
洞庭湖DOM 组成与有机碳收支对江湖水力连通的响应
2024-03-28 08:09:46郭燕妮周永强太原师范学院地理科学学院山西晋中030619中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室江苏南京10008
中国环境科学 2024年3期
庄 奔,李 冰,郭燕妮,周永强,谷 娇(1.太原师范学院地理科学学院,山西 晋中 030619;.中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,江苏 南京 10008)
湖泊是全球碳循环的重要组成节点,其面积仅占除两极冰盖外全球陆地面积总和的3.7%[1],却是陆地碳循环的热点及陆地与海洋间重要连接枢纽与输送通道[2-3],在全球碳循环中发挥着重要作用.有研究表明,湖泊等内陆水体每年输移、埋藏及排放的碳通量为5.1Pg C /a,占陆地年净固碳通量的70%,向大气排放的碳通量为3.9Pg C/a[2,4].相较于非通江湖泊而言,通江湖泊通常具有水力滞留时间较短,河湖间水交换快,营养盐及有机碳等生源要素流动性强等诸多特点.分析水体交换如何影响这类湖泊中溶解有机物(DOM)的平衡与输出,有助于更好地了解内陆水体环境中碳元素的迁移过程.
溶解性有机物(DOM)主要成分为溶解性有机碳(DOC),由腐殖酸、富里酸、脂肪族及芳香烃等一系列复杂的有机物组成[5].DOM 经微生物和光化学降解能释放大量小分子有机酸和无机碳、氮、磷等营养元素,从而影响湖泊富营养化过程.湖泊DOM的来源和组成在很大程度上受湖泊和河流间的水力联系的影响,同时受流域人类活动的显著影响,因而具有高度的空间异质性[6].湖泊是特大和大中型城市最重要的集中式供水源,其水体中过量DOM 赋存通常使得水体酸臭刺鼻,并在水处理过程中产生致癌物质,严重威胁用水安全[7].降水过程引起地表径流的增加可能会导致土壤有机质、腐殖酸和生产生活废水等其他有机物向湖泊输入的通量增加,并在一定程度上提高土壤腐殖酸在DOM 中的占比[8-10].除陆源输入外,藻源DOM 在湖泊DOM 库中占比亦较高,尤其在水力滞留时间较长的富营养湖泊中表现得尤为显著[3].
洞庭湖与长江相连,是我国第二大淡水湖,在汛期对干流洪峰有显著调节作用[11].汛期周边水体自湘江、资水、沅江、澧水四个支流以及松滋、虎渡、藕池汇入洞庭湖,并最终经城陵矶排放到长江干流.由于三峡水库蓄水调度运用,洞庭湖不同湖区的水力滞留时间从0.1 年到0.5 年不等[12].相较于传统的CODMn、BOD5等水化指标而言,DOM的光学测量方便快捷,且能提供关于DOM 组成来源的大量指纹信息.DOM 光化学活性和微生物活性与其组成、来源特征息息相关[13].目前国内外围绕湖泊DOM 来源组成特征已开展了大量的研究.越来越多的研究利用傅里叶变换回旋共振质谱(FT-ICR MS)结合电喷雾电离技术对各种淡水生态系统中DOM 的分子组成进行了研究[14-15].针对洞庭湖的研究水体研究目前主要集中在水沙平衡、水体环境治理以及部分水体的富营养化.然而,以洞庭湖为代表的通江湖泊与河流间的水力连通特征如何影响湖泊DOM 组成及碳收支的研究目前尚未涉及.此本文以大型通江湖泊洞庭湖为研究对象,于不同季节自湘、资、沅、澧四支流和长江荆南松滋、虎渡、藕池三口与出湖口城陵矶处采集水样,通过对DOM 光学组成的测定以阐明洞庭湖与长江干流之间的水力联系如何影响洞庭湖DOM 组成及碳收支,旨在为保障湖泊供水安全、揭示湖泊碳收支提供参考.
1 材料与方法
1.1 研究区域
洞庭湖(107°16′~114°17′E,24°38′~30°26′N)位于长江中游,天然湖泊总面积2625km2,总蓄水量167 亿m3,为周边两千多万居民提供水源[16].洞庭湖流域年、降水量为1148~1837mm,2018 年洞庭湖主控站年径流量1740亿m3,其中荆南三口汇入洞庭湖径流量占全湖总来水量占比为29.0%,湘资沅澧四水径流量占洞庭湖来水量为71.0%(http://www.cjw.gov.cn/).

图1 洞庭湖采样点位置及江湖水力联系Fig.1 Location of sampling sites in Lake Dongting and the hydrological connection between the lake and the Yangtze River
1.2 样品采集与处理
本研究于2018 年5 月、7 月、9 月和11 月开展了4 次采样活动(n=56,个别月份湖泊水位偏高或偏低,对应采样点位略有偏移和增减).因水力收支在各来水与出水口处变化最为活跃,采样点主要分布在南洞庭湖和西洞庭湖各大河口.采用1.5L 酸洗后聚氯乙烯瓶采集水样,并立即置于4℃车载冰箱中避光储存,并在采集结束后迅速送实验室.水样首先经高温灼烧(450℃烧4h)的0.7µm孔径的Whatman GF/F玻璃纤维滤膜过滤,用于测定溶解性有机碳(DOC)浓度,随后使用水洗后的0.22µm 孔径的Millipore 膜过滤,用以测定DOM 吸收光谱和三维荧光光谱.
1.3 水文资料的获取
洞庭湖松滋、虎渡、藕池荆南三口及湘、资、沅、澧四水多年平均月径流量及洞庭湖流域多年平均逐月降水量等水文数据采集自全国水雨情信息网(http://xxfb.mwr.cn/sq_djdh.html).
1.4 微生物培养实验
DOC 微生物活性水平(biodegradable DOC,即BDOC)的概念为室温避光好氧环境下,28 天时间内快速被微生物降解的DOC 占初始(即培养前)DOC值的百分比,用以反映湖泊等水体中DOC 的微生物可降解潜力[17].BDOC 的测定采样国际规范流程操作,即样品采集后取100mL 经0.22µm 孔径滤膜的滤后液置于120mL 经酸洗和高温灭菌的的棕色玻璃瓶中,随后加入2mL 菌种(菌种为对应样点的原水).为避免微生物培养过程中存在的营养盐限制,通过添加2mL 营养液将培养瓶中营养盐浓度增加至80µmol/L 的NH4+-N 和10µmol/L 的PO43--P[18].通过每日轻晃数次并拧松玻璃瓶盖以确保培养过程中维持好氧环境,同时避免蒸发引起DOC 浓缩从而影响BDOC 的测算.样品经过28d 的微生物培养后重新经Whatman GF/F(孔径0.7 µm)滤膜过滤,进行光学测量,包括DOM 紫外-可见吸收光谱、三维荧光光谱和溶解性有机碳DOC,以确定DOM 的微生物活性BDOC.
1.5 紫外-可见吸收光谱参数的测定与计算
DOM 样品经0.7 µm 过滤后采用高温(680℃)燃烧法,在NPOC 模式下通过TOC-L 分析仪(日本岛津)测定DOC 浓度.采用岛津Shimadzu UV-2550UV-Vis 分光光度计及配套的5cm 石英比色皿完成DOM吸光度的测定.以Milli-Q 水为空白对照,在200~800nm 范围内每隔1nm 测量其吸光度,扣除700nm 出吸光度以消除潜在颗粒物散射效应.由于DOM 化学成分复杂,通常以特定波长的DOM吸收系数通常用于表征DOM 中带发色团组分的丰度[19-20].本研究中采用254nm 处的吸收系数表示DOM 相对丰度[21-22],根据公式(1)计算对应波长吸收系数[23]:
式中:aλ表示波长为λ 对应的DOM 吸收系数,m-1;Dλ代表对应波长λ 处矫正后吸光度;r 为光程路径,m.
DOM光谱斜率(S275-295µm-1)是由275~295nm波长范围内经指数函数拟合得到[24],DOM 在紫外及蓝光波段吸收光谱波长呈现指数函数递减.S275-295与DOM 腐殖化呈负相关,根据公式(2)计算[25]:
式中:λ 是测量波长,nm;λ0是440nm 处的参考波长,nm;S 表示光谱斜率.
1.6 三维荧光光谱的测定及平行因子分析
使用Hitachi 公司生产的F-7000 荧光光度计测定DOM 三维荧光光谱,其激发波长范围设置为230~450nm,间隔5nm,发射波长范围设置为300~550,间隔1nm.拉曼散射通过扣除当日纯水EEMs予以去除,而瑞利散射通过drEEM 工具箱的线性插值予以消除[26].内滤波效应采用对应样品激发、发射波长下的吸光度予以校正.平行因子分析(PARAFAC)将EEMs 矩阵分解为若干个具有唯一发射波长荧光强度最大值的荧光峰[27].采用MATLAB R2015b 中内置的drEEM 工具箱(ver.0.1.0)完成PARAFAC 建模[26].本文中PARAFAC 数据集包括56 个样品.每个矩阵对应于45 个激发波长和32 个发射波长.数据集分为6 个随机子集,3 个子集用于建模,其余3 个子集用于验证PARAFAC 模型.结果表明,3 个组分均能很好地通过对半检验(split-half analysis)、随机初始化分析(random initialization analysis)和残差分析(residual analysis).
1.7 数据分析
采用 SPSS 23.0 软件进行统计分析,包括平均值、标准差和t 检验,使用 ArcGIS 10.7 软件进行采样点的空间分布、不同月份DOM 通量变化模式、DOM 光谱参数的时空分异图的绘制.柱状图、主成分分析等采用Origin 2018 软件进行绘制.
2 结果与分析
2.1 水文条件
5 月和7 月,北部荆江段3 条主要入湖河流(松滋、虎渡、藕池)和湖南四水(湘江、资水、沅江、澧水)之和高于出水口城陵矶径流量,而在9 月和11月,北部三口与南部四水径流量与出水口城陵矶流出量大致持平(图2).洞庭湖区降水和径流的变化特性基本相似,径流量总体呈现出明显的下降趋势,而径流的变化也直接作用于湖泊本身,影响着洞庭湖水力的收支[28].
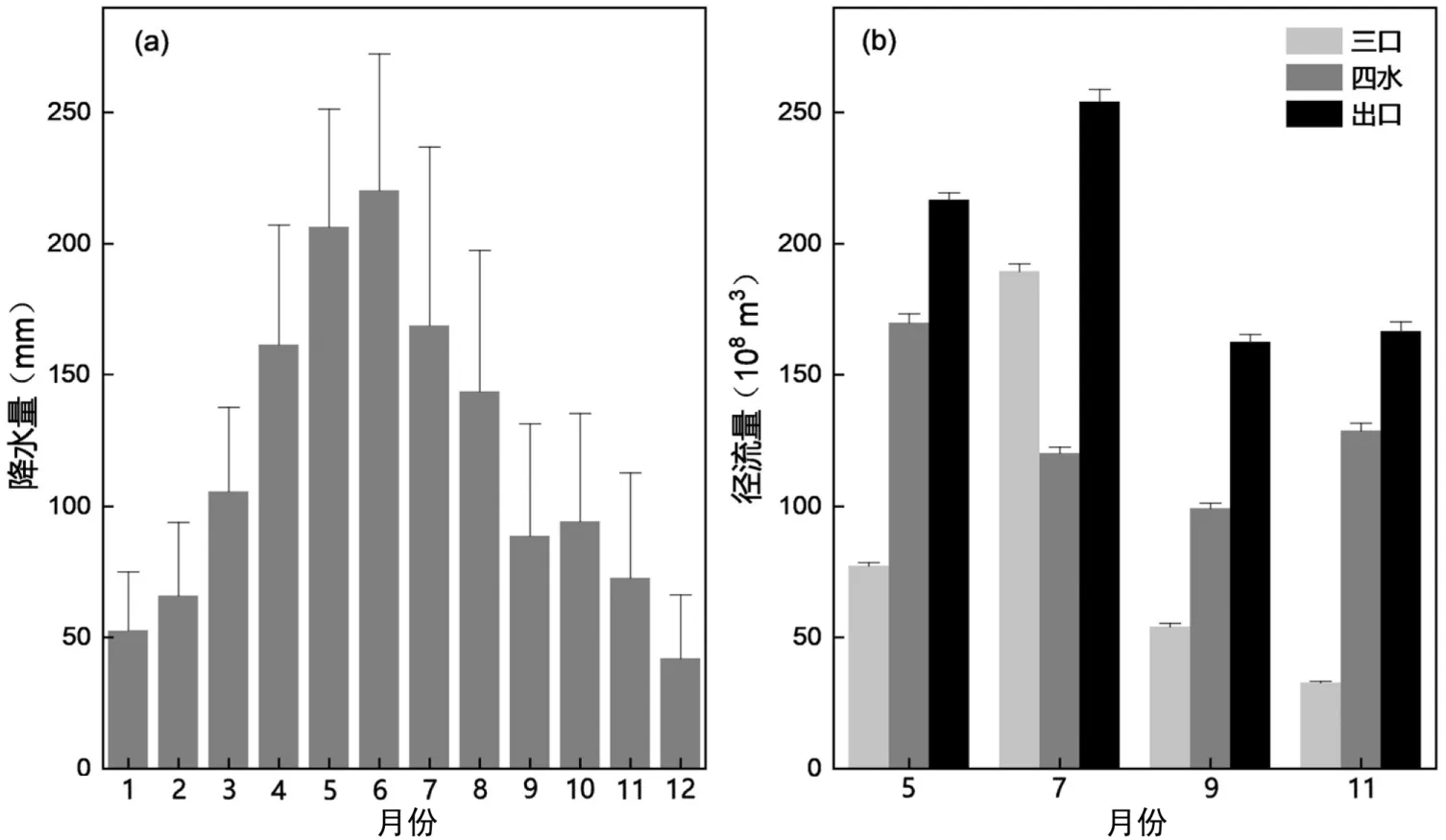
图2 洞庭湖流域多年(1961~2018)平均逐月降水量及2018 年5 月、7 月、9 月和11 月洞庭湖输入河与出水口月径流量Fig.2 Multi-year mean monthly rainfall of the Lake Dongting watershed in 1961~2018 and monthly runoff of input rivers and the sole outlet of Lake Dongting in May, July, September, and November 2018
2.2 PARAFAC 的荧光成分
应用EEMs-PARAFAC 对生物培养前水样进行荧光分析,得到3 种荧光组分(图3),包括一个类腐殖酸组分C1,两个类蛋白组分C2 和C3.C1 激发和发射波长分别为250nm 和420nm,组分C1显示出较强的陆源特征,同时也包含微生物或浮游植物的降解物[29].组分C2 和C3 是类蛋白组分,C2(Ex/Em=280/316nm)是类酪氨酸,C3(Ex/Em=230/340nm)是类色氨酸,二者广泛存在于河流与海洋中.组分C2 与类酪氨酸的荧光峰(峰B)一致,主要来自于原生地,与藻类降解和微生物矿化及城市污染物排放有关[30-31].C3 的光谱形状(Ex/Em=230/340nm)与类色氨酸相似,与传统色氨酸类单体的T 峰相对应[32].

图3 平行因子分析所得3 个荧光组分光谱特征及对半检验结果Fig.3 Spectra shapes of the three fluorescent components identified using PARAFAC and the three-component model was well validated using split-half validation procedure

图4 洞庭湖不同来水条件下DOM 光谱参数和荧光成分强度的空间分布Fig.4 Spatial distribution of DOM spectral parameters and fluorescence intensities of PARAFAC-derived components in Lake Dongting during different sampling seasons
2.3 不同江湖水力连通条件下的DOM 组成特征
洞庭湖区多年降水和径流的变化特性基本吻合,都表现为年内分布不均、且长期降水变化趋势不明显的特征[28].采样期间,7月入湖河流DOM光谱参数和荧光成分强度差异较大,汛期5 月和7 月a254浓度高于9月和11 月,长江荆南松滋、虎渡、藕池3 个入湖口显著高于湘、资、沅、澧四支流(t 检验, P<0.05).总体而言,9 月S275-295高于其他月份,这意味着当径流量较小时,陆地类腐殖酸输入信号也随之显著减弱.
7 月DOM3 个组分的荧光强度高于5 月、9 月和11 月,且在3 个入湖口处的强度均较高,这与输入河流中a254的空间分布相似,即在径流较小的9 月和11月,径流量提升会促进DOM浓度升高和外源输入的增加.DOM 浓度与类蛋白含量息息相关,类蛋白含量受到生活污水以及少量工业企业废水排放等高强度人类活动干扰[28].7 月湘江C2 和C3 荧光强度较高,对输入河流的贡献率为36.3%,表明湘江上游排放的生活污水排放较多.
2.4 洞庭湖汛期生物降解前后DOM 光学参数和荧光强度的变化
在汛期入湖与出湖河流中类腐殖酸组分通常占主导地位.荆南松滋、虎渡、藕池三口的腐殖质荧光强度高于湖南湘、资、沅、澧四水.不同之处在于,类蛋白质成分C2 和C3 在四条河流较高,而洞庭湖流域土地利用类型变化中,人类活动中建设用地比例相较耕地、林地等变化增量最多,表明四条河流可能吸纳了更多来自上游建设用地的生活污水.洞庭湖中DOM 丰度和DOC 浓度以及3 个组分的荧光强度结果均表明,河流输入的总和远高于湖的输出的总和.
如图5 所示,微生物降解并没有改变类腐殖质作为洞庭湖DOM 库主要成分的现状.微生物降解后,DOM 的3 种组分的荧光强度总体上均呈现下降趋势.与湖南四水相比,荆南三口DOC 更容易被生物降解.而在湖南四水中,生物降解前的DOC(2.1±0.8)mg /L 和生物降解后的DOC(1.6±0.3)mg/L 之间差异显著(t-test,P<0.05).在3 个入湖口中,类腐殖质C1 在微生物降解后降幅最大,为34.7%,而酪氨酸类C2 在四条入湖支流和出水口最易被微生物降解,降幅分别为36.7%和51.2%.

图5 汛期生物降解前后荆南三口、湖南四水和城陵矶的DOM 光谱参数和荧光成分强度Fig.5 DOM spectral parameters and fluorescence intensities of PARAFAC-derived components of the three inlets, four rivers, and the sole outlet at Chenglingji before and after biodegradation during the flood season
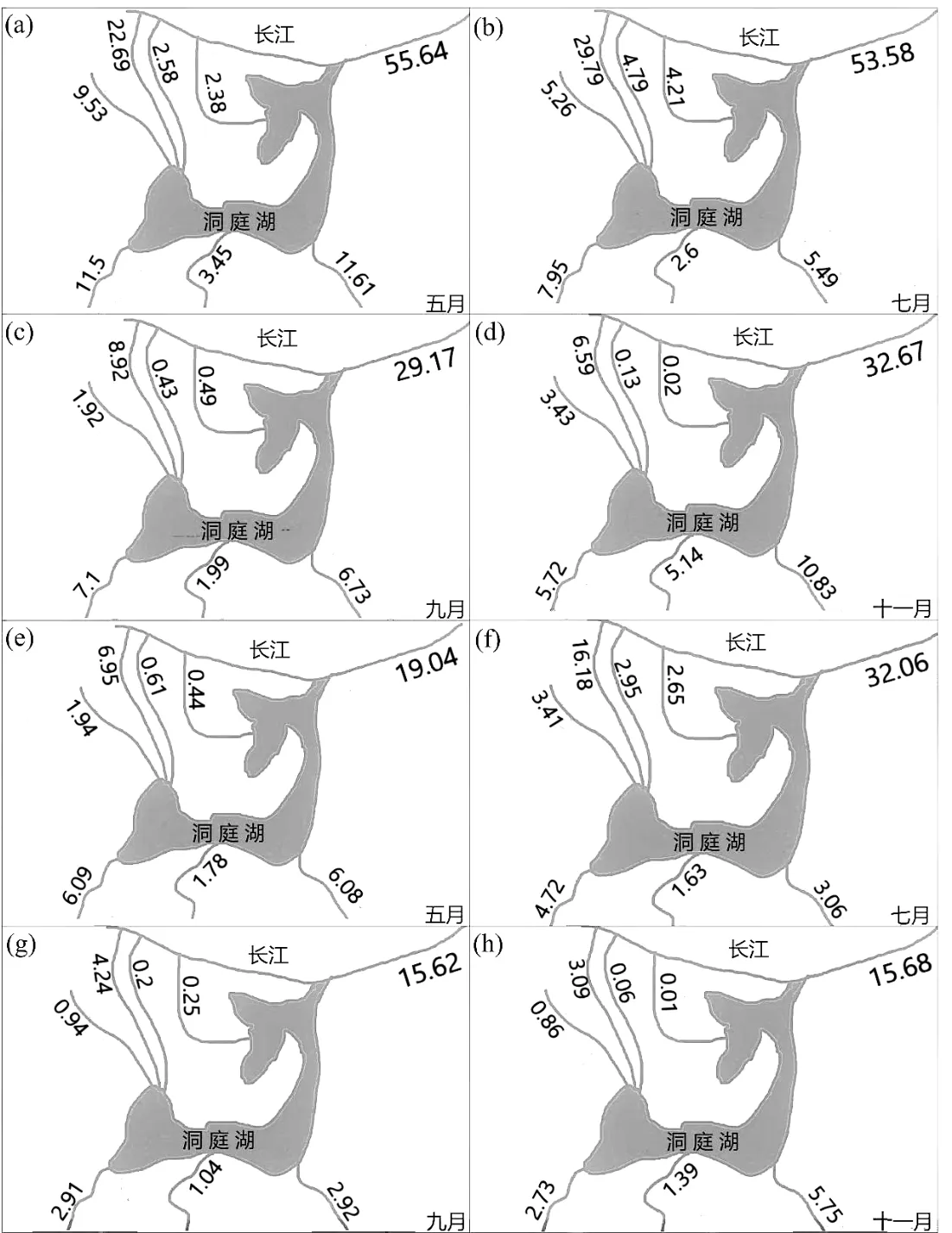
图6 5 月、7 月、9 月和11 月长江松滋、虎渡和藕池3 入湖口及湘、资、沅、澧四支流不同月份DOC 通量和DOM 吸收值通量Fig.6 DOC fluxes and DOM fluxes of rivers(Songzi, Hudu, and Ouchi) from the Yangtze River and four rivers during different runoff in May, July, September, and November
2.5 洞庭湖蓄水对DOM 吸收系数和DOC 浓度的影响
洞庭湖河流输入和输出的a254变化范围为6.0~14.6m-1,而DOC 浓度为1.3~4.7mg/L.在湖南四水中,湘江径流输入对全湖5 月的a254通量具有重要影响.城陵矶的a254在7月达到最大值12.6m-1.湖泊蓄水后,a254浓度与径流量的变化一致(图7),但DOC 浓度和径流量的变化并不完全一致.与总输入径流量不同,5 月入湖DOC 浓度最大,这意味着径流量变化对洞庭湖DOM 输出具有较强的主导作用,但在DOC 上未得到体现.而9 月径流量最小,洞庭湖蓄水对输入与输出的DOC 和a254的影响最小,其次是11 月.
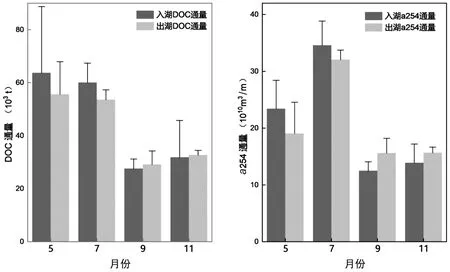
图7 5 月、7 月、9 月、11 月洞庭湖蓄水对DOC 和a254 入出湖通量影响Fig.7 The effect of Lake Dongting storage on DOC flux and DOM flux in May, July, September and November
3 讨论
3.1 洞庭湖的DOM 特征
江湖水力连通特征变化对洞庭湖DOC 浓度和DOM 和光学组成具有深刻影响.相较于9 月和11月,5 月和7 月松滋河存在更高的DOC 浓度和a254(图6),且同时期入湖径流量也呈现相似结果,a254表现为丰水期最大、枯水期最小,表明水位与水量的升高,即来水量增多使得DOC 浓度上升[33].5 月和7月藕池河和唯一出湖口城陵矶的DOC 浓度和a254均比其他样品更高,同时期输入径流量也较大.7 月湖泊的汇入径流量最大,3 个入湖口的S275-295和类腐殖质组分C1 显示出强烈的陆源特征.当流入径流量增加时,洞庭湖入湖支流携带土壤腐殖质和有机物的能力大大增加[25].当其他条件不变,径流流出流量越大,城陵矶类腐殖质C1的荧光强度也越高.该结果与近年来的研究结论一致,即径流增加会增加陆源DOM输入[31,34].对比5月和11月类腐殖质荧光强度,结果表明径流量变化与温度等外部条件皆会对DOM 输入产生影响[35].11 月径流量较低,表明陆源腐殖质外部输入量相对较小,其可能是在微生物作用下,浮游生物(如湖泊中的藻类)降解产生的腐殖质,而类腐殖质的积累过程快于类蛋白消耗的过程.同一时期藕池河色氨酸组分C3 在11 月中的荧光强度最弱,这进一步加速了这一时期微生物降解过程中类蛋白的降低和类腐殖质的积累[36].Mayer 等[37]也在其研究中发现微生物氧化分解会产生类腐殖质物质,因此这可能是由于11 月微生物群落的衰变导致了DOM 的降解.
在汛期来临的7 月和11 月,类腐殖质组分C1在进入湖泊的支流中荧光强度最高.夏季,湘、资、沅、澧四支流都以较大的径流量进入湖泊,导致外源输入中的人类建设与生活用地的废水输入量增大[38].同时,长江的高水位会导致长江荆江段上游带来的外源物质涌入洞庭湖及其支流.在此条件下,DOM 的生物地球化学行为和湖泊储存可能更加复杂,以至于城陵矶在7 月中没有表现出较高的类腐殖质来源.此外,由于高温湖泊微生物增殖,且微生物对腐殖质类降解影响较大,因此城陵矶腐殖质含量变低[39].11 月,该湖的总外部径流输入几乎等于出湖口城陵矶径流(图2).因此,从输入河流带入洞庭湖的DOM 能被有效地转移到长江下游.Lambert 等[40]通过对热带河流的研究,发现大型水库DOM 的通量和组成会受到水力收支的影响,这与本文的结果相似.在5 月和7 月,来自藕池河的径流量增加在一定程度上稀释了外源类腐殖酸输入信号[41],并且DOM 整体表现出较低的芳香性.同时期藕池河DOM 浓度显著高于其他出入口河流,而光谱斜率S275-295也相应呈现最大值(图4).
3.2 DOM 生物降解与江湖水力连通间的联系
汛期由于荆南三口有大量上游来水,与湖南四水相比,荆南三口DOC 更容易被微生物降解.强烈的江湖水体交换增加了水体中的溶解氧含量,更有利于微生物的生存[42].此外,汛期荆南三口DOC 外部输入的增加也极大地提高了微生物活性[43].湖南四水DOC 浓度在生物培养前后有显著差异,可能是由于湖南四水DOM 中类蛋白占比较高,更容易被微生物利用[36,44].经过微生物降解后,3 种荧光成分的荧光强度均有所减弱,说明洞庭湖的DOM 不稳定且易于被微生物降解,且类腐殖质仍是DOM 库的主要成分.相关研究发现,微生物降解可使DOM 趋于腐殖化[45],且腐殖化程度主要受光降解的影响[46-48].类腐殖质组分C1 在汛期的总体下降也可能是由于单一入湖支流在7 月径流量显著下降,减弱了其他径流的增加效果.张亚等[49]研究发现,在外来补水较少的非汛期,DOM 主要以自生源为主,而在汛期则受到陆源和自生源的共同影响,微生物在DOM 转化中扮演重要角色.Ni 等[50]在其研究中发现,一些大分子DOM 在培养56 天后被部分降解,说明温度和营养状况激发了 DOM 中难溶成分降解的驱动因素.Wilson 等[51]研究发现,在遮光培养后,类腐殖质的荧光强度降低,说明相关化合物可能更容易被微生物转化.湖南湘、资、沅、澧四水,在5 月生物降解后荧光强度明显减弱,从而印证了上述观点.少部分杆菌和生物细菌主要降解或生成类腐殖质DOM[52],在DOM 的光降解过程中,色氨酸样组分更容易被光降解[53],说明微生物对进入湖泊的河流中的荧光成分具有选择性,这种现象与DOM 所在的水体环境有关,例如温度和营养状况.
3.3 湖泊蓄水与DOM 和DOC 通量
7 月湘江的DOC 浓度较高,表明径流的增加也将增加外源输入的DOC[40].湘江受周围陆地环境输入的影响比其他河流更大,上游地区沿河排放的污水荧光强度更高[54].采样期间径流量变化对a254的输出有重要影响,然而在径流量较大时期,水力变化并不是影响DOC 输出的决定性因素.洞庭湖5 月DOC 调节能力最弱,其次是7 月,这是由于7 月的径流量最大,而长江补水效应加速了水体更新速率,从而减弱了该效应,而5 月份的径流在输送DOC 过程中的作用并不突出也印证了该推论.部分DOC 可能蓄积在沉积物中,另一些则可能作为无机碳排放到湖外.洞庭湖9 月DOC 和a254的蓄积效果最好,其次是11 月,由此可知9 月上游DOM 可以很好地转运到下游区域.
4 结论
4.1 采样期间,洞庭湖流域输入河流DOC 浓度高于唯一出湖口城陵矶,且在径流量较大的5 月和7 月表现得更为明显.
4.2 各输入河流中DOM 丰度存在较大的空间差异.类腐殖质物质多见于长江荆南松滋、虎渡、藕池三口,而类蛋白质物质多见于湖南湘、资、沅、澧四水.
4.3 生物培养实验表明,微生物对输入河流不同荧光组分的利用具有选择性,腐殖质样蛋白和类蛋白质的荧光强度均被微生物有效利用而减弱.
4.4 湖泊9 月径流较小,湖泊水力滞留时间相对较长,而7月的较大径流量增强了外源DOM输入,而不同水力连通条件下DOM 迁移转化的特征及环境效应则有待于进一步研究.
猜你喜欢
安徽文学(2023年4期)2023-04-11 22:49:27
小学生学习指导(低年级)(2021年10期)2021-11-01 08:22:58
金桥(2020年8期)2020-05-22 06:22:54
中国环境科学(2018年10期)2018-10-29 04:22:40
水利规划与设计(2017年5期)2017-06-09 08:56:23
音乐天地(音乐创作版)(2016年4期)2016-07-28 09:39:37
林业与生态(2016年3期)2016-02-27 14:24:22
水土保持通报(2014年5期)2014-06-09 08:27:00
水土保持通报(2014年5期)2014-06-09 08:26:46
湖南水利水电(2014年3期)2014-02-27 14:46:05
