营养输送到叶片上,促进了叶生物量的积累。同时,本研究结果显示,氮沉降处理下刺萼龙葵幼苗的叶生物量和茎生物量均大于根生物量的积累,可能的原因是氮沉降使刺萼龙葵减少了地下生物量的分配,根系仅需少量的能源分配便能满足其生长发育所需的水分和营养条件[20],将更多的资源分配到地上部分,叶生物量的积累使更多的生物量投入到碳同化器官,促进叶片的养分吸收能力,从而提高刺萼龙葵的竞争能力[21]。
3.3 模拟氮沉降对刺萼龙葵幼苗生物量分配的影响
植物生物量积累与分配特征能反映出植物对环境的适应性,构件不同,对氮的响应不同[22]。在本研究中,不同氮沉降水平下刺萼龙葵幼苗生物量的分配表现为地上生物量比大于地下生物量比,可能的原因是刺萼龙葵幼苗在养分丰富的环境中更倾向于投资地上部分,增加茎生物量以便支持、固着植株,并促进分枝,形成更多的叶片,而叶片是植物利用氮最为集中的器官,植物通过根系吸收的氮常常大量转移到绿色叶片以固定相对较多的碳[23],以增强其竞争力,获取和利用更多资源。本研究结果显示,未进行氮沉降处理的刺萼龙葵幼苗生物量分配表现为茎生物量比大于叶生物量比,而不同氮沉降处理下刺萼龙葵幼苗的生物量分配表现为叶生物量比大于茎生物量比,这说明不同水平的氮沉降对刺萼龙葵幼苗生物量的分配方式有明显的影响。根冠比是衡量植株生长状态特别是植株对土壤水分、养分状态反应的一个指标[17]。本研究结果显示,除了铵态氮和硝态氮浓度为10 g/(m2·a)时刺萼龙葵幼苗的根冠比小于无氮沉降处理,其他氮沉降处理的根冠比均大于无氮添加处理,说明氮沉降在一定程度上影响了刺萼龙葵幼苗的根冠比,而铵态氮和硝态氮浓度过大则抑制了刺萼龙葵幼苗的根冠比。
4 结论
综上所述,不同氮沉降处理对刺萼龙葵幼苗的生长产生了不同影响。所有处理对刺萼龙葵幼苗的株高无显著影响,所有处理均极显著降低了刺萼龙葵幼苗的根粗,且在处理13~处理15区间降低最为明显,二者交互作用和铵态氮处理对刺萼龙葵幼苗茎粗均具有极显著的降低作用,处理14降幅最大,所有处理均显著增大了刺萼龙葵幼苗的比叶面积,其中处理10的比叶面积最大。
此外,不同氮沉降水平对刺萼龙葵幼苗的生物量积累和分配影响不尽相同。在铵态氮和二者交互作用浓度为4~8 g/(m2·a)时,会增加刺萼龙葵幼苗生物量的积累与分配,促进其入侵,而铵态氮和二者交互作用的浓度均为10 g/(m2·a)时,会减少刺萼龙葵幼苗的生物量积累和分配,从而达到抑制其生长的作用,不利于其入侵。
猜你喜欢
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
中国诗歌(2018年6期)2018-11-14
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代农业科技(2017年16期)2017-09-22
国学(2016年8期)2017-03-20
环球人文地理(2015年11期)2015-09-10
湖泊科学(2015年1期)2015-05-06
江苏农业科学(2014年1期)2014-07-18
