
m6A甲基化修饰蛋白在卵巢癌中的研究进展*
2024-04-01 09:40杨惠雯李安蓝婷
中国现代医学杂志 2024年6期
杨惠雯,李安,蓝婷
(徐州医科大学 医学技术学院,江苏 徐州 221004)
表观遗传学修饰包括DNA、RNA和蛋白质的化学修饰。其中RNA存在100余种化学修饰,RNA修饰是一种转录后水平的调控方式。N6-甲基腺苷(N6-methyladenosine,m6A)即RNA分子腺嘌呤第6位氮原子上发生甲基化修饰,是真核细胞mRNA最常见的转录后修饰,20世纪70年代首次在真核生物的mRNA中发现,占RNA甲基化修饰的80%。m6A甲基化修饰涉及甲基转移酶、去甲基化酶和甲基化阅读器,同时m6A甲基化修饰也被证明是一个可逆性过程。卵巢癌是女性生殖系统最常见且病死率最高的恶性肿瘤之一,大多数患者发现时已处于晚期,5年生存率不足50%,具有高复发率、高病死率[1-2]。近年来,越来越多的研究表明m6A修饰与癌症之间存在潜在联系。本综述将重点介绍m6A甲基化修饰对卵巢癌的增殖、转移的影响及其分子机制。
1 m6A甲基化修饰蛋白的组成及功能
m6A修饰是mRNA最常见的一种RNA修饰,m6A修饰具有保守基序RRACH(R表示A或G,H表示A、U或C),其分布富集于长外显子、终止密码子以及3′-非翻译区(3′-UTR),m6A甲基化根据其修饰的基因组位置不同调节不同的RNA过程和生物功能,5′-非翻译区(5′-UTR)的m6A甲基化修饰可以绕过5′-帽子结构,直接与真核起始因子3(eukaryotic initiation factor 3,EIF3)结合,促进蛋白质翻译的启动,位于编码序列的m6A修饰有利于RNA的稳定性和细胞增殖,位于终止密码子的m6A甲基化与3′-UTR处的多聚腺苷酸化有关[3]。m6A是个动态可逆的过程,m6A修饰的调节主要是由m6A甲基转移酶、去甲基化酶和结合蛋白组成(见图1)[4]。其中m6A甲基转移酶即“写入器”,催化m6A修饰的发生。m6A甲基转移酶包括甲基转移酶样蛋白3(methyltransferase-like 3,METTL3)、甲基转移酶样蛋白14(methyltransferase-like 3,METTL14)和Wilms肿瘤1相关蛋白(Wilms′tumor 1-associating protein,WTAP),m6A去甲基化酶即“擦除器”,如肥胖相关蛋白(fat mass and obesity-associated protein,FTO)和ALKB同系物5(AlkB homolog 5,ALKBH5)等蛋白质可以使细胞中mRNA的m6A水平降低,即发生了去甲基化。同时m6A结合蛋白即“阅读器”,包括YTH结构域蛋白家族成员YTHDC2、YTHDF1、YTHDF2、YTHDF3和异质性胞核核糖核蛋白、IGF2BP1等,负责读取识别m6A甲基化修饰。m6A修饰发生在转录和mRNA加工过程中的pre-mRNA阶段[5],该过程主要由m6A甲基转移酶复合体催化。
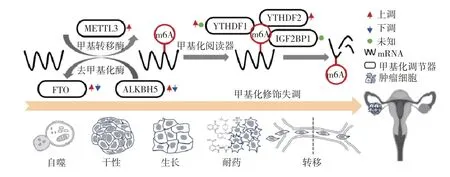
图1 m6A修饰相关蛋白的示意图
2 m6A甲基转移酶在卵巢癌中的研究进展
METTL3作为m6A修饰的写入器参与RNA生命周期的所有阶段,并通过调节关键癌基因或抑癌基因的m6A修饰来影响肿瘤进展,METTL3调节RNA的半衰期,并调控mRNA前体的剪切、miRNA处理、核转运、翻译和RNA降解[6]。m6A修饰通过影响剪接因子与其结合位点的结合来调控mRNA前体的剪切,敲低细胞中的METTL3后,导致选择性剪接变化,并且与具有选择性、剪接特性的外显子和内含子相比,组成性的外显子和内含子具有更多的m6A修饰。METTL3还能够对pri-miRNA上GGAC motif的腺苷酸(A)进行m6A修饰。m6A修饰能够促进pri-miRNA被DiGeorge综合征危象区基因8特异性识别,并影响后续成熟体miRNA的加工。因此,METTL3也可以调控miRNA前体并加速其成熟[7]。METTL3可以通过多种机制参与m6A修饰介导的mRNA翻译过程,包括依赖YTHDF1-EIF3调控翻译、与EIF3直接作用调控翻译、结合mRNA的蛋白质编码区(coding sequence,CDS)调控翻译[8-9]。此外,在METTL3催化的m6A中,m6A识别酶YTHDF2的羧基末端结构域能够选择性地与含m6A的mRNA结合,而氨基末端结构域则负责将YTHDF2-mRNA复合物定位到细胞质处理小体等RNA降解的场所,从而调节mRNA的降解[10]。METTL3在不同癌症中会发生表达上调或下调,所以其在肿瘤发生、发展中的具体作用仍存在争议。有研究表明,在胃癌、结直肠癌、膀胱癌及肝细胞癌等恶性肿瘤中,METTL3的表达水平上调[11],同时也存在METTL3在子宫内膜癌中表达降低[12]。在卵巢癌中,METTL3过表达后,m6A修饰水平明显上升并对肿瘤细胞的增殖和转移有促进作用。BI等[13]在分析对比75例卵巢癌患者和健康人的卵巢组织METTL3 mRNA水平后,得出METTL3在卵巢癌患者中的表达明显高于健康人。因此METTL3作为卵巢癌中的肿瘤促进因子,其表达水平与卵巢癌的进展密切相关。
关于METTL3是如何调控卵巢癌的进展,BI等[13]研究发现,METTL3通过介导pri-miR-126-5p的m6A修饰来调控miR-126-5p的表达,进而激活抑癌基因人张力蛋白同源物基因(phosphatase and tensin homologue deleted on chromosome 10,PTEN)介导的PI3K/Akt/mTOR通路,促进卵巢癌的进展[13-16]。除此之外,有研究表明,miR-1246在卵巢癌患者样本中高表达,METTL3可以通过调控miR-1246的m6A甲基化修饰来促进miR-1246的表达,进而促进卵巢癌细胞的增殖、侵袭和迁移,同时抑制其凋亡。METTL3可以识别pri-miR-1246的m6A修饰,并进一步促进pri-miR-1246的成熟,miR-1246通过与细胞周期蛋白G2(CCNG2)的3′-UTR区结合抑制CCNG2的表达从而促进卵巢癌的发生和转移[17]。在结直肠癌中,METTL3也通过上调pri-miR-1246的m6A修饰,催化miR-1246成熟,并促进结直肠癌的转移[18]。此外,HUA等[19]研究发现,METTL3通过调节受体酪氨酸激酶翻译和上皮-间充质转化,促进卵巢癌的生长和侵袭。SHEN等[20]发现存在磷脂酶A2激活蛋白(phospholipase A2 activating protein,PLAA)-METTL3-瞬时受体电位通道经典3(transient receptor potential channel canonical 3,TRPC3)轴调控卵巢癌的转移,METTL3介导的m6A修饰促进了TRPC3的表达,进一步促进卵巢癌的转移,而PLAA作为METTL3的上游分子通过靶向泛素化METTL3,使其降解,进而抑制由METTL3介导的对TRPC3 mRNA的修饰,抑制卵巢癌细胞转移。
METTL14是m6A甲基转移酶复合物的关键部分,负责与RNA结合[21]。METTL14表达增加已被证明抑制某些癌症的进展,但其会导致急性髓细胞增殖[22]。肌钙蛋白相关蛋白(trophinin associated protein,TROAP)被发现参与许多癌症的增殖、侵袭和迁移[23]。当METTL14过表达时,TROAP mRNA的3′-UTR区m6A修饰水平增加,进而降低TROAP mRNA的稳定性,抑制卵巢癌细胞的增殖[24]。
3 m6A结合蛋白在卵巢癌中的研究进展
目前,对卵巢癌中m6A甲基化修饰“读写器”的研究主要集中于YTHDF1和YTHDF2。YTH结构域能直接与RNA的m6A位点相结合。YTHDF1是m6A修饰结合蛋白,其可以结合到m6A修饰的转录本的终止密码子附近,整体分布与m6A修饰非常相似。而YTHDF2可以介导m6A修饰的mRNA降解,正常条件下,YTHDF2可与脱腺苷酸酶复合物及脱帽复合物蛋白共定位,并将其靶基因的转录本带入降解小体[10]来介导m6A修饰的mRNA降解。
YTHDF1可直接与翻译起始复合物作用,促进m6A修饰的RNA底物的翻译效率[25]。在乳腺癌、肝癌、前列腺癌等恶性肿瘤中存在YTHDF1的明显上调[26-27]。在LIU等[25]研究中,通过癌症基因组图谱数据库发现,YTHDF1在高级别浆液性卵巢癌中上调,表明YTHDF1是一种原癌基因。通过分析卵巢癌数据库发现YTHDF1表达与肿瘤分级、FIGO分期和总生存率相关。YTHDF1通过募集EIF3来调节mRNA的翻译,从而调控卵巢癌的进展。此外,有报道称YTHDF1通过结合蜗牛蛋白miRNA的CDS的m6A位点,并通过招募真核延伸因子来增强其翻译,表明YTHDF1可以通过调节翻译延长和翻译启动来发挥作用[28]。除此之外,有学者认为YTHDF1通过调控三元基序蛋白(tripartite motif-29,TRIM29)的表达来促进卵巢癌的顺铂耐药。TRIM29是一种致癌基因,在各种癌症中异常表达,TRIM29通过不同的信号通路促进或抑制不同的癌症进展[29-35]。有研究发现,TRIM29在顺铂耐药卵巢癌细胞中的表达上调,而在敲除TRIM29之后显著抑制了卵巢癌细胞的干细胞样特征,并且抑制了顺铂耐药卵巢癌细胞皮下植瘤的裸鼠肿瘤的体积[35]。顺铂耐药卵巢癌细胞中YTHDF1对TRIM29 mRNA的招募增加,YTHDF1通过识别卵巢癌细胞中TRIM29的3′-UTR区,促进TRIM29的翻译[35]。
YTHDF2可以介导m6A修饰的mRNA降解,有研究表明,在卵巢癌细胞中敲除YTHDF2后,其mRNA的m6A修饰水平上调,并且卵巢癌细胞的增殖明显减少,凋亡增加,敲除YTHDF2还抑制了卵巢癌细胞的迁移和侵袭;而当YTHDF2过表达时,则得到相反的结果。在LI等[36]研究中发现,卵巢癌中miR-145的表达与YTHDF2呈负相关,当YTHDF2过表达后miR-145的表达水平降低,而当miR-145过表达后YTHDF2的表达水平又受到了抑制,表明在YTHDF2与miR-145之间存在着一种双向负反馈调节机制:miR-145以YTHDF2为靶点,靶向结合YTHDF2的3′-UTR,从而抑制YTHDF2的表达,YTHDF2介导m6A修饰的mRNA的降解,使miR-145降解,从而促进了卵巢癌的发生、发展。
4 m6A去甲基化酶在卵巢癌中的研究进展
FTO最初被称为肥胖相关蛋白,是第一个发现的m6A去甲基化酶[37],可以使单链RNA上的m6A修饰位点去甲基化。目前研究表明FTO在恶性肿瘤中的表达情况并不一致,在胶质母细胞瘤、白血病和乳腺癌中均有着促进肿瘤发生的功能[38-39],而在高级别浆液性卵巢癌中FTO的表达明显低于正常输卵管上皮。据HUANG等[40]研究发现,FTO对卵巢癌细胞的转录有显著影响,包括与干细胞信号传导、RNA转录、mRNA剪接和DNA修复相关的通路。FTO过表达时,可以使磷酸二酯酶4B(phosphodiesterase 4B,PDE4B)和磷酸二酯酶1C(phosphodiesterase 1C,PDE1C)m6A甲基化修饰水平降低并处于低表达状态,抑制由PDE4B和PDE1C介导的第二信使环磷酸腺苷(cyclic adenosine monophosphate,cAMP)及下游通路的激活,从而保持高水平的cAMP,抑制卵巢癌的发生、发展。因此PDE4B和PDE1C可以作为FTO调控m6A修饰的潜在靶点[40]。然而,也有研究表明,FTO在卵巢癌中起着关键的致癌作用,过表达时显著促进卵巢癌细胞增殖,但对FTO是如何作为m6A修饰的去甲基化酶促进卵巢癌的发展还不明确[41]。
除FTO之外,同样具有m6A去甲基化作用的ALKBH5也调控着恶性肿瘤的发生、发展。ALKBH5在上皮性卵巢癌组织中过度表达,并通过抑制上皮性卵巢癌细胞的自噬促进癌症进展[42]。研究发现,在卵巢癌细胞中存在着ALKBH5-同源框A10(homeobox A10,HOXA10)双向调节环路,ALKBH5通过维持HOXA10 mRNA的稳定性介导HOXA10上调,HOXA10作为转录因子与ALKBH5的启动子区域相结合,从而上调ALKBH5在卵巢癌细胞中的表达[43]。JAK2/STAT3信号通路已被广泛证明与各种恶性肿瘤的肿瘤生长和化疗耐药性有关。ALKBH5过表达降低了卵巢癌细胞中JAK2 mRNA 3′UTR区的m6A修饰水平,并通过降低YTHDF2介导的mRNA降解来维持JAK2 mRNA的表达。ALKBH5-HOXA10过表达后激活JAK2/STAT通路,进而导致上皮性卵巢癌的化疗耐药性[43]。SUN等[44]研究发现,ALKBH5过表达促进卵巢癌淋巴结转移。进一步研究发现,ALKBH5通过转录后机制下调了整合素β1(Integrin β1,ITGB1)mRNA的m6A修饰水平,抑制了YTHDF2蛋白介导的m6A依赖的ITGB1 mRNA降解,导致ITGB1表达增加,并磷酸化了局部黏着斑激酶和酪氨酸受体激酶原癌基因蛋白,增加了卵巢癌淋巴结转移。
5 总结
综上所述,m6A修饰作为真核细胞最常见的转录后修饰,近年来在恶性肿瘤发生、发展中的作用引起了极大关注,在卵巢癌中,m6A修饰蛋白通过调节关键癌基因或抑癌基因促进或抑制卵巢癌的发生或转移。目前的研究发现,METTL3、YTHDF1、YTHDF2和ALKBH5通过不同机制促进卵巢癌的进展,METTL14则抑制卵巢癌的发展,FTO在卵巢癌的作用尚不明确,促进及抑制均有报道(见表1)。虽然m6A在卵巢癌中的作用正逐渐被揭示,但其分子机制并未完全明确,需要在以后的研究中进一步探索,其可能通过其涉及的上游刺激因子、下游信号级联、癌细胞和肿瘤微环境等不同的甲基化修饰蛋白作用的靶点,明确其在卵巢癌中具体的作用。

表1 卵巢癌m6A修饰相关蛋白

续表1
猜你喜欢
保健与生活(2022年11期)2022-06-09
生物学通报(2020年10期)2020-08-13
广东农业科学(2017年10期)2018-01-25
国际妇产科学杂志(2016年2期)2016-06-16
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
现代检验医学杂志(2015年2期)2015-02-06
沈阳医学院学报(2014年4期)2014-12-27
遗传(2014年3期)2014-02-28
遗传(2014年3期)2014-02-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
