
厌氧消化酸累积条件下强化产甲烷研究与应用
2024-03-07 11:59:46王云琦杜玉莹梅红汪炎
工业用水与废水 2024年1期
王云琦, 杜玉莹, 梅红, 汪炎
(东华工程科技股份有限公司, 合肥 230088)
厌氧消化是利用微生物在厌氧条件下代谢去除有机污染物的一种方法[1], 由于其具有运行能耗低、 处理负荷高、 产泥量低且可回收能源等优势,目前已经广泛应用于各类有机废水的处理处置过程中[2]。 厌氧消化通常被认为分为4 步: 水解、 酸化、乙酸化、 产甲烷[3]。 在这个过程中, 酸化、 乙酸化(统称产酸阶段)与产甲烷阶段存在着精准的平衡,以保证厌氧消化中有机物更好地向甲烷转化[4-5]。由于厌氧消化是包含产酸菌、 产乙酸菌和产甲烷菌等多种微生物之间的复杂相互作用的过程, 不同功能的微生物生长和代谢速率不同[6], 对pH 值等环境条件要求也不尽相同[7], 产酸阶段和产甲烷阶段之间精准的平衡很容易被打破, 导致挥发性脂肪酸(VFAs)出现积累, 系统pH 值降低最终使厌氧消化系统酸化崩溃[8-9]。 对于某些特定的工业废水的处理, 在利用厌氧消化处理时, 由于其pH 值低, 且主要成分为高浓度的短链脂肪酸、 醇, 因此也经常会发生厌氧消化系统崩溃现象[10]。
为了避免发生酸累积, 保证厌氧消化正常运行, 在实际生产中, 通常采用投加碱的方式提高系统pH 值, 防止产甲烷过程被抑制[11-12], 这又带来了药剂的消耗以及废水含盐量升高的问题, 给污水处理厂增加了额外的成本。 随着对酸累积抑制产甲烷机理研究的深入, 研究重点逐渐转移到酸累积条件下强化产甲烷能力的方向上, 尝试采用生物强化、 外源强化等方法对酸累积条件下厌氧消化强化产甲烷进行研究, 取得了不错的效果[1]。 本文综述了厌氧消化中酸累积产生的原因、 影响因素及发生酸累积情况下强化产甲烷的方法, 以期为保证厌氧消化反应器的正常运行提供参考。
1 酸累积原因及抑制机理
1.1 酸累积原因及其对产甲烷菌的影响
有机物厌氧消化过程如图1 所示。 厌氧消化过程中首先发生有机物的水解, 在这个过程中大分子如蛋白质、 碳水化合物和脂肪分别被细菌和真菌分解成氨基酸、 多糖和脂肪酸。 在厌氧消化的酸化阶段, 产酸菌将糖、 氨基酸和脂肪酸转化为短链脂肪酸(如丙酸、 丁酸)、 醇和酮、 乙酸、 CO2和H2。 随后在乙酸化阶段产氢产乙酸菌将短链脂肪酸和醇转化为乙酸、 H2和CO2, 最后这些产物被产甲烷菌用来产生甲烷[13]。 由于产甲烷阶段是抑制厌氧消化速率的关键步骤[14], 当进水负荷过高时, 酸化和乙酸化过程产生大量的VFAs, 这些VFAs 无法在短时间内被产甲烷菌利用, 便会造成VFAs 的累积, 导致系统pH 值持续降低。
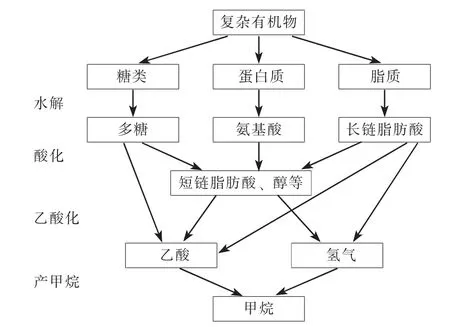
图1 有机物厌氧消化过程Fig.1 Process of anaerobic digestion of organic matters
VFAs 的累积和pH 值的下降会抑制产甲烷菌的活性, 李建政等[15]发现降低消化体系的pH 值,甲烷逐渐从产物中消失。 Horn 等[16]研究结果表明VFAs 浓度是甲烷生成的关键抑制剂, 在pH 值为4.5 时, 乙酸质量浓度仅为0.3 g/L 时, 氢营养型甲烷的生成完全被抑制。 产甲烷菌活性受到抑制导致VFAs 进一步累积和系统pH 值进一步降低, 直到厌氧消化系统崩溃。
1.2 酸累积影响因素
厌氧消化中酸累积的发生受多种因素的影响,如VFAs 的浓度与种类、 系统pH 值等。 Sun 等[17]的研究发现当pH 值为5.1 时, 几乎90% 的乙酸营养型甲烷菌和氢营养型甲烷菌在6 d 的培养过程中从培养物中消失, 低pH 值环境提高了产甲烷微生物的死亡速度, 且这种抑制是不可逆的, 而产酸菌对酸性环境的耐受性较强。 Xiao 等[18-19]利用两相厌氧工艺研究了乙酸浓度对产甲烷菌的抑制作用,在相同pH 值条件下乙酸浓度对产酸相产甲烷菌没有抑制作用, 但对产甲烷相产甲烷菌的抑制作用明显, 从而造成酸累积。 Zhang 等[20]研究了在不同pH 值条件下不同的乙酸浓度对产甲烷作用的影响,结果表明自由乙酸的浓度与甲烷的抑制作用成线性关系, 自由乙酸浓度是影响酸累积的主要原因。
厌氧消化系统中产甲烷代谢途径主要有2 条,乙酸营养型和氢营养型[21-22], 产甲烷菌无法直接利用丙酸等短链脂肪酸, 需要互营产乙酸菌将其降解为乙酸和氢气才能继续产甲烷, 方程式如下所示:
丙酸降解吉布斯自由能大于0, 只有在降解产物浓度低以及氢分压极低的情况下该反应才能向右进行, 发生丙酸的降解。 因此, 越来越多的研究者认为丙酸等短链脂肪酸是影响酸累积的关键因素[23]。
2 酸累积条件下增强产甲烷方法
2.1 生物强化
生物强化法是向厌氧消化系统中投加某一种或某几种功能的微生物来提高厌氧消化系统中产甲烷能力。 传统认为, 产甲烷菌最适pH 值范围为6.8 ~7.2[24], 随着研究的深入, 产甲烷菌被发现可在更低的pH 值条件下生存和代谢。 Wolfe 等[25]在西伯利亚泥炭沼泽中首先发现产甲烷微生物Methanobacillus在pH 值为4.0 条件下能正常生长, Williams 等[26]在明尼苏达泥炭沼泽中得到了嗜酸产甲烷微生物, 该微生物可在pH 值为3 的条件下产甲烷, Sizova 等[27]在西伯利亚泥炭沼泽中富集得到了2 个含嗜酸产甲烷的微生物菌群, 其最适的生长pH 值为4.9±0.5。Savant 等[28]从酿酒废水厌氧处理酸化反应器的污泥中分离了一种嗜酸产甲烷菌Methanobrevibacter acididurans, 并用该菌与常见的产甲烷菌Methanobacterium bryantii 的厌氧消化处理效果进行了对比,结果表明, 在VFAs 浓度高时, 特别是高浓度乙酸存在的条件下, 前者表现出明显的优势, 并且无论是在酸化反应器还是在产甲烷反应器中投加该菌种的富集物, 均可提高甲烷产率并降低VFAs 在系统中的积累。
利用嗜酸产甲烷菌进行生物强化研究较少, 主要原因在于嗜酸产甲烷菌主要被发现于酸性泥炭沼泽等地区, 且分离提纯较复杂。 近年来的生物强化研究主要集中在富集丙酸产甲烷微生物方面。
Li 等[29]在一个半连续式反应器中通过每日调节消化液pH 值对微生物进行驯化, 结果表明, 被驯化的微生物在pH 值为4.8 ~5.5 条件下可降解丙酸, 产生甲烷, 反应器内的优势产甲烷菌由乙酸营养型产甲烷菌(占产甲烷菌总数的87%)转变为氢营养型产甲烷菌(占产甲烷菌总数的67%)。 将该富集的菌群加入到发生酸累积的反应器中, 可以通过降解累积的VFAs 和提高系统pH 值来恢复系统产甲烷能力, 且生物强化后, 乙酸营养型产甲烷菌从占产甲烷菌总数的5% 提高到35%。 Tale 等[30]在严格厌氧和微氧条件下(氧气负荷为有机负荷的1/10)分别富集丙酸降解产甲烷微生物, 并将2 种条件下富集的微生物投加到反应器中进行生物强化, 并与未进行生物强化的反应器进行对比, 结果表明, 在发生酸累积后, 经过生物强化的反应器比不经过生物强化的反应器提前恢复处理效果, 与严格厌氧条件富集的微生物相比, 微氧条件下富集的菌群具有更高的产甲烷活性。
2.2 外源强化
细菌与古细菌之间的种间电子传递可以突破热力学障碍, 是微生物互营产甲烷的关键[31]。 种间电子传递分为间接电子传递和直接电子传递(DIET)2种类型, 其中DIET 不需要电子穿梭体, 细胞与细胞之间可实现直接电子传递[32]。 外源强化是利用该原理, 通过向厌氧消化体系中投加磁铁矿、 活性炭等导电材料, 加强DIET 功能, 加速VFAs 的降解利用和产甲烷速率。
2.2.1 磁铁矿强化
Viggi 等[33]的研究表明, 添加微米大小的磁铁矿(Fe3O4)颗粒可提高丙酸盐的甲烷产量(高达33%), 推测这种刺激效应很可能是因为建立了一种DIET, 基于磁铁矿颗粒作为丙酸氧化型产乙酸菌和二氧化碳还原型甲烷菌之间的电子通道。 王德欣[34]通过投加磁铁矿粉末的方式加强厌氧消化处理费托合成废水, COD 去除率达到(84.3±2.0)%,在该条件下, Geobacter 和Methanosaeta 2 种菌属之间形成了以Fe3O4为基础的DIET 机制, 且其取代了传统的种间氢转移(IHT)机制而成为主导性的电子传递机制。 Yamada 等[35]研究了补充导电氧化铁(磁铁矿)对嗜热厌氧消化池的嗜热产甲烷微生物群落的影响, 在嗜热条件下, 添加磁铁矿可诱导有机酸氧化菌和产甲烷菌之间的电共生, 加速产甲烷。
2.2.2 纳米零价铁(ZVI)强化
ZVI 除了具有良好的导电性可参与DIET 外,还可以降低厌氧消化反应器的氧化还原电位(ORP), 为产甲烷菌等专性厌氧微生物创造了良好的环境, 因此也常常用于加强产甲烷的研究。Zhong 等[36]研究了不同粒径的ZVI 对产甲烷的强化作用, 结果表明在ZVI(48 μm)投加量为2 g/L 的条件下对照组比空白组增加了84.12% 的甲烷产量, ZVI 可使拟杆菌门和变形菌门的丰度分别从1.46% 和0.07% 提高到15.56% 和2.27%, 使产甲烷菌的丰度从3.59% 提高到51.64%, 基因预测结果表明ZVI 颗粒能够促进细胞复制, 提高甲烷生成酶的活性, 此外, 还可以介导发酵菌和产甲烷菌之间的直接种间电子传递。
2.2.3 炭材料强化
除了磁铁矿外, 不同类型的炭材料也常被作为DIET 的介质。 活性炭是由炭材料经过物理或化学活化制备的高微孔导电材料, 具有高比表面积、 高孔容及高吸附性能的优点。 Yang 等[37]在间歇式厌氧污泥消化反应器中加入颗粒活性炭, 当活性炭的添加量从0 增加到5.0 g 时, 产甲烷量提高了17.4%;在20 d 的消化过程中, 活性炭的投加明显富集了氢型产甲烷菌、 Geobacter 和其他能直接进行种间电子转移的产甲烷菌, 增强了合养菌和产甲烷菌之间的电子交换, 加速了底物消耗和甲烷的产生。
生物炭是由天然生物质在缺氧条件下经过热解、 碳化等处理而制成的多孔碳基导电材料。 Qi等[38]发现投加高温(900 ℃)热解产生的生物炭的反应器甲烷含量从对照组的抑制值60% 左右提高到理论值75%, 微生物群落和代谢途径的宏基因组分析结果表明, 假单胞菌可以通过生物炭作为电子接触体, 通过还原二氧化碳参与同甲烷菌的DIET,从而提高甲烷代谢。
石墨烯是一种新型碳材料, 其极高的导电性可为提高厌氧消化的稳定性和效率提供解决方案。Tian 等[39]探索了石墨烯对微生物产甲烷的潜在影响, 结果表明, 石墨烯添加量为30 mg/L 时对甲烷产率有显著的积极影响, 乙酸盐消耗的甲烷生成增强, 长期运行后, 甲烷产率比未投加石墨烯的对照组提高14.3%; 微生物测序数据显示, 添加石墨烯的污泥中, 古细菌群落结构基本保持不变, 其中以Methanoregula、 Methanosaeta 和Methanospirillum 为优势种, 此外, 石墨烯还富集了Geobacter, 这表明石墨烯可能促进了物种间的DIET。
2.3 其他强化方法
微量元素, 如Fe、 Co、 Ni 等可作为产甲烷微生物胞外电子传递的导体, 同时也是产甲烷过程所需酶的重要组成部分[40]。 Zhang 等[41]的研究发现适量添加微量金属元素可显著提高甲烷产量, 在Fe、Co、 Mo、 Ni 的投加量分别为100、 1、 5、 5 mg/L的条件下, 反应器甲烷产率最高, 比未添加金属元素的反应器增加了35.5%。
电化学系统可以提供能量, 促进丙酸、 丁酸的降解, Jiang 等[42]在餐厨垃圾的厌氧消化中施加了外加电场来解决酸累积的问题, 在没有施加电压的情况下, 酸累积严重抑制了甲烷的生成; 相比之下,当施加电压为0.6 ~1.2 V 时, 酸累积得到缓解,VFAs 质量浓度始终低于6 000 mg/L。 Hou 等[43]将厌氧消化反应器(AD)作为藻类辅助微生物燃料电池(AMFC)的阳极室, 构建了AD-AMFC 系统处理厨余垃圾, 研究结果表明, AD-AMFC 系统的甲烷产出量是AD 系统的2 倍多, 生物电化学系统和藻类成功地提高了AD 性能和能量产量。
3 结论与展望
厌氧消化作为一种成熟的废物处理及能源回收技术, 目前已广泛应用于各类有机污染物的处理处置过程中。 厌氧消化中产酸和产甲烷的平衡容易被打破从而导致酸累积, 酸累积进一步抑制产甲烷作用最终导致反应系统崩溃。 随着研究的深入, pH值、 VFAs 浓度和种类被认为是酸累积抑制产甲烷的原因, 通过生物强化和外源强化, 可以增强酸累积条件下厌氧消化的产甲烷能力。
考虑到厌氧消化的复杂性和稳定运行厌氧消化反应器的重要性, 有必要对以下几个方面进行研究: ①加强酸累积对产甲烷作用抑制的机理研究。目前的研究虽然得到了不同条件对产甲烷作用的抑制, 但未从代谢机理角度对其抑制作用进行研究,加强酸累积对产甲烷菌群代谢机理的研究可以为寻找更好的强化方法奠定基础; ②通过数学模型对酸累积的发生进行预警和调控。 深入研究pH 值、VFAs 的种类和浓度、 碱度、 氨氮浓度等厌氧消化指标相互之间的关系及对酸累积的影响, 在此基础上通过数学建模来实现监测某些特定指标, 从而对厌氧消化系统的酸化进行预警, 避免酸累积的发生或在酸累积发生后指导选择合适的调控方法和剂量, 在快速恢复厌氧消化系统效能的前提下减少成本消耗; ③复合强化方法的研究。 目前研究的酸累积条件下强化产甲烷的方法均比较单一, 没有考虑结合不同的方法对强化进行研究。 复合强化方法结合不同的强化产甲烷的方法以提高强化效果, 降低强化产甲烷作用的成本。
猜你喜欢
智富时代(2019年5期)2019-07-05 18:43:32
黑龙江八一农垦大学学报(2017年5期)2017-10-31 06:17:12
湖南农业科学(2017年7期)2017-03-04 02:01:03
现代食品(2016年24期)2016-04-28 08:12:06
化工进展(2015年3期)2015-11-11 09:07:41
医学研究杂志(2015年5期)2015-06-10 06:43:26
河北建筑工程学院学报(2015年2期)2015-04-29 12:23:52
河北建筑工程学院学报(2015年4期)2015-03-24 09:48:11
应用化工(2014年3期)2014-08-16 13:23:50
环境工程技术学报(2014年5期)2014-07-19 06:44:22
