
菌藻共生系统污水处理及CO2固定作用机制的研究进展
2024-03-22 09:47吴鑫明操家顺刘伟京
净水技术 2024年3期
安 浩,吴鑫明,操家顺,李 超,*,刘伟京
(1.中国五冶集团有限公司,四川成都 610000;2.河海大学环境学院,江苏南京 210098;3.河海大学浅水湖泊综合治理与资源开发教育部重点实验室,江苏南京 210098;4.江苏省环境科学研究院,江苏南京 210036)
近年来,随着人类生产及生活活动的增加,工农业废水及生活污水大量排放,严重污染了水生态环境,极易引起水体富营养化[1]。目前,许多生物处理技术已经被广泛应用到了市政污水的处理中,包括传统活性污泥(CAS)技术[2]、人工湿地技术[3]、植物修复技术[4]、生物膜技术[5]和微生物燃料电池技术[6]等。此外,高级氧化技术也可实现对有机物和氮磷的高效去除[7]。虽然这些工艺取得了良好的污染物去除效果,但它们仍存在能源消耗大和温室气体(例如:CO2、CH4)排放等缺点,不满足当前低碳发展的要求。因此,开发既能实现污水处理同时又能减少温室气体排放的处理技术迫在眉睫。
菌藻共生技术作为一种能实现同步处理污水和固定CO2的绿色处理技术引起了越来越多的关注[8]。细菌和微藻通过二者间的相互作用和协同增效来提高污水处理过程中的污染物去除率(图1)。微藻能有效同化CO2同时产生O2来代替外部曝气,而O2又被细菌用来氧化有机物产生CO2。在这个过程中,光作为菌藻系统处理废水的能量来源,可以减少曝气电能消耗,并且光合作用吸收CO2可以减少温室气体排放[9]。微藻吸收污水中氮磷等污染物并将其转化为生物质,细菌自身代谢产生信号分子、微量元素和无机碳源等供微藻生长。微藻被证明可以和硝化细菌的相互作用来改变氮转化效率[10-11],并影响磷酸盐吸收的微生物群落丰度[12]。作为一种绿色工艺,菌藻共生技术为未来的城市污水处理实现“碳达峰”和“碳中和”目标提供了可能。然而,目前研究都集中在菌藻共生系统处理废水和固定CO2的菌种和藻种筛选上,重点关注菌藻共生系统对污染物的去除效率和CO2的固定效率:Xu等[13]研究了微藻与内生细菌(S395-2)共生系统净化沼气,当菌藻比例为10∶1时取得最佳净化效果,COD、氮、磷和CO2的去除效率分别为88.29%±5.03%、88.31%±4.29%、88.21%±4.51%和68.13%±1.69%。这忽略了细菌和微藻在处理废水和固定CO2过程中二者相互作用机理的研究。
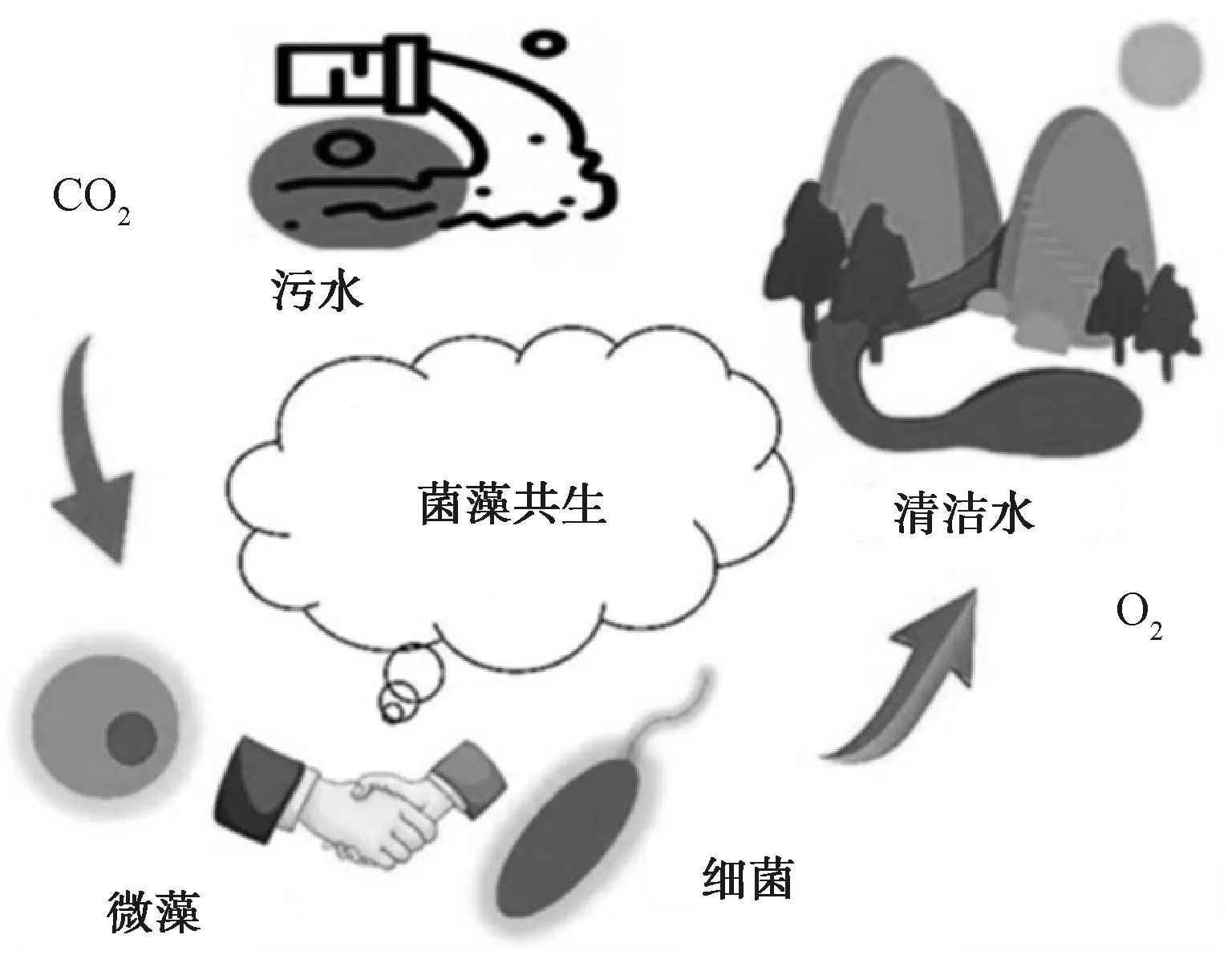
图1 菌藻共生系统净水固碳作用
本文综述了菌藻共生系统在减污固碳过程中的作用效能,并从脱氮除磷以及CO2固定的角度阐述了菌藻共生系统的相互作用机制,最后对菌藻共生体系实际应用中的现存问题进行了总结,并对未来的研究方向提出了建议,为今后菌藻共生体系的工程应用提供了理论支持。
1 菌藻共生系统去除氮
1.1 菌藻共生系统去除氨氮


图2 菌藻共生系统脱氮机理
硝化作用并不是菌藻共生系统中去除氨氮的唯一途径。微藻和部分异养细菌将氨氮作为营养物质吸收,然后将其同化为细胞组分,从而实现水中氨氮的去除。氨氮通过微藻细胞内产生的转运器转运到微藻细胞中,在谷氨酰胺合成酶和谷氨酸脱氢酶等酶的作用下,氨氮可以很容易地被同化为氨基酸,成为微藻生物质的一部分[17]。Bankston等[12]发现,通过微藻同化作用去除的氨氮几乎是硝化作用去除的2倍。细菌可以促进微藻生物质吸收同化较高浓度的氨氮。Bankston等[8]发现,投加活性污泥后的菌藻共生系统与微藻单独作用时相比,既可以促进微藻对氨氮的同化吸收,还能使微藻保持较高的生长速率。
水中氨氮浓度是影响菌藻间作用关系的重要参数。在低氨氮浓度下,由于缺乏氨氮,微藻和硝化细菌之间为竞争关系,二者相互释放具有抑制作用的物质(例如藻毒素、植物激素等),对微藻和细菌间关系产生负面影响[18]。Choi等[19]在一项研究中发现,当水中氨氮含量不足,微藻的存在抑制了菌群77%的硝化作用。当氨氮浓度高时,可供硝化细菌反应和微藻同化的氮源充足,微藻和硝化细菌之间是互利共生关系。Su等[20]发现,在氨氮含量充足的条件下,小球藻和细菌共同培养与藻类、细菌分别单独培养相比,氨氮的去除率明显提高。
1.2 菌藻共生系统去除硝态氮
反硝化过程主要发生在缺氧条件下,但在菌藻共生系统中,微藻通过光合作用产生O2会抑制反硝化过程相关酶的活性,进而影响反硝化进程[20]。Petrovi等[24]发现,菌藻共生系统中反硝化过程的反应速率低于反硝化细菌单独作用时的反应速率。此外,在菌藻共生系统中,夜间藻类呼吸会消耗大量的DO[25],从而进一步强化了反硝化过程所需的厌氧环境。白天微藻通过光合作用制造O2提高DO含量,有助于硝化作用的发生;黑暗条件下微藻通过呼吸代谢进一步消耗DO,为反硝化作用创造了良好的缺氧环境。废水中有机碳源的存在有助于黑暗条件下微藻和细菌呼吸代谢进一步消耗废水中DO,为反硝化过程构建缺氧环境。Rezvani等[26]证明,菌藻共生体系中微藻可以利用光照期间产生的O2,在黑暗条件下降解废水中有机物。
菌藻间的协同作用可以实现二者代谢产物的交换,以满足二者各自的营养需求[27]。例如,微藻光合作用产生的有机物可以为低有机负荷废水中的细菌提供有机碳源,细菌反过来可以通过自身代谢活动降解微藻产生的有机物,抑制废水中COD含量的增长[28]。另外,某些微藻可以为自养反硝化细菌在硝酸盐还原过程中提供其所需的氢气。微藻黑暗条件下产生氢气,为反硝化的进行提供物质保障;在白天,微藻通过光合作用重建碳储备,为后期氢气的产生提供动力。Rezvani等[29]比较了耗氢脱硝器(HCD)-菌藻共生系统与HCD-单独细菌作用下的硝酸盐去除效率,在不注入外部氢气的情况下,HCD-菌藻共生系统具有更高的硝酸盐去除率。菌藻间可以通过群体感应(QS)信号分子实现胞间通讯,从而调节各种生命活动与代谢过程。Ji等[30]研究发现,地衣芽孢杆菌产生的一种信号分子AIP可被QS受体感知,细菌细胞内的c-di-GMP受体接收并放大信号,诱导细菌产生某些物质,刺激普通小球藻细胞中叶绿素代谢相关基因的表达。结果发现,共生系统中的叶绿素a浓度随着干细胞重量的显著增加而增加。当初始菌藻比例为3∶1时,共生系统的相互作用和协同效应最为明显,COD、TP、TN的去除率分别为86.55%、80.28%、88.95%,此时c-di-GMP的质量分数达到了3.85 μg/g,AIP的质量浓度达到了2.81 mg/L。
2 菌藻共生系统去除磷


图3 菌藻共生系统除磷机理
聚磷菌是污水处理厂中起主要作用的除磷菌[32]。聚磷菌在厌氧-好氧交替条件下将废水中吸收的正磷酸盐以多磷酸盐的形式储存在细胞内[33],在厌氧条件下,聚磷菌利用聚磷酸盐水解产生的能量吸收废水中的碳源,并以聚羟基烷酸(PHAs)和聚羟基丁酸(PHBs)的形式储存在细胞内,同时水解产生的正磷酸盐释放到水中。在好氧条件下,聚磷菌利用细胞内的PHAs和PHBs为碳源,过量吸收废水中的正磷酸盐合成聚磷酸盐储存在细胞内,将废水中的磷转移到聚磷菌体内。最终,聚磷菌体内的聚磷酸盐随着剩余污泥的排放而去除[34-35]。微藻可以与聚磷菌相互作用,促进对磷的去除,甚至微藻可以直接吸收积累聚磷酸盐[36]。Ji等[37]研究了非曝气菌藻颗粒污泥工艺中磷的去除,结果表明,进水中近86%的磷酸盐在6 h内被去除,其中包括在黑暗条件下反应2 h和光照条件下反应4 h,观察到菌藻颗粒污泥在黑暗阶段有少量的磷释放,在光照条件下废水中的聚磷酸盐被大量吸收。
微藻还可以通过同化作用从废水中去除磷酸盐。正磷酸盐被吸附在细胞表面[38],然后被运送进细胞内同化与供细胞生长[39]。根据Rezvani等[29]的一项研究,微藻对磷酸盐的同化是菌藻共生系统中磷酸盐去除的主要机制。磷是微藻光合作用所需的重要元素,其平均含量占微藻干重的1%[18],磷的缺乏会降低光合作用卡尔文循环中底物的合成和再生速率,从而抑制微藻光合作用的进行[18]。菌藻共生系统是提高除磷效率的一种可持续方法。Rezvani等[29]表明,与单一微藻[4.5 mg/(L·d)]和细菌[2.6 mg/(L·d)]除磷效率相比,菌藻共生系统[6.34 mg/(L·d)]拥有更高的磷酸盐去除率。菌藻共生系统的同化率主要由藻类生长率决定,并且细菌的存在可以促进微藻的生长。细菌可以通过提供维生素[40]和植物激素[41](例如,生长素和发光色素)来促进微藻的生长。细菌提供的维生素等辅助因子已被证明可以改善藻类生长和废水养分吸收[40],例如,生长素可以促进植物和微藻细胞生长[42],同时抑制氧化应激反应[43]。此外﹐Liu等[44]在研究菌-藻生物膜EPS时发现,由于菌-藻生物膜表面含有蛋白质和多聚糖等组分﹐其具有丰富的含氧官能团,有机磷可以与EPS发生配位作用结合,通过剩余污泥沉淀去除。
3 菌藻共生系统固定CO2
微藻具有从空气中固定CO2的能力,并将CO2作为无机碳源供其生长和代谢[45]。据报道,每生产1 t微藻类生物质可以固定1.83 t CO2,其固碳效率是陆生植物的10~15倍[46]。表1总结了不同种类的菌藻共生系统减污降碳性能。

表1 不同种类的菌藻共生系统减污降碳性能
微藻有3种固碳模式,分别为光合自养固碳、异养固碳和兼养固碳[17]。光合自养固碳是指微藻通过光合作用,在叶绿素或其他光合色素的作用下将CO2和H2O转化为三磷酸腺苷(ATP)和还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)等光反应产物的过程[51],微藻细胞光合作用固碳机理如图4所示。微藻细胞中的光合作用分光反应和暗反应两个阶段进行。光反应由光驱动,利用光能将烟酰胺腺嘌呤二核苷酸磷酸(NADP+)和二磷酸腺苷(ADP)转化为储存能量的NADPH和ATP。暗反应不需光参与,且与光反应同时发生,微藻细胞利用光反应过程中产生的NADPH和ATP固定大气中CO2。暗反应中,微藻通过卡尔文循环的羧化、还原和再生三阶段来实现CO2捕获。羧化阶段,在核酮糖-1,5-二磷酸羧化酶(RuBisCo)的催化作用下,CO2与核酮糖-1,5-二磷酸(RuBP)结合生成3-磷酸甘油酯(3-PGA),从而实现CO2固定。还原阶段,在3-磷酸甘油酯激酶的催化下,3-PGA被ATP磷酸化形成1,3-二磷酸甘油酯;然后在甘油醛磷酸脱氢酶的催化下,1,3-二磷酸甘油酯被NADPH还原生成3-磷酸甘油醛(G-3-P)。在最后的再生阶段,G-3-P经过一系列反应生成核酮糖-5-磷酸,在核酮糖磷酸激酶的作用下生产RuBP,重新进入羧化阶段,开启下一个卡尔文循环[51-52]。微藻异养固碳是指在没有外加无机碳源条件下,以有机碳作为电子供体,微藻通过光合自养生长过程将自身呼吸代谢产生的所有CO2全部吸收[53]。微藻兼养固碳时,藻细胞会优先进行光自养过程[53],通过光自养途径产生的O2不断积累,在微藻细胞内形成高浓度DO环境时促进微藻的异养过程的发生。
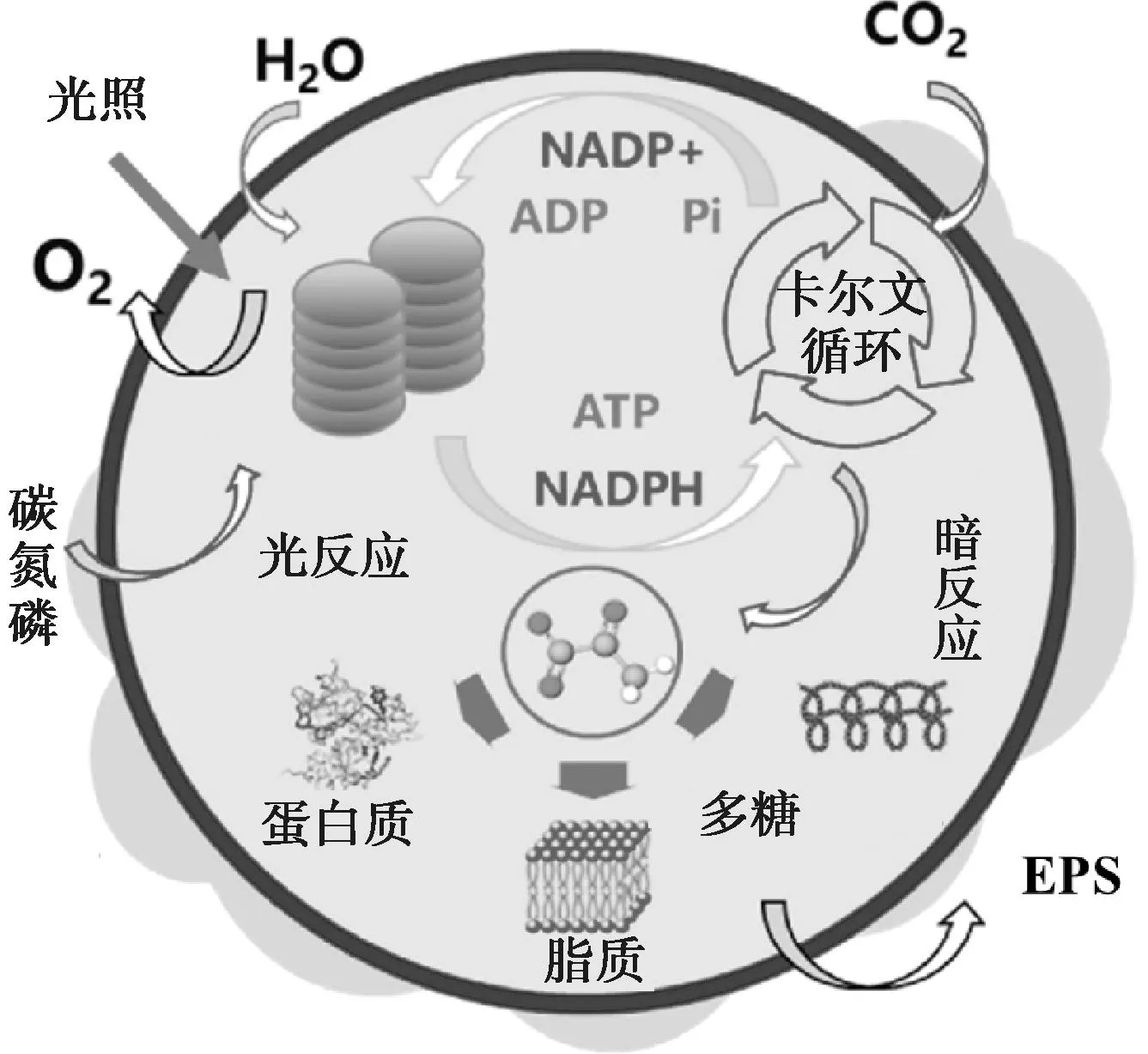
注:Pi为游离的磷酸基团。

4 结论与展望
针对水生态环境污染严重、现有水处理工艺能耗高、温室气体大量排放等问题,菌藻共生系统为去除废水中氮磷污染物以及减少温室气体排放提供了一种绿色可持续的解决方案。文章综述了菌藻共生体系提高减污固碳的作用效能及作用机制,微藻可为硝化细菌提供O2促进对氨氮的转化,菌藻共生系统生成的EPS能够减轻强光照射对菌藻共生系统细胞活性的抑制作用,保证氨氮相关转化菌的活性。夜间藻类呼吸会消耗大量的DO有利于营造缺氧环境,有助于反硝化作用的进行。微藻对磷酸盐的同化是菌藻共生系统中磷酸盐去除的主要机制,细菌通过提供维生素和植物激素来促进微藻生长,从而有助于磷的去除。微藻利用光合作用通过光反应和暗反应两个阶段来实现CO2固定,提高CA和RuBisCO羧化酶的活性可以提高微藻固定CO2的效率。菌藻共生系统在降解水中氮磷污染物及CO2固定上展现出广阔的应用前景。
目前,菌藻共生系统在减污降碳领域的研究仍处于起步阶段,菌藻共生系统的规模化应用及微藻生物燃料的资源化回收面临一些障碍。笔者认为未来菌藻共生系统在减污降碳领域的发展可能主要分为以下几个方向:(1)高效减污降碳菌藻的筛选,各种现有菌藻之间按照不同物质、比例、方式、条件进行复合,调配出高性能减污降碳的菌藻共生系统;(2)开发附着生长的菌藻生物膜系统,使其易于从水中分离沉降、结构稳定,并实现规模化应用;(3)新型菌藻共生催化剂的开发,随着化工和材料科学的发展,越来越多的人工和天然材料作为菌藻共生系统高效减污降碳的催化剂供人们研究开发。总之,构建高效可靠的菌藻共生系统,降低实际运行维护及回收成本,将进一步加速菌藻共生系统在去除污水中氮磷污染物和固定CO2领域的实际应用。
猜你喜欢
科学(2022年4期)2022-10-25
古今农业(2022年1期)2022-05-05
疯狂英语·新读写(2018年3期)2018-11-29
环境保护与循环经济(2017年3期)2017-09-26
电镀与环保(2016年2期)2017-01-20
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
食品工业科技(2014年23期)2014-03-11
无机化学学报(2014年12期)2014-02-28
