
取食不同烤烟品种对斜纹夜蛾幼虫体内保护酶和解毒酶活性的影响
2024-03-25 10:38李勇辉雷婷徐元昌赵明富严乃胜柳青唐国文
江苏农业科学 2024年2期
关键词:烤烟
李勇辉 雷婷 徐元昌 赵明富 严乃胜 柳青 唐国文
摘要: 明确取食不同烤烟品种对斜纹夜蛾(Spodoptera litura)幼虫体内保护酶和解毒酶活性的影响。在室内分别用5种烤烟品种叶片(云烟87、0790A、GH、TW48、TW53)饲养斜纹夜蛾,测定其5龄幼虫体内保护酶(CAT、POD、SOD)及解毒酶(Car E、GSTs、CYP450)活性,并分析不同烤烟品种次生代谢物及营养物质含量与斜纹夜蛾幼虫体内酶活性的相关性。结果表明,5个烤烟品种中TW48叶片可溶性蛋白含量最低,0790A叶片单宁、总酚和类黄酮含量均为最高;其中单宁含量是TW53叶片的1.60倍,类黄酮含量是 GH叶片的1.37倍。取5个烤烟品种的叶片喂养斜纹夜蛾幼虫至5龄后体内CAT、POD、SOD、Car E、GSTs活性存在显著差异(P<0.05),其中取食烤烟品种0790A叶片后体内CAT、POD和SOD活性最高,取食烤烟品种TW48叶片后体内CAT、POD、Car E、GSTs活性最低,而取食不同烤烟品种叶片后的幼虫CYP450活性则无显著性差异。可见,取食次生物质含量不同的烤烟品种叶片物会使斜纹夜蛾体内保护酶和解毒酶活性发生变化。
关键词: 斜纹夜蛾;烤烟;次生代谢物物质;保护酶;解毒酶
中图分类号:S435.72 文献标志码:A
文章编号:1002-1302(2024)02-0131-05
斜纹夜蛾(Spodoptera litura)为鳞翅目(Lepidoptera)夜蛾科(Noctuidae)斜纹夜蛾属,是一种重要的农作物害虫[1],主要分布于温带和亚热带地区[2],具有寄主范围广、繁殖力强、适生范围广和危害严重等特点[1-3]。烟草作为我国的主要经济作物,种植面积和产量均位于世界第一[4]。全球约有800多种害虫会对烟草进行危害,其中斜纹夜蛾对烟草的危害最为致命[5],特别是幼虫进入过量摄食期后,会对整株烟草进行危害, 使烟草的市场价值大幅降低,造成巨大的经济损失[6]。然而,随着国家对生态文明建设的高度重视及人们对农产品安全要求的提高,化学防治带来的问题越来越突出,相比之下,筛选优质的烟草种质资源,利用烟草自身的抗虫性,已经成为有效防治斜纹夜蛾的新途径之一[7]。
次生代谢物在植物抗虫性中占有重要地位,是构成植物抗虫性的主要化学基础[8-9]。次生代谢物作为启动昆虫体内相关解毒酶系的关键因子,会显著改变害虫对药剂的敏感性,影响其田间防治效果,同时昆虫体内解毒酶活性的变化也为其适应环境提供了更多的可能性[10]。相关研究也表明,昆虫体内相关酶活性的变化,会受到其取食寄主植物的次生代谢物和营养物质含量的影响[11-14]。近年来国内外关于不同寄主植物对植食性昆虫体内酶活性影响的研究,主要集中在不同属植物对某种昆虫酶活性的影响上,如张林林等对小地老虎取食3种寄主植物后体内保护酶活性进行了研究,结果表明取食白菜和玉米的幼虫体内过氧化物酶(POD)和超氧化物歧化酶(SOD)活性均显著低于取食大豆的幼虫[12];李时荣等研究发现1代麦长管蚜在取食大麦后体内乙酰胆碱酯酶(AchE)和谷胱甘肽S-转移酶(GSTs)活性的诱导效果最显著[13];Jin等分析了棉铃虫幼虫取食不同宿主植物(棉花、玉米、大豆和辣椒)后体内酶活性变化情况,结果表明取食4种宿主植物后幼虫体内解毒酶活性存在显著差异[14]。但是,目前对于取食同种植物的不同品种对植食性昆虫酶活性的影响研究,鲜见报道。
因此,本研究选取烤烟选育品种0790A和GH14-102,地方主栽品种云烟87,野生种TW48和TW53的叶片连续喂养斜纹夜蛾至5龄后,测定了取食不同烤烟品种叶片后斜纹夜蛾幼虫体内酶活性的变化情况,并研究了烤烟品种叶片内营养物质及次生代谢物含量与2类酶活性的关系,旨在明确斜纹夜蛾对不同烤烟品种的适应机制,也为下一步烤烟品种选育研究提供一定的参考。
1 材料与方法
1.1 供試虫源及寄主植物
斜纹夜蛾于2021年5月在云南省玉溪市元江县(101°58′E,23°35′N,海拔为421 m)采集,将采集到的昆虫放入人工气候室内进行饲养[光—暗周期为16 h—8 h;相对湿度为(70±10)%;温度为(26±1) ℃]。等待成虫羽化结束,按雌雄对比 1 ∶ 1 的比例饲喂,试验用虫来自同一批雌虫产下的卵。
供试烤烟种子由云南省烟草农业科学研究院提供,地方性品种为云烟87,选育品种为0790A、GH14-102(以下简称GH),野生种为TW48和TW53。种植前,对种植所用的土壤采用立式压力蒸汽灭菌锅进行消毒灭菌。使用花盆室内种植,并置于300目纱笼(1.6 m×0.9 m×1.1 m)中,花盆所处位置每隔48 h更换1次,随机摆放。烟草生长阶段不打农药、不施化肥,定期浇水,及时清除杂草。
1.2 试验方法
1.2.1 试虫饲养 将初孵幼虫放入玻璃管(直径为2.6 cm,高度为8 cm)中进行单头饲养,用纱网封口(150目),在管壁上编号,放入人工气候室[光—暗周期为16 h—8 h;湿度为(70±10)%;温度为(26±1) ℃]进行饲养。每个烤烟品种饲养 30 头幼虫,每天09:00和21:00准时观察并记录幼虫龄期,将进入5龄的幼虫用液氮迅速冷冻,放入冰箱(-80 ℃)存储,以备测定其体内相关酶活性。
1.2.2 斜纹夜蛾体内酶活性的测定 酶液提取:取1头 5 龄供试幼虫,用 4 ℃ 的2~5 mg 生理盐水冲洗,滤纸拭干,称取幼虫体质量,放入离心管(2 mL)中。适量加入预冷匀浆介质,置于高通量研磨仪中粉碎,4 ℃环境中进行600 s离心处理,待离心结束,取上清液为待测液。
酶活性测定:采用南京建成生物工程研究所的试剂盒测定过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)、羧酸酯酶(Car E)和谷胱甘肽S-转移酶(GSTs)活性;上海优选生物科技有限公司的试剂盒测定细胞色素P450(CYP 450)活性。
1.2.3 寄主植物叶片次生物质含量的测定 将“1.1”节中用于饲喂斜纹夜蛾的5种烤烟品种叶片进行收集,按照试剂盒说明书对叶片的营养物质(可溶性蛋白、可溶性糖)和次生代谢物(单宁、总酚、类黄酮)含量进行测定及计算。试剂盒来源:南京建成生物工程研究所(用于测定可溶性糖、可溶性蛋白、总酚和类黄酮含量);苏州格瑞思生物科技有限公司(用于测定单宁含量)。
1.3 数据处理
2 类酶活性和寄主植物次生代谢物、营养物质含量数据的分析均在 Excel 2019 和 SPSS 23.0 软件中进行处理。应用 Duncans 法(α=0.05) 对比不同处理间的差异显著性,用 Pearson 线性相关系数对斜纹夜蛾 5 龄幼虫体内酶活性与寄主植物次生代谢物、营养物质含量的相关性进行分析(P<0.01)。
2 结果与分析
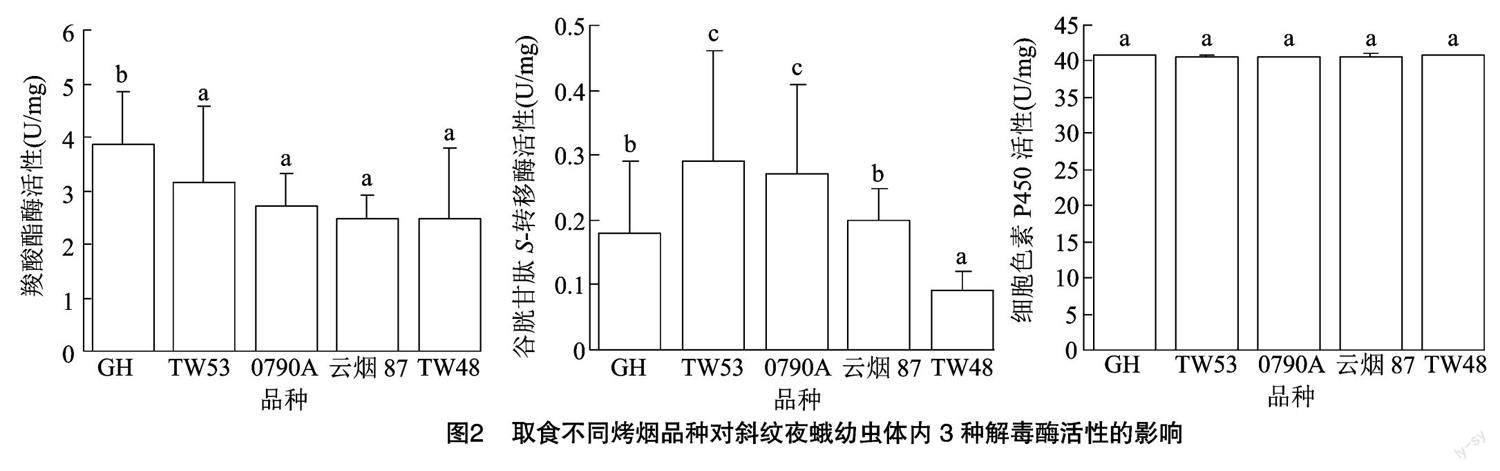
2.1 5种烤烟品种叶片内营养物质及次生代谢物质含量分析
研究表明,不同烤烟品种叶片内的营养物质含量存在一定差异。可溶性蛋白含量为 GH>云烟 87>0790A>TW53>TW48,其中GH可溶性蛋白含量显著高于0790A、TW53和TW48(P<0.05);可溶性糖含量为 TW48>0790A>TW53>云烟87>GH,其中TW48的可溶性糖含量[(6.64±1.63) mg/g]是GH[(4.14±0.45) mg/g]的1.60倍(表1)。
5种烤烟品种叶片中单宁含量为 0790A>GH> TW48>云烟87>TW53,其中 0790A 单宁含量[(31.60±7.83) mg/g]是TW53[(19.78±5.24) mg/g] 的1.60倍;总酚含量为 0790A>GH>云烟87>TW53>TW48,其中0790A烤烟品种的总酚含量[(654.33±42.42) μmol/g]显著高于其他4个烤烟品种,分别是TW48、TW53、云烟87和GH的3.78、2.46、1.59和1.54倍;类黄酮含量为 0790A>云烟87>TW53>TW48>GH,其中0790A的类黄酮含量[(3.12±0.12) mg/g]是GH[(2.28±0.50) mg/g]的1.37倍(表1)。
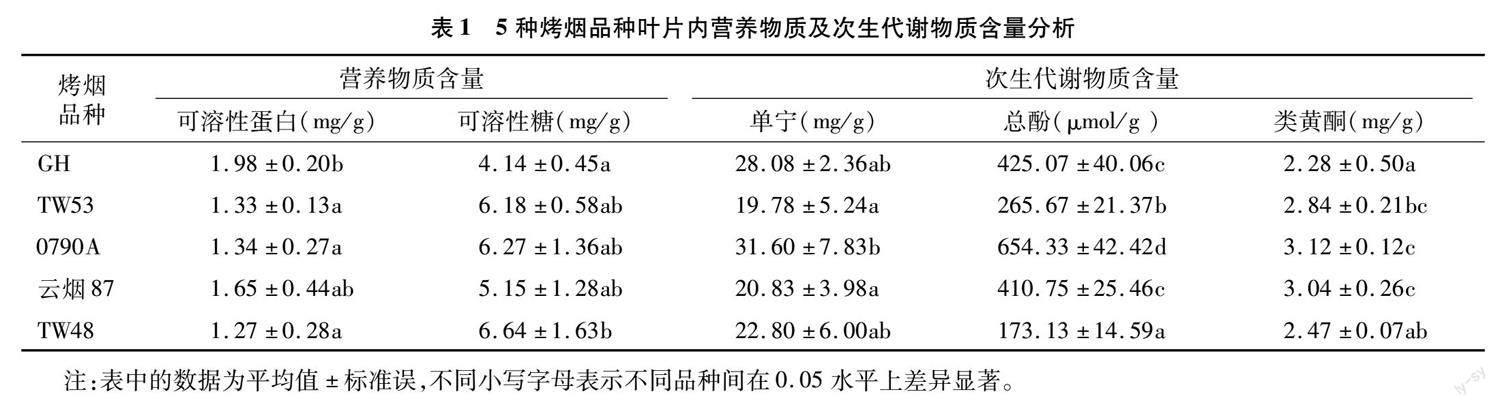
2.2 取食不同烤烟品种对斜纹夜蛾幼虫体内保护酶活性的影响
斜纹夜蛾幼虫取食5个烤烟品种叶片至5龄后, 体内3种保护酶活性变化情况存在差异(图1)。CAT活性为 0790A(23.75 U/mg)>TW53(20.67 U/mg)> 云烟87(15.04 U/mg)>GH(13.01 U/mg)>TW48(3.95 U/mg),其中取食0790A和TW53烤烟叶片的斜纹夜蛾幼虫体内酶活性显著高于其他3个品种(P<0.05)。POD活性为0790A(10.12 U/mg)>云烟87(8.67 U/mg)>TW53(4.84 U/mg)>GH(3.16 U/mg)>TW48(1.43 U/mg),取食0790A和云烟87 烤烟叶片的斜纹夜蛾幼虫体内酶活性与取食其他3个烤烟品种的幼虫POD活性间存在显著差异(P<0.05)。SOD活性为0790A(30.24 U/mg)> TW53(24.24 U/mg)>云烟87(19.71 U/mg)>TW48(15.06 U/mg)>GH(14.75 U/mg),其中取食0790A叶片的幼虫体内SOD活性显著高于取食TW48、云烟87和GH,取食GH和TW48相比,体内SOD活性无显著差异(P>0.05)。
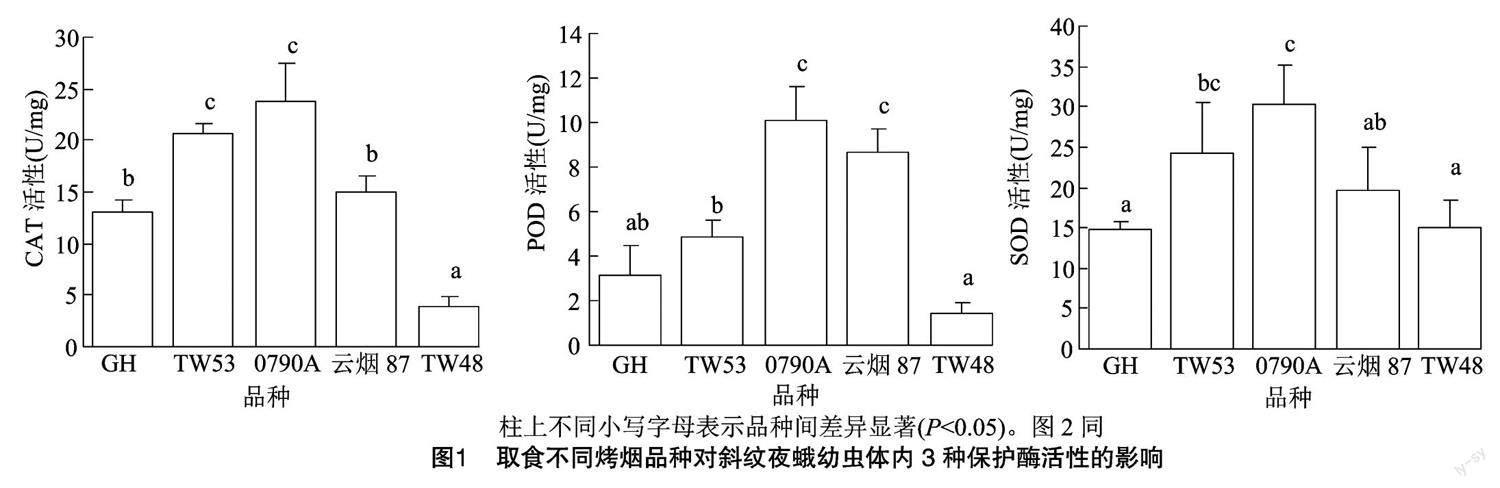
2.3 取食不同烤烟品种对斜纹夜蛾幼虫体内解毒酶活性的影响
分别对斜纹夜蛾幼虫喂食不同品种叶片至5龄后,测定其體内的解毒酶活性。结果表明,取食不同品种叶片幼虫体内GSTs、Car E 活性均存在显著差异,CYP450活性则无显著差异(图2)。Car E 活性为 GH(3.88 U/mg)>TW53(3.16 U/mg)>0790A(2.71 U/mg)>云烟87(2.48 U/mg)>TW48(2.46 U/mg),其中取食烤烟品种GH叶片的斜纹夜蛾幼虫体内Car E活性显著高于其他4个烤烟品种;GSTs活性为 TW53(0.29 U/mg)>0790A(0.27 U/mg)>云烟87(0.20 U/mg)>GH(0.18 U/mg)>TW48(0.09 U/mg),取食烤烟品种 TW53 和 0790A叶片的斜纹夜蛾幼虫体内GSTs活性显著高于其他3个品种(P<0.05)。
2.4 斜纹夜蛾幼虫体内酶活性与烤烟品种叶片内营养物质及次生代谢物质含量相关性分析
经皮尔森相关分析,结果(表2)表明,斜纹夜蛾幼虫体内CAT活性与总酚和类黄酮含量均 表现为正相关,与单宁含量呈负相关,正相关关系中 r类黄酮> r可溶性糖>r总酚>r可溶性蛋白(r为相关系数绝对值,下同);POD活性仅与类黄酮含量存在正相关关系,其余均呈负相关,表现为r总酚>r单宁>r可溶性糖>r可溶性蛋白;SOD活性与2种营养物质含量存在较高的正相关关系,表现为r可溶性蛋白>r可溶性糖,与3种次生代谢物质含量均呈负相关, 其中与类黄酮含量的相关性最高;Car E活性与2种营养物质和单宁含量均呈正相关,与总酚含量呈负相关,与类黄酮含量呈极显著负相关(P<0.01),正相关关系中r可溶性蛋白>r单宁>r可溶性糖;GSTS 除了和可溶性 蛋白呈负相关外,其余均呈正相关,表现为r类黄酮>r总酚> r可溶性糖>r单宁;CYP450与2种营养物质呈正相关, 与3种次生代谢物质呈负相关,正相关中r可溶性蛋白> r可溶性糖;负相关中r类黄酮>r总酚>r单宁。
3 讨论与结论
在漫長的进化历程中,昆虫与寄主植物逐渐形成了复杂的协同进化关系[15]。一方面,昆虫以寄主植物为食,为其生长、发育和繁殖获取能量[16],同时寄主植物也能产生对昆虫有害的抗虫物质,抵御昆虫的取食[16-17]。昆虫体内的解毒系统是昆虫适应抗虫物质的重要代谢机制,在受到有害物质侵扰后会诱导其体内以保护酶和解毒酶为主的酶活性发生改变,从而增强昆虫对抗虫物质的代谢能力,使其能维持正常的生命活动[18-19]。斜纹夜蛾适生范围广,繁殖力强,对烤烟危害较大。研究斜纹夜蛾取食不同烤烟品种叶片后幼虫体内酶活性变化情况对烟田抗虫品种的选育及害虫综合防治具有很好的借鉴意义。
植物体内营养物质、次生代谢物的种类及含量与植物抗虫性密切相关[20],如Vanitha等对抗虫和感虫水稻品种叶片中部分化合物含量进行了研究,发现抗虫水稻品种叶片内次生代谢物含量显著高于感虫水稻品种,而感虫品种叶片内糖含量、粗蛋白和总游离氨基酸的含量则高于抗性品种[21-22];不同地区的红松球果单宁、黄酮和总酚含量不同,其受到球果尺蛾 (Eupithe ciaabietaria) 和果梢斑螟 (Dioryctria pryeri) 的危害率与3种次生代谢物含量呈负相关关系[23]。贾尊尊等研究结果也证实了总蛋白和脯氨酸含量(次生代谢物) 的变化是随着危害密度的升高而升高,而可溶性糖(营养物质)含量则是随着危害密度升高而降低[24]。结合本研究结果,5个品种中 0790A 单宁、 总酚和类黄酮含量最高,可溶性蛋白含量较低;TW48的可溶性糖含量最高,总酚含量最低,说明在一定程度 0790A 烤烟具有较强的抗虫性特征,而 TW48 则具有一定的感虫特征。
昆虫在受到次生代谢物的侵扰后,体内会产生大量的超氧离子自由基,从而改变其身体中的保护酶活性[25-26]。SOD是昆虫体内重要的防御机制,能使昆虫体内的自由基发生歧化反应,CAT 能将过量的 H2O2 转化为 H2O 和 O2,POD 在有毒物质的清除及活性氧的代谢过程中发挥着重要作用[27]。鲁智慧等研究表明,草地贪夜蛾取食总酚、单宁和类黄酮含量较高的马铃薯叶片与取食总酚、单宁和类黄酮含量较低的马铃薯叶片相比,其幼虫体内 POD、SOD 和 CAT 活性均表现出显著升高的趋势[26]。本研究发现,0790A 烤烟叶片的总酚、单宁和类黄酮含量显著高于其他4个烤烟品种,且取食该叶片的斜纹夜蛾幼虫体内 CAT、POD 和 SOD 活性水平均较高,说明斜纹夜蛾幼虫体内保护酶被 0790A 烤烟叶片内次生代谢物质激活,从而降低次生代谢物的毒害作用,维持正常的生理代谢。
植食性昆虫会形成独特的解毒代谢机制[28],分解植物体内的外源有毒物质和代谢物质,主要有羧酸酯酶、谷胱甘肽S-转移酶和细胞色素P450等,这些酶会随着取食寄主植物的不同而发生变化,如亚洲小车蝗解毒酶(CYP450s,CarEs,GSTs)活性在摄入次级代谢产物含量高的牧草后也更高[16],而臭椿在取食总酚含量最高的寄主植物后,其体内的 CarE 和 GSTs 活性则较低[29]。定量摄入单宁、芸香苷及没食子酸后,斜纹夜蛾体内羧酸酯酶活性得到显著提升[30]。在本研究中,取食总酚和单宁含量最高的烤烟品种叶片(0790A)后斜纹夜蛾幼虫体内GSTs活性较高;取食总酚和单宁含量较高的烤烟品种叶片(GH)后斜纹夜蛾幼虫体内Car E活性也处于较高水平。而取食类黄酮含量较高的烤烟品种(云烟87),其体内 Car E 和 GSTs 活性并不高,出现这种情况的原因可能是由于取食该品种烤烟后,斜纹夜蛾幼虫体内保护酶活性特别是 POD 较高,减轻了类黄酮的毒害作用。取食不同烤烟品种叶片的斜纹夜蛾幼虫体内 CYP 450 活性变化不显著,这可能是由于斜纹夜蛾对不同烤烟品种的适应可塑性较强,经过5龄长期以同一寄主植物为食,逐渐适应了这些寄主[31]。
本研究结果表明,斜纹夜蛾对不同烤烟品种表现出一定程度的适应可塑性,体内2类酶活协同发挥作用,充分利用营养物质含量不同的烤烟,调节保护酶和解毒酶活性,降低虫体自身能量的消耗,从而进一步提升对寄主植物次生代谢物和有毒物质的适应性。本研究仅通过营养物质、次生代谢物质和2类酶活性初步探索斜纹夜蛾对不同烤烟品种的适用机制,对于其他酶活性和分子适应机制有待进一步探究。
参考文献:
[1] Hou W T,Staehelin C,Elzaki M E A,et al. Functional analysis of CYP6AE68,a cytochrome P450 gene associated with indoxacarb resistance in Spodoptera litura (Lepidoptera:Noctuidae)[J]. Pesticide Biochemistry and Physiology,2021,178:104946.
[2]Fong C J,Chuang Y Y,Lai H Y. Effects of amendment with various vermicomposts on the soil fertility,growth of Brassica chinensis L. and resistance of Spodoptera litura Fabricius larvae[J]. Sustainability,2021,13(16):9441.
[3]赵胜园,罗倩明,孙小旭,等. 草地贪夜蛾与斜纹夜蛾的形态特征和生物学习性比较[J]. 中国植保导刊,2019,39(5):26-35.
[4]陈丽鹃,陈 闺,周冀衡,等. 烟田秸秆腐解特性及其腐解液对烟草疫霉菌的影响[J]. 中国烟草科学,2021,42(1):33-39.
[5]李庆亮,张 佳,宗 浩,等. 烟草抗虫机制研究进展[J]. 农学学报,2017,7(8):48-54.
[6]Chen J Y,Li X H,Li Y Y,et al. Effect of fertilization level on the tobacco nutritional quality and the development,nutritional indices,fecundity of Spodoptera litura (F.)[J]. Entomology,Ornithology & Herpetology:Current Research,2021,10(6):1-7.
[7]李晓红,黄志友,杨贤均,等. 不同品种烟草对斜纹夜蛾生活史和产卵选择行为的影响[J]. 植物保护,2021,47(1):123-127,152.
[8]Yuan G G,Zhao L C,Du Y W,et al. Repellence or attraction:secondary metabolites in pepper mediate attraction and defense against Spodoptera litura[J]. Pest Management Science,2022,78(11):4859-4870.
[9]Bai C M,Liu Y F,Lu X C. The application of secondary metabolites in the study of sorghum insect resistance[J]. IOP Conference Series:Earth and Environmental Science,2018,128:012169.
[10] Yan J X,Tan Y,Lyu Y R,et al. Effect of jasmonate treatments on leaves of Rosa rugosa ‘Plena and detoxification enzymes and feeding of adult Monolepta hieroglyphica[J]. Journal of Forestry Research,2021,32(3):1253-1261.
[11]鲁智慧,和淑琪,郭 晋,等. 取食不同寄主植物对草地贪夜蛾幼虫体内3类酶活性的影响[J]. 南方农业学报,2020,51(10):2461-2469.
[12]张林林,李艳红,仵均祥. 不同寄主植物对小地老虎生长发育和保护酶活性的影响[J]. 应用昆虫学报,2013,50(4):1049-1054.
[13] 李时荣,葛朝虹,刘德广,等. 寄主植物对不同基因型麦长管蚜解毒酶活性的影响[J]. 西北农业学报, 2018,27(2):283-293.
[14]Jin M,Liao C,Fu X,et al. Adaptive regulation of detoxification enzymes in Helicoverpa armigera to different host plants[J]. Insect Molecular Biology,2019,28(5):628-636.
[15]沈海燕,梁 运,陈正浩,等. 不同寄主植物对云南松毛虫生长发育、繁殖及生理酶活性的影响[J]. 林业科学研究,2022,35(5):63-70.
[16]陈德霞,刘 旭,罗林华,等. 典型草原4种优势种牧草次生代谢物对亚洲小车蝗解毒酶活性的影响[J]. 草业学报,2020,29(1):183-192.
[17]郝江宁,常 静,李 妍,等. 油菜叶片性状及次生代谢物质与其对黄宽条跳甲抗性的关系及对跳甲解毒酶活性的影响[J]. 植物保护,2022,48(4):158-164.
[18]陈澄宇,康志娇,史雪岩,等. 昆虫对植物次生物质的代谢适应机制及其对昆虫抗药性的意义[J]. 昆虫学报,2015,58(10):1126-1139.
[19]杨金睿,肖关丽. 植物抗虫生理研究进展[J]. 中国农学通报,2021,37(6):130-136.
[20]Afroz M,Rahman M M,Amin M R,et al. Insect-plant interaction with reference to secondary metabolites:a review[J]. Agricultural Reviews,2021,42(4):427-433.
[21]Vanitha B,Kumar C A,Kumar L,et al. Association of biochemical factors in rice against infestation of rice leaf folder,Cnaphalocrocis medinalis (Guenee) (Lepidoptera:Pyralidae)[J]. Journal of Experimental Zoology,2015,18:373-376.
[22] 王麗艳,金永玲,朱 莹,等. 水稻负泥虫对不同水稻品种次生物质的行为反应[J]. 植物保护,2016,42(4):114-118.
[23] 左彤彤,李兴鹏,周春艳,等. 不同地理种源红松抗虫性及其与次生代谢产物相关性[J]. 中国森林病虫,2019,38(2):22-25.
[24]贾尊尊,付开赟,丁新华,等. 烟粉虱持续取食对棉花叶片防御酶活性、营养物质和代谢产物含量的影响[J]. 新疆农业科学,2022,59(4):916-924.
[25]贾变桃,焦 鹏,杨素梅. 虱螨脲亚致死浓度对小菜蛾保护酶系和解毒酶系活力的影响[J]. 植物保护学报,2016,43(2):293-299.
[26]鲁智慧,陈亚平,周叆春,等. 不同寄主植物继代饲养后草地贪夜蛾幼虫生理酶活性差异[J]. 环境昆虫学报,2020,42(6):1361-1368.
[27]Piskorski R,Dorn S.How the oligophage codling moth Cydia pomonella survives on walnut despite its secondary metabolite juglone[J]. Journal of Insect Physiology,2011,57(6):744-750.
[28]Chen Y Z,Zhang B W,Yang J,et al. Detoxification,antioxidant,and digestive enzyme activities and gene expression analysis of Lymantria dispar larvae under carvacrol[J]. Journal of Asia-Pacific Entomology,2021,24(1):208-216.
[29]李路莎,袁郁斐,武 磊,等. 不同寄主植物对美国白蛾幼虫取食行为及解毒酶活性的影响[J]. 昆虫学报,2018,61(2):232-239.
[30]黄敏燕,李雪峰. 植物次生物质对斜纹夜蛾解毒酶活性的影响[J]. 基因组学与应用生物学,2018,37(8):3495-3502.
[31]Rivero A,Magaud A,Nicot A,et al. Energetic cost of insecticide resistance in Culex pipiens mosquitoes[J]. Journal of Medical Entomology,2011,48 (3):694-700.
收 稿日期:2023-02-20
基金项目: 中国烟草总公司科技项目[编号:110202101044(JY-21)];中国烟草总公司云南省公司科技项目(编号:2022530000241010)。
作者簡介:李勇辉(1999—),男,云南楚雄人,硕士研究生,研究方向为害虫生物防治方面。E-mail:liyonghui0705@163.com。
通信作者:唐国文,博士,副教授,主要从事农业害虫研究,E-mail:guowen03@163.com;柳 青,博士,副教授,主要从事农业害虫研究,E-mail:liuqinggc065@126.com。
猜你喜欢
湖北农机化(2020年4期)2020-07-24
活力(2019年19期)2020-01-06
中国烟草学报(2017年1期)2017-11-16
中国烟草学报(2017年1期)2017-11-16
作物研究(2014年6期)2014-03-01
湖南农业科学(2014年18期)2014-02-27
湖南农业科学(2014年18期)2014-02-27
湖南农业科学(2014年8期)2014-02-27
中国烟草学报(2012年3期)2012-04-10
中国烟草学报(2012年1期)2012-04-09
