
富含OPO、OPL和LPL的结构脂质促进C57BL/6小鼠脂质代谢
2024-04-03 12:02孙永贺杨正邓泽元李静彭小雨潘丽娜李威郭丹颖汪家琦
现代食品科技 2024年2期
孙永,贺杨正,邓泽元,李静,彭小雨,潘丽娜,李威,郭丹颖,汪家琦*
(1.澳优乳业(中国)有限公司,湖南长沙 410219)
(2.南昌大学食品科学与技术国家重点实验室,江西南昌 330047)
结构脂质是指通过改变油脂甘油三酯支链上脂肪酸的碳链长度和在支链上的位置从而改善营养功能性质的油脂[1]。与传统的天然油脂相比,结构脂质的脂肪酸组成和分子结构可以更精确地设计和控制,因此具有更多的营养和功能优势[2]。结构脂营养功能已有许多报导,如提高Sn-2 脂肪酸吸收利用,促进其他种类脂质吸收,降低血清TG 及TC水平等[3]。1,3- 二油酸-2- 棕榈酸甘油三酯(OPO)是一种特殊的甘油三酯,已被报道可用于降低高脂血症[4]。此外,课题组先前的研究表明具有不同立体构型和比例的甘油三酯对LO2 细胞的脂质代谢具有显著不同的影响。与Sn-2 位为棕榈酸的1-油酸-2-棕榈酸-3-亚油酸甘油三酯(OPL)、1,3-二亚油酸-2-棕榈酸甘油三酯(LPL)和OPO 相比,1-棕榈酸-2,3-亚油酸甘油三酯(POO)可以显著诱导脂质积累,而OPL 比LPL 和OPO 更容易诱导脂质积累[5]。但是这些结构脂质对小鼠的脂质代谢影响并不清楚。
当膳食中的甘油三酯(TAGs)被机体吸收后,它们会在在血液中被脂蛋白脂肪酶水解,生成甘油二酯(DAGs)。脂蛋白脂肪酶具有Sn-1,3 特异性,因此它主要水解TAGs 的Sn-1 位置,生成2,3-DAGs,而不水解TAGs 的Sn-2 位置的脂肪酸[6]。接下来,脂蛋白脂肪酶会进一步水解TAGs 的Sn-3 位置的脂肪酸,生成2-甘油单酯(MAGs)。然而,如果TAGs 的Sn-3 位置上的脂肪酸是花生四烯酸或二十碳五烯酸,脂蛋白脂肪酶很难水解这些酯键,因此不会形成2-MAGs[7]。如果血浆中肝脂肪酶的活性高,它可以进一步水解含有多不饱和脂肪酸(PUFAs)的DAGs 成为2-MAGs。然而,如果血浆中缺乏肝脂肪酶,乳糜微粒残留物中会含有大量PUFAs 的DAGs,并且在肝脏中被肝脂肪酶进一步水解成2-MAGs。这些2-MAGs 一部分会进入肝脏作为能源使用或重新合成其他脂质,另一部分则进入周围组织进行再酯化合成TAGs 或磷脂[7]。因此,膳食中的TAGs 会经历一系列代谢作用,其中它们的Sn-2 位置上的脂肪酸最终会在乳糜微粒的TAGs和组织中被保留。由于相同脂肪酸组成的TAGs 可能具有不同的结构,例如POP 和OPP,因此它们可能会影响血浆中血脂的清除、组织炎症和胰岛素抵抗等方面[8]。
本研究采用酶促酯交换反应制备了一种富含OPO、OPL 和LPL 特殊结构脂质(SL)的混合物,并将其添加至C57BL/6 小鼠的饲料中进行为期7 d的短期喂养。在此期间,监测小鼠的体质量变化,检测小鼠血清和肝脏生化指标,以及观察肝脏中脂质代谢相关蛋白的表达差异。本研究旨在通过分析这些影响,为开发基于消费者需求和油脂特性的个性化功能性食品和营养食品提供理论基础。
1 材料与方法
1.1 材料与试剂
大豆油,益海嘉里公司;InFat CC(一种市售特殊的脂肪酸结构,用于生产婴儿奶粉配方的产品,其OPO 含量≥40 g/100 g),澳优乳业(中国)有限公司;结构脂质(SL),实验室合成的一种富含OPO、OPL和LPL特殊结构脂质;乙醚、异丙醇、氯仿、甲醇、正己烷等色谱纯有机试剂,国药集团化学试剂有限公司;TC 含量检测试剂盒、TG 含量检测试剂盒、HDL-C 测定试剂盒、LDL-C 测定试剂盒、脂肪酸合成酶ELSA 试剂盒(FAS)、PPAR-αELSA 测试盒,南京建成生物工程研究所。
1.2 仪器与设备
Agilent 6890N 型气相色谱,美国Agilent 科技公司;Agilent G1260 高效液相色谱-G6430A 三重串联四级杆质谱联用仪,美国Agilent 科技公司;AL104精密天平,梅特勒-托利多;WGL-125B 干燥箱,天津市泰斯特仪器;台式电动离心机,金坛市华锋仪器;DF-101K 集热式恒温磁力搅拌器,郑州长城科工贸公司;QL-861 涡流振荡器,其林贝尔仪器制造公司;DSY-V1 氮吹仪,北京金科精华苑科技);UVmini-1240 分光光度计,日本岛津公司;DZKW-4恒温水浴锅,北京中兴伟业仪器)。
1.3 动物实验
1.3.1 实验设计
动物实验经江西中医药大学动物伦理委员会批准(SCXK 2017-0004)。无脂纯合日粮购买于小黍有泰(北京)生物技术有限公司。21 只5 周龄健康C57BL/6J 小鼠(湖南斯莱克实验动物公司,SCXK(湘)2019-0004),实验环境为普通环境,实验期间环境温度22~24 ℃,湿度52%~56%,实验动物自由摄食饮水。表1 列出了实验中喂养小鼠的无脂纯合日粮饲料的组成情况。另外每天给小鼠灌胃1 次, 每次灌胃剂量100 μL/10 g(油体积/小鼠体质量)。实验中,将小鼠分为3 组,每组7 只。在饲养期间每天测量1次体质量,以便观察变化。分组情况如表2 所示。

表1 实验小鼠日常饲料成分组成(每100 g含量)Table 1 Composition of experimental daily feed (100 g basis)

表2 实验小鼠分组Table 2 Grouping of experimental mice
1.3.2 血清及肝脏指标测定
严格按测定试剂盒使用说明书测定TC、TG、高密度脂蛋白胆固醇HDL-C、高密度脂蛋白胆固醇LDL-C、脂肪酸合成酶FAS 和过氧化物酶体增殖物活化受体αPPARα含量。
1.4 总脂肪酸组成的测定
在玻璃试管中称取2.0 mg 的油,加入1.5 mL正己烷,涡旋混合后加入40 μL 的乙酸甲酯并涡旋混合。接着加入100 μL 甲醇钠-甲醇,在37 ℃水浴中反应20 min。取出反应混合物,放置于-20 ℃的冰箱中10 min 后加入60 μL 的草酸-乙酸乙酯并涡旋混合,离心(转速为4 200 r/min,离心时间为5 min),取上清液过无水硫酸钠柱,用氮气吹干后加入1 mL 色谱级正己烷复溶于6 890 N 气相色谱仪中进行检测。色谱条件为使用CP-Sil88 柱(100 m×0.25 mm×0.2 μm),载气为N2,燃烧气为H2和空气,进样口250 ℃,进样量1 μL,FID 检测器温度为250 ℃,使用不分流模式。升温程序为:0~4 min,45 ℃;4~14 min,从45 ℃升温至175 ℃;14~41 min,保持在175 ℃;41~51 min,从175 ℃升温至215 ℃;51~86 min,保持在215 ℃。以标准品的保留时间为对照,鉴定脂肪酸的组成。
1.5 甘油三酯构型的测定
采用LC-MS/MS 测定TAG 组成,方法参照张东等[9],称取20 mg 反应粗产物,用1 mL 正己烷溶解后,转移至填充有500 mg 硅藻土材质的SPE-NH2固相微萃取小柱(3 mL 规格,上海安谱仪器有限公司)。采用正己烷和乙醚(85:15,V/V)混合的洗脱液进行洗脱,将甘油三酯吹干。取洗脱后的甘油三酯5~8 mg,用异丙醇稀释至1 mL,通过0.45 μm有机滤膜过滤。
液相条件:Zorbax Eclipse Plus C18,250 mm×4.6 mm×5 μm;溶剂A:异丙醇,B:乙腈;洗脱条件为0.00~25.00 min 65% B,25.00~30.00 min 35% B,30.00~30.10 min 65% B,30.10~40.00 min 65% B;流速:0.8 mL/min;进样量:5 μL;柱温:35 ℃。
MS 条件:APCI 电离源,正离子模式;雾化气流4 L/min,雾化电压20 psi;气化温度325 ℃,汽化室温度350 ℃;扫描模式,EMS-APCI 或者MRM-APCI;裂解电压90 V;碰撞能,35 V;mass范围,EMS 400~1 500m/z。EMS-EPI 模式用于定性分析,MRM-EPI 模式用于定量分析。
1.6 数据分析
实验结果表示为“平均值±标准偏差”,每组实验重复3 次。数据处理及方差分析采用GraphPad Prism 8 以及SPSS 26.0 统计分析软件包,图中不同字母表示具有显著性差异(P<0.05)。
2 结果与讨论
2.1 结构脂质的总脂肪酸组成分析
表3 显示了大豆油、InFat CC 和SL 中的总脂肪酸组成。结果表明,大豆油中含有大量的不饱和脂肪酸,其中相对含量最高的是亚油酸(C18:2),约占41.25%,其次是油酸(C18:1),约占23.03%,亚麻酸(C18:3),约占17.67%,这与王志高等[10]的研究结果一致,他们发现大豆油主要含有51.51%的亚油酸以及26.43%的油酸。InFat CC 主要含有棕榈酸(16:0)和油酸(C18:1),分别占总脂肪酸的39.30%和45.86%。而合成的结构脂质中主要脂肪酸为棕榈酸(16:0)24.43%、油酸(C18:1)33.46%和亚油酸(C18:2)25.82%。

表3 结构脂质中总脂肪酸组成(相对含量%)Table 3 Total fatty acids of structured lipids
2.2 结构脂质的甘油三酯组成分析
表4 显示了大豆油、InFat CC 和SL 具体甘油三酯组成。大豆油中甘油三酯相对含量最高的前3 种为OLLn、OLO 和OOO,分别占26.74%、23.09%和20.74%,且甘油三酯的2 位脂肪酸主要是不饱和脂肪酸。张东等[11]研究表明,大豆油中甘油三酯的2 位脂肪酸主要是不饱和脂肪酸,但相对含量较高的甘油三酯分别是LLL(23.19%~33.57%)、LLO(15.23%~22.74%)、PLL(9.73%~16.18%)和LLLn(9.06%~14.22%),造成这种差异的原因可能是检测方法的不同。InFat CC 中主要含有5 种甘油三酯,其中OPO(44.04%)和MPO(20.97%)为主要的两种甘油三酯(占比>60%)。结构脂质SL的甘油三酯种类最多,相对含量较高的前3 种甘油三酯为LPL(18.35%)、OPL(17.04%)和OPO(14.28%)。结果表明,InFatCC 和SL 中均含有较高相对含量的OPO 结构酯,Huang 等[4]研究表明OPO 结构酯具有显着降低高脂饮食小鼠血清TC、TG、LDL-C 水平以及增强抗氧化酶活性的功能。同时,InFat CC 和SL 含有的甘油三酯中2 位脂肪酸主要是饱和脂肪酸棕榈酸,这种独特的甘油三酯(TAG)结构对婴儿具有有益作用,例如解决便秘、增强钙吸收和骨骼发育、脂肪吸收和脂质代谢[12]。

表4 结构脂质甘油三酯组成(相对含量%)Table 4 Triglyceride composition ofstructured lipids
2.3 结构脂质对小鼠体质量变化的影响
图1a 表现出7 d 中每组小鼠的饮食量情况,可以看出彼此之间饮食量没有明显差异(大豆油组14.51 g/d,InFatCC 组14.73 g/d,SL 组15.11 g/d)(P>0.05),说明小鼠体质量的变化没有受到饮食量不同的影响。图1b 反应了不同油喂养期间小鼠体质量的变化情况,对大豆油和InFat CC 喂养的小鼠进行对比,发现其体质量的变化趋势均呈现缓慢增加的趋势,而SL 喂养的小鼠体质量变化呈现出下降趋势,喂养结束三组小鼠的平均体质量分别为27.29 g、26.54 g 和23.3 g,且SL 喂养的小鼠体质量显著低于大豆油喂养的小鼠(P<0.05)。结果表明,SL 可以控制小鼠的体质量增加,这可能是因为SL 的特殊甘油三酯结构和脂肪酸组成促进了小鼠脂质代谢,加速了能量代谢,从而控制体质量增加。大量临床和动物研究表明,结构脂质可以帮助控制体质量,抑制肥胖的发生[13]。Lee 等[14]的研究发现,与物理混合组相比,摄入棕榈仁油和棕榈油酯交换结构脂的小鼠体质量增长显著较少。然而,也有研究报道称,酯交换结构脂与物理混合油脂对大鼠的体质量没有影响[15,16]。
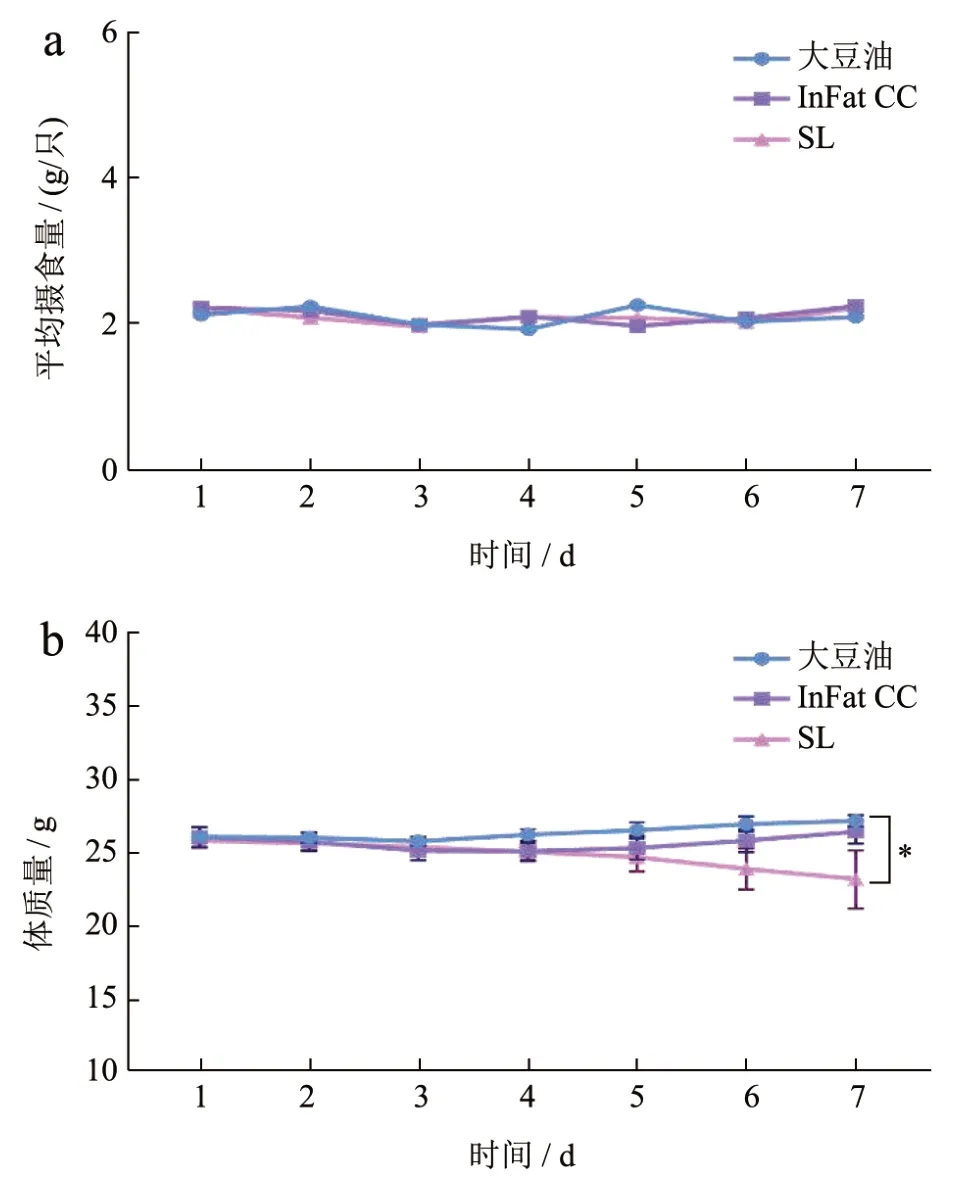
图1 小鼠摄食量和体质量变化Fig.1 Changes in food intake and body weight of mice
2.4 结构脂质对小鼠血清甘油三酯、总胆固醇、高密度胆固醇和低密度胆固醇水平的影响
由图2 可知,SL 组小鼠血清的甘油三酯水平(1.17 mmol/L)要显著低于InFat CC 组(1.54 mmol/L),与大豆油组(1.10 mmol/L)相比没有显著性差异。大豆油、InFat CC 和SL 组小鼠血清的TC、HDL-C 和LDL-C 浓度都没有明显的差异。结果表明,结构脂质能够降低小鼠血清TG 水平,而对小鼠血清TC、HDL-C 和LDL-C 浓度影响较小。Alfieri 等[17]综述了含有棕榈酸和硬脂酸的酯交换油脂和非酯化油脂的甘油三酯结构对人体健康的影响。他们的结论是,与原料非酯化的物理混合油脂相比,没有足够的证据表明酯交换油脂不会影响或降低空腹血脂水平,但可以降低受试者餐后血浆甘油三酯浓度,这与本研究结果相一致。此外,早期的几项研究报告表明,结构化脂质可以降低高血脂小鼠的脂质沉积,特别是甘油三酯、低密度脂蛋白胆固醇和总胆固醇水平,同时维持血清和肝脏中必需脂肪酸的状态[18]。

图2 结构脂质对小鼠血清甘油三酯(a)、总胆固醇(b)、高密度胆固醇(c)和低密度胆固醇(d)的影响Fig.2 Effect of structural lipid on the serum level of TG (a),TC (b), HDL-C (c) and LDL-C (d) in mice
2.5 结构脂质对小鼠肝脏甘油三酯和总胆固醇水平的影响
由图3 可知,与大豆油组和InFat CC 组相比,SL 组小鼠的肝脏甘油三酯(0.14 mmol/g prot)和总胆固醇(0.05 mmol/g prot)浓度显著降低,大豆油组和InFat CC 组小鼠甘油三酯(0.21 和0.19 mmol/g prot)和总胆固醇(0.07 和0.08 mmol/g prot)浓度均没有显著性差异,说明SL 能够降低小鼠肝脏脂肪和胆固醇的堆积。与上诉结果相似,Malongil 等[15]进行了一项大鼠实验,将含有椰子油和米糠油(或芝麻油)的酯交换结构脂和物理混合油脂添加到正常饮食中。结果表明,尽管结构脂和物理混合油脂的脂肪酸组成相同,结构脂仍然能够显著降低肝脏中甘油三酯的浓度。而SL 组小鼠胆固醇浓度显著降低,这可能是SL 能够促进肝脏胆固醇转化为胆汁酸从而降低小鼠肝脂水平[19],这需要进一步的研究来证明。此外,Wu 等[5]研究发现,在使用25 μmol/L 的结构脂质OPO、OPL 和LPL 分别处理LO2细胞24 h,三种结构脂质均表现出一定的降脂作用,在本研究中OPO、OPL 和LPL 是SL 含有的三种主要甘油三酯。Zhang 等[20]研究表明与富含油酸的甘油三酯相比,富含亚油酸的甘油三酯消化性较低,但吸收性较高,与InFatCC 相比大豆油和SL 均含有更高含量的亚油酸甘油三酯,这可能是大豆油和SL 对小鼠血脂影响程度更大的原因之一。
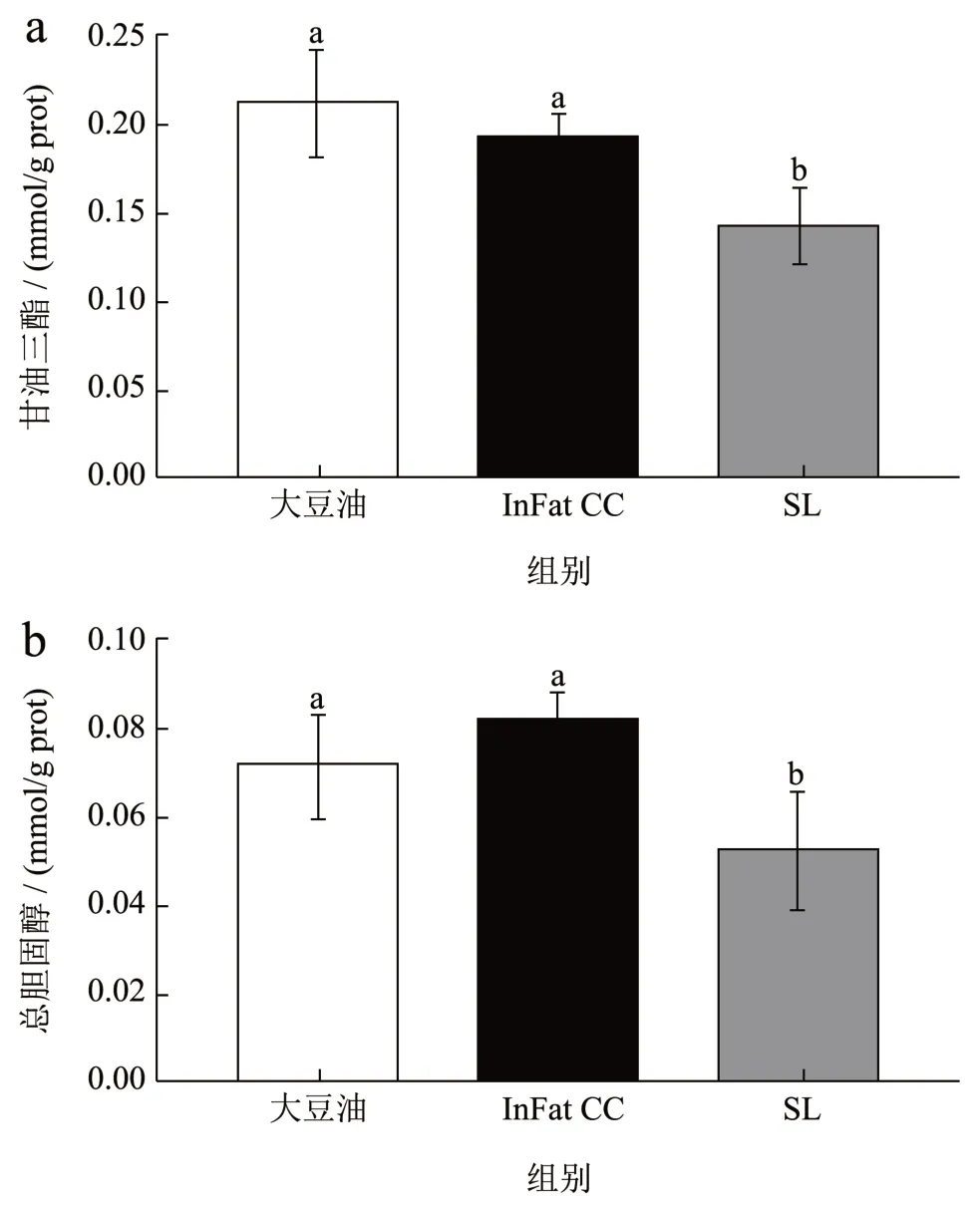
图3 结构脂质对小鼠肝脏甘油三酯(a)和总胆固醇(b)的影响Fig.3 Effect of structural lipid on the liver level of TG (a)and TC (b) in mice
2.6 结构脂质对小鼠肝脏脂肪酸代谢相关蛋白的影响
图4 的结果表明,与InFat CC 组(FAS 0.17 mg/g肝脏,PPARα0.05 mg/g 肝脏)相比,SL 组小鼠肝脏中FAS 质量分数(0.13 mg/g 肝脏)显著降低(P<0.05)而PPARα的质量分数(0.08 mg/g 肝脏)显著增加(P<0.05),说明SL 喂养的小鼠肝脏中脂肪酸的合成减少且脂肪酸氧化增加。FASN 基因调控脂肪酸的脂肪合成,该基因编码脂肪酸合成酶FAS,并催化棕榈酰辅酶A 的合成[21]。脂肪β-氧化是脂肪的主要代谢途径,PPARα是转录因子,能够直接转录参与脂肪酸β-氧化的基因,增加PPARα的表达或活性可以有效增强脂肪酸β-氧化[22]。总的来说,小鼠通过摄入SL 以后,可以增强其肝脏内的脂肪酸β-氧化并且抑制脂肪酸从头合成,这可能是SL 降低小鼠肝脏甘油三酯的一个主要原因。最近一项研究发现,SL 不但具有天然植物油的特性,而且还有一定的营养功能。例如降低血清胆固醇、减少体质量和促进脂质代谢等[23],根据这些功能,建议结构脂质可用作婴儿食品和药物制剂中的营养补充剂[24]。因此本研究能够为结构脂质用作婴儿食品和药物制剂中的营养补充剂通过理论依据。

图4 结构脂质对小鼠肝脏脂质代谢相关蛋白的影响Fig.4 Effect of structural lipids on genes related to liver lipid metabolism in mice
3 结论
通过动物实验研究表明,富含OPO、OPL 和LPL 结构脂质(SL)可以通过增强小鼠肝脏内的脂肪酸β- 氧化并且抑制脂肪酸从头合成,降低了小鼠肝脏内脂肪和胆固醇的堆积,从而达到抑制肥胖的作用。因此,本研究可以为结构脂质在减少体内脂质堆积、促进脂质代谢和抑制肥胖的新型食品开发应用提供理论依据。
猜你喜欢
锦州医科大学报(2021年8期)2021-11-18
小天使·一年级语数英综合(2020年11期)2020-12-16
祝您健康(2018年12期)2018-11-27
福建质量管理(2018年18期)2018-04-02
文理导航·科普童话(2017年4期)2018-02-10
散文诗(2017年17期)2018-01-31
风景园林(2017年3期)2017-06-22
中国油脂(2017年12期)2017-01-18
中国粮油学报(2016年5期)2016-01-23
食品工业科技(2014年13期)2014-03-11
