
Soil and microbial C:N:P stoichiometries play vital roles in regulating P transformation in agricultural ecosystems:A review
2024-03-07 07:33GuangleiCHENJiahuiYUANShenqiangWANGYutingLIANGDengjunWANGYiyongZHUandYuWANG
Pedosphere 2024年1期
Guanglei CHEN ,Jiahui YUAN ,Shenqiang WANG ,Yuting LIANG ,Dengjun WANG ,Yiyong ZHU and Yu WANG,*
1State Key Laboratory of Soil and Sustainable Agriculture,Changshu National Agro-Ecosystem Observation and Research Station,Institute of Soil Science,Chinese Academy of Sciences,Nanjing 210008(China)
2School of Life Sciences,Jiangsu Normal University,Xuzhou 221000(China)
3School of Fisheries,Aquaculture,and Aquatic Sciences,Auburn University,Auburn AL 36849(USA)
4Jiangsu Collaborative Innovation Center for Solid Organic Waste Resource Utilization, College of Resources and Environmental Science, Nanjing Agricultural University,Nanjing 210095(China)
ABSTRACT Stoichiometry plays a crucial role in biogeochemical cycles and can modulate soil nutrient availability and functions.In agricultural ecosystems,phosphorus(P)fertilizers(organic or chemical)are often applied to achieve high crop yields.However,P is readily fixed by soil particles,leading to low P use efficiency.Therefore,understanding the role of carbon:nitrogen:P stoichiometries of soil and microorganisms in soil P transformation is of great significance for P management in agriculture.This paper provides a comprehensive review of the recent research on stoichiometry effect on soil P transformation in agricultural ecosystems.Soil microorganisms play an important role in the transformation of soil non-labile inorganic P to microbial biomass P by regulating microbial biomass stoichiometry.They also mobilize soil unavailable organic P into available P by changing ecoenzyme stoichiometry.Organic materials,such as manure and straw,play an important role in promoting the transformation of insoluble P into available P as well.Additionally,periphytic biofilms can reduce P loss from rice field ecosystems.Agricultural stoichiometries are different from those of natural ecosystems and thereby should receive more attention due to the influences of anthropogenic factors.Therefore,it is necessary to conduct further stoichiometry research on the soil biochemical mechanisms underlying P transformation in agricultural ecosystems.In conclusion,understanding stoichiometry impact on soil P transformation is crucial for P management in agricultural ecosystems.
Key Words:ecoenzyme,microbial biomass,microbial community,microorganism,P availability,periphytic biofilm,soil P transformation
INTRODUCTION
Carbon (C),nitrogen (N),and phosphorus (P) are essential elements for plants and microorganisms(Finziet al.,2011;Oldroyd and Leyser,2020).The coupling between C,N,and P is the basis of primary production and accumulationdecomposition in ecosystems (Mackenzieet al.,2002;Delgado-Baquerizoet al.,2013).This coupling can also affect P biogeochemical cycling and is thus key for ecosystem functionality(Zechmeister-Boltensternet al.,2015;Achatet al.,2016;Sardanset al.,2021).The average global C:N:P ratio in soil ranges from 287:17:1 to 186:13:1(Cleveland and Liptzin,2007;Xuet al.,2013)and differs between natural and agricultural ecosystems.For example,it is from 391:21:1 to 86:5:1 in forest ecosystems(Cleveland and Liptzin,2007;Xuet al.,2013;Zhanget al.,2018;Zhenget al.,2021),from 82:10:1 to 28:4:1 in grassland ecosystems (Du and Gao,2021;Yanget al.,2022),and 64:5:1 in farmland(Zhenget al.,2021).The lower C:N:P ratios in agricultural ecosystems might be directly related to the use of phosphate fertilizers (Wanget al.,2010;Rodrigueset al.,2021).In other words,P fertilizer input in agricultural ecosystems may have altered the soil ecological stoichiometry,leading to imbalanced soil C:N:P ratios,which weaken agricultural ecological functions and eventually result in resource waste and low crop productivity.
When applied to soils,P fertilizers are easily fixed by soil minerals,such as iron and aluminium minerals,and become unavailable to crops (Dumaset al.,2011;Arai and Livi,2013).Excessive use of P fertilizers not only leads to low P use efficiency but also results in P loss to the environment.A meta-analysis shows that P addition upsets the balance of terrestrial ecosystem C:N:P stoichiometry (Sunet al.,2022)and in turn affects microbial growth and community diversity,the trophic web structure,and ecosystem functions(Mariotteet al.,2017;Shaoet al.,2017;Peñuelaset al.,2020).For example,when microorganisms are P-limited,the relative abundances of Actinobacteria andSphingomonasincrease,whereas that of Acidobacteria decreases(Cui JWet al.,2022).Additionally,P input reduces the C:P ratio of microbial biomass,possibly because P input has a stronger promoting effect on bacterial abundance than on fungal abundance,and bacteria generally exhibit a lower C:P ratio compared to fungi(Mouginotet al.,2014).Furthermore,rare taxa(i.e.,taxa with low abundances)are more sensitive to soil P limitation and play a more important role in driving organic P(Po)mineralization(Xuet al.,2022).In summary,fertilizer input to agricultural ecosystems leads to imbalance in soil C:N:P ratio(Peñuelas and Sardans,2022;Sunet al.,2022),and the excessive use of P fertilizers and low P use efficiency urgently require attention to the effect of stoichiometry on P transformation.
Soil microorganisms control key biogeochemical processes in agroecosystems and play a vital role in nutrient cycling(Daiet al.,2018,2020).They can partially offset soil stoichiometric imbalances by slowing down the cycling of limiting bio-elements while accelerating that of abundant bio-elements (Chenet al.,2018).On a global scale,the labile inorganic P(Pi)concentration is primarily controlled by biological processes such as microbial uptake,immobilization,and mineralization(Houet al.,2016;Heet al.,2023).Through life strategy change(Biet al.,2020;Weiet al.,2020),microbial community structure change(Luoet al.,2017;Chen G Let al.,2022),and extracellular ecoenzyme secretion(Cuiet al.,2020a,2021)to adapt to the changing resources(Fig.1),the stoichiometry imbalance of organic matter accumulation can be alleviated(Mooshammeret al.,2014).For example,manure application can improve soil stoichiometry,promote P absorption by microorganisms,and stimulate the transformation of soil moderately labile P into microbial biomass P(Chen G Let al.,2022).The microbial mineralization of soil organic matter is accelerated at low soil C:N and C:P ratios,which allows microbes to obtain C for metabolism and utilize the available P(Zechmeister-Boltensternet al.,2015).While progress has been made in regulating C and N transformation pathways in terrestrial ecosystems,the important role of P in stoichiometry has not been fully highlighted due to the lack of P-related data in meta-analyses(Cleveland and Liptzin,2007;Xuet al.,2013).Hence,this review focuses on the stoichiometry response to resource changes in agricultural ecosystems,especially the effect of stoichiometry on P transformation so as to promote rational application of P fertilizers and improve P use efficiency in agricultural ecosystems.
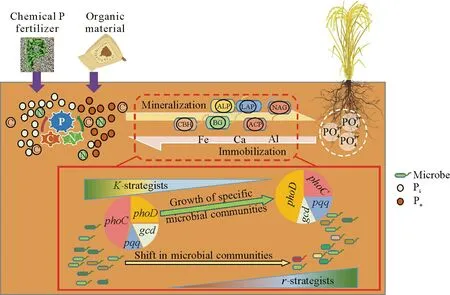
Fig.1 Schematic showing the influence of stoichiometry imbalance caused by resource input on soil P transformation.Pi=inorganic P;Po=organic P.ALP=alkaline phosphatase;ACP=acid phosphatase;BG=β-1,4-glucosidase;CBH=β-D-cellobiosidase;NAG=β-1,4-N-acetylglucosaminidase;LAP=L-leucine aminopeptidase.phoC,phoD,gcd,and pqqrepresent phoC-,phoD-,gcd-,and pqq-harboring bacterial communities,respectively.
MICROBIAL COMMUNITY STRUCTURE CHANGES WITH RESOURCE CHANGE
Microbial diversity is related to soil C,N,and P contents(Delgado-Baquerizoet al.,2016;Zhanget al.,2016;Yanget al.,2020).Soil bacteria can be divided into copiotrophic bacteria (r-strategists) and oligotrophic bacteria(K-strategists)according to their lifestyle(e.g.,growth rate and life strategy) (Trivediet al.,2013;Weiet al.,2020)(Fig.1).K-strategists,such as Acidobacteria,have low growth rates and nutrient use efficiencies,whereasr-strategists,such as Bacteroidetes andα-Proteobacteria,have the characteristics of rapid growth and low C utilization efficiency (Hoet al.,2017).Application of organic materials,such as chicken manure,significantly increases the abundances ofr-strategists,includingα-Proteobacteria,Bacilli,and Bacteroidetes,in soils with high C:N:P ratios (Biet al.,2020).In contrast,K-strategists,such asγ-Proteobacteria,dominate in soils with low C:N:P ratios due to their strong ability to degrade recalcitrant C such as lignin(Poldet al.,2015).When the soil C cycle is dominated by unstable organic matter,the abundance ratio of Gram-positive bacteria(K-strategists)/Gram-negative bacteria(r-strategists)decreases(Cui JWet al.,2022).Previous research showed that long-term (16-year) application of P(superphosphate)had a stronger promoting effect onrstrategists than onK-strategists (Linget al.,2017).This is mainly due to the fact thatr-strategists require more P for growth(Elseret al.,2003;Fiereret al.,2007;Delgado-Baquerizoet al.,2017).Therefore,P fertilizer input alters soil microbial community structure by increasing the abundance ofr-strategists.Additionally,a combination of organic and inorganic fertilizers can significantly increase the abundance of Bacilli(r-strategists),which possessβ-propeller phytase-encoding genes and are capable of hydrolyzing soil phosphates for rapid utilization(Jorqueraet al.,2011).In soils with high C:N:P ratios,Bradyrhizobium and Methylobacterium,alsor-strategists,can mineralize Poby secreting phosphatase(Jorqueraet al.,2011;Ikoyiet al.,2018).In a word,r-strategists may have an advantage overK-strategists in obtaining P for growth.
Microbe-mediated nutrient transformation is a coupled and integrated process(Falkowskiet al.,2008).For example,the synthesis of phosphatase needs C and N as raw materials(Spohn,2016;Wanget al.,2016).Nitrogen inputs can improve soil phosphatase activity(Marklein and Houlton,2012).At the same time,the mineralization of Pomay be driven by microbial C demand (Spohn and Kuzyakov,2013;Heucket al.,2015;Wanget al.,2016;Maet al.,2020).A change in elemental stoichiometry will also affect the expression of microbial functional genes,thus regulating P turnover.For example,it has been reported in the literature that in response to chemical P fertilizer application in paddy soil,the expression ofphoDwas upregulated in Actinobacteria and Cyanobacteria,leading to increased mineralization of Po(Weiet al.,2019).Meanwhile,the abundance of the gene3-phytaseis higher in P-deficient soil than in P-rich soil (Nealet al.,2017),which indicates that phytate can be a good P source in P-limited soils(Oliverioet al.,2020).Moreover,a meta-analysis showed that Proteobacteria,Actinobacteria,and Firmicutes with thephoCorphoDgene are quite sensitive to P fertilizer input or organic amendments,and soil C:P ratio is a main predictor of the diversity and composition of thephoC-andphoD-harboring populations(Luoet al.,2019).Therefore,organic material application appears to be a good strategy for soil P management in agricultural ecosystems.
HIGH MICROBIAL STOICHIOMETRIC RATIO CAN PROMOTE THE TRANSFORMATION OFSOIL NONLABILE Pi TO MICROBIAL BIOMASS P
Soil microbial biomass is a driving force of soil nutrient transformation and circulation (Sinsabaughet al.,2009)(Fig.2a).Soil microbial biomass C:N:P(MBC:MBN:MBP)ratio ranges from 60:7:1 to 42:6:1 at the global scale(Cleveland and Liptzin,2007;Xuet al.,2013;Gaoet al.,2022)and from 175:13:1 to 77:7:1 in forest ecosystems(Faninet al.,2013).However,the MBC:MBN:MBP ratio in cultivated soils ranges from 93:10:1 to 9:1:1 (Tischeret al.,2014;Chenet al.,2017;Biet al.,2020).The different microbial stoichiometries in natural and agricultural ecosystems indicate that they are limited by different nutrients.Indeed,one study found that after returning agricultural land to grassland,the MBC:MBN:MBP ratio changed from 17.8:4.5:1 to 24.3:6.6:1,and the MBC:MBP ratio increased from 17.8:1 in the 1st year to 24.3:1 in the 30th year(Yanget al.,2022).Compared to natural ecosystems,P addition significantly reduces the MBN:MBP and MBC:MBP ratios in agricultural ecosystems,resulting in the alleviation of soil microbial P limitation in farmlands(Sunet al.,2022).
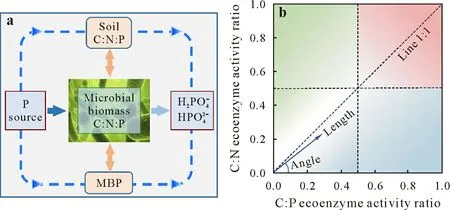
Fig.2 Schematic showing that soil and microbial biomass C:N:P stoichiometries are driving forces of P transformation in soil(a)and the vector model of ecoenzymatic stoichiometry(b)(adapted from Cui et al.,2021).In b,vector length represents soil C limitation for microbes,with vector angle representing soil N or P limitation for microbes,and pink,blue,and green indicate microbial C,N,and P limitations,respectively,with darker color indicating stronger limitation.MBP=microbial biomass P.C:P ecoenzyme activity ratio=ln(BG+CBH):lnALP,where BG,CBH,and ALP are the β-1,4-glucosidase,β-D-cellobiosidase,and alkaline phosphatase activities,respectively;C:N ecoenzyme activity ratio=ln(BG+CBH):ln(NAG+LAP),where NAG and LAP are the β-1,4-N-acetylglucosaminidase and L-leucine aminopeptidase activities,respectively.
In a natural ecosystem,due to the input of plant litter,the resultant increased supply of microbial substrate C and N leads to P restriction on microbial growth(Yanget al.,2020).In an agricultural ecosystem,the application of inorganic fertilizers lowers the soil C:N(P)ratio to below the ratio required for microbial metabolism,leading to C limitation for microorganisms (Cuiet al.,2021).In contrast,organic material(544:8:1 of crop residue C:N:P)inputs with high C:N(P)ratios result in microbial N or P limitation(Liuet al.,2023).Therefore,only material input with balanced C:N:P stoichiometry,126:(6-16):1,is optimal for the growth of microorganisms and crops (Liuet al.,2023).Changes in microbial biomass stoichiometry will affect P transformation in ecosystems.Research showed that the addition of C(glucose)increases the MBC:MBP ratio and promotes the transformation of citrate-P and hydrogen chloride-P(recalcitrant inorganic P forms)to MBP(labile organic P forms)(Sunet al.,2019).The MBC:MBP ratio of poultry litter decreases in the later stage of incubation,which leads to the transformation of Resin-P and sodium bicarbonate-P into Poor MBP (Maliket al.,2013).Pig manure addition leads to the decrease of soil MBC:MBP ratio with time during incubation,which indicates that the P source increases microbial P absorption and promotes the transformation of ferric phosphate and orthophosphate diester to Resin-P and MBP (Chen G Let al.,2022).In summary,increase in the MBC:MBP ratio due to organic material input prompts microorganisms to transform soil labile or non-labile P into MBP,a potential available P source for plants(Richardson and Simpson,2011),thereby reducing the risk of labile P loss and providing a slow-release P source for plants in agricultural ecosystems.
HIGH ECOENZYMATIC STOICHIOMETRY PROMOTES THE TRANSFORMATION OFSOIL Po INTO AVAILABLE P
Ecoenzymes(i.e.,enzymes not bounded by the membranes of living cells) are secreted by microorganisms to obtain soil energy and nutrients by mediating the decomposition of soil organic matter and catalyzing the transformation of soil nutrients (Sinsabaughet al.,2009).Various enzymes are involved in P transformation,including acid and alkaline phosphomonoesterases,phytases,and nucleotidases(Nannipieriet al.,2011).Ecologists have focused on the ecoenzymes involved in the terminal decomposition of organic matters(Sinsabaugh and Follstad Shah,2012).Alkaline phosphatase(ALP),which is mainly produced by microorganisms(Tanet al.,2013),is an important driver of microbial P turnover.Therefore,it is used in the calculation of ecoenzyme activity vector.Here,the C cycle ecoenzymes includeβ-1,4-glucosidase (BG) andβ-D-cellobiosidase (CBH),the N cycle ecoenzymes includeβ-1,4-N-acetylglucosaminidase(NAG)andL-leucine aminopeptidase(LAP),and the P cycle ecoenzymes include ALP(Tapia-Torreset al.,2015;Moorheadet al.,2016;Cuiet al.,2021).Sinsabaughet al.(2009)put forward the concept of“ecoenzymatic stoichiometry”;that is,the ln(BG+CBH):ln(NAG+LAP) ratio represents the ratio of C:N ecoenzyme activities,the ln(BG +CBH):ln(ALP)ratio represents the ratio of C:P ecoenzyme activities,and the ln(NAG+LAP):ln(ALP)ratio represents the ratio of N:P ecoenzyme activities.These ratios are used as nutritional indicators to reflect soil nutrient availabilities and evaluate microbial growth (Sinsabaugh and Follstad Shah,2012;Cuiet al.,2020a,b).The vector length of the ecoenzymatic stoichiometry indicates energy(C)limitation on microbial growth.A vector angle smaller than 45°indicates N limitation,whereas a vector angle larger than 45°indicates P limitation(Fig.2b).
Soil ecoenzymatic stoichiometry reflects the response of microbial cells to the ambient nutrient availability(Tapia-Torreset al.,2015).Compared with agricultural ecosystems,the vector angle of ecoenzyme activity in natural ecosystems is larger(>45°),and microbial metabolism is limited by soil P content(Cuiet al.,2019,2020a).In agricultural ecosystems,due to the addition of C,N,and P,the growth restriction of microorganisms becomes complex,which also provides a new way for exploring P transformation.When microorganisms are P limited,phosphatase activity is increased(the(BG+CBH):ALP ratio is decreased),leading to increased decomposition of Pocompounds(Cuiet al.,2020b).Research has shown that the expression of exopolyphosphatase,which catalyzes Podegradation,is up-regulated with an increase in P limitation,indicating that soil microorganisms mainly decompose active Pocompounds,rather than recalcitrant Pocompounds(Cui JWet al.,2022).Nutrient limitation may lead to changes in microbial community structure and secretion of extracellular enzymes.For example,when ecoenzyme activity is C-or P-limited,C(e.g.,straw)input will enhance the respiratory intensity of microorganisms,increase the quantity of ecoenzyme(e.g.,ALP)secreted by bacteria,and ultimately accelerate the transformation of Poto available P (Rosingeret al.,2019).High organic N (e.g.,cow manure) input can promote the secretion ofL-asparaginase by soil microorganisms,reduce soil N limitation,and finally alleviate P limitation and improve P utilization(Fujitaet al.,2019).Cuiet al.(2020b)reported that after adding N (urea) fertilizer in corn-wheat rotation,the ecoenzyme activity vector indicated P limitation,and the N circulationrelated genes were up-regulated,leading to ALP secretion increase and P conversion.Microbial P limitation (vector angle>45°) was mitigated gradually with the increased addition of P,which may lead to reduced secretion of ALP and improved C metabolism of microorganisms(Rosingeret al.,2019;Cui Y Xet al.,2022),Similarly,microbial P limitation can promote the production of acid phosphatase,ALP,and phytase(Samaddaret al.,2019;Cui J Wet al.,2022)to catalyze the mineralization of inositol phosphates,nucleotides,phosphoproteins,and sugar phosphates(Turneret al.,2005).In agricultural ecosystems,microorganisms determine the conversion of soil Poto available P by adjusting the ecoenzymatic stoichiometry to cope with resource changes.
FUTURE CHALLENGES IN COUPLING C:N:P STOICHIOMETRY WITH NUTRIENT BALANCE IN AGRICULTURAL ECOSYSTEMS
In recent years,research on elemental stoichiometry has advanced rapidly.However,the relation between soil C:N:P stoichiometry and elemental cycling still needs further exploration.
In agricultural ecosystems,attention should be paid to nutrient balance.Currently,there are some vexing issues to be addressed,such as excessive and insufficient fertilizer application.Due to the coupled C,N,and P transformation,the shortage of one element would seriously affect the use efficiency of the other elements.Considering resource conservation and environmental protection,organic fertilizers application or combined application of organic and inorganic fertilizers should be promoted.Organic inputs can reduce resource waste,provide multiple elements,and maintain the balance between elements.However,the mechanism of elemental turnover under organic application is highly complicated,which demands further research.
The coupled utilization of soil elements often involves the action of microorganisms.The complexity of soil microbial community leads to intricate soil C:N:P coupling.This is not only related to the differences in soil amendments(e.g.,pure chemical fertilizers,organic materials,and combined organic-inorganic application) but also related to microbial community competition (e.g.,fungivs.bacteria,Kstrategistsvs.r-strategists).As a result,the mechanism of microbial response to C:N:P coupling is complicated,and there are no consistent results and conclusions.Therefore,researchers should pay more attention to the mechanism of C:N:P balance in agricultural ecosystems.
In recent years,periphytic biofilms (PBs) have been found to play a crucial role in the soil C cycle(Wanget al.,2022;Zhouet al.,2023),N cycle(Chen Z Het al.,2022),and P cycle(Caiet al.,2021;Liuet al.,2021)in rice field ecosystem.Periphytic biofilms are aggregates formed by filamentous algae,bacteria,fungi,and other microorganisms(Wuet al.,2018;Zhouet al.,2023),and PB coverage on paddy soil varies between 20% to over 40% (Liuet al.,2019).Studies have shown that PBs can immobilize more than 1-10 kg P per hectare of rice soil and reduce P loss by approximately 30 000-300 000 t per season in China(Liuet al.,2019).Soil stoichiometry affects P immobilization by PBs.Specifically,high C:P and N:P ratios are beneficial to P immobilization by PBs due to the high abundance of green algae (Liuet al.,2019).Meanwhile,PBs can also produce phosphatase to hydrolyze Pointo orthophosphate(Wuet al.,2018;Caiet al.,2021).The balance between C,N,and P is crucial for P fixation by PBs.Application of soil amendments with high contents of organic C and total N,such as rice straw,is beneficial to P immobilization by PBs(Liuet al.,2019).Therefore,PBs should be taking into account when developing strategies to reduce P loss from rice field ecosystems.
Based on a large body of basic research on C and N,models of C and N cycles for various regions around the world have been gradually established(Baiet al.,2023;Caiet al.,2023;Xiaet al.,2023).However,no meta-analysis of P cycle has been performed,and our understanding of global P cycle is still limited.In the future,basic research on P cycle should be strengthened to improve our understanding of regional P cycle and elucidate the underlying mechanism of C:N:P stoichiometric coupling.
In research on the biogeochemical cycle of C,N,and P,total contents of these elements are typically presented.However,elements present in various forms in soils,total contents cannot sufficiently reflect their availabilities.For instance,strongly adsorbed and occluded inorganic P are considered as inorganic P loss in most C,N,and P models because they are considered as non-bioavailable(Achatet al.,2016).Therefore,further analysis of available C,N,and P is necessary for a more comprehensive understanding of elemental cycling.
CONCLUSIONS
This review highlights the interdependent relationship between stoichiometry and soil P transformation in agricultural ecosystems.Specifically,microbial biomass stoichiometry regulates P fixation and release by microbes,whereas ecoenzymatic stoichiometry mobilizes Po.Changes in the stoichiometries of soil organic matter,soil microbial biomass,and ecoenzyme activities are crucial for soil P transformation.In order to better understand soil P cycling,it is important to take into account soil amendment (e.g.,chemical fertilizers and organic materials) and microbial response effects on different stoichiometries and to pay attention to P immobilization and transformation by PBs in paddy field ecosystems as a means of reducing P loss.This review provides valuable guidance for applying stoichiometry to understand soil P transformation in agricultural ecosystems.
ACKNOWLEDGEMENTS
We acknowledge the financial support from the Natural Science Foundation of Jiangsu Province,China (No.BK20230049),the National Natural Science Foundation of China(No.42277026),the 14th Five-Year Plan Innovation Program of the Institute of Soil Science,Chinese Academy of Sciences (No.ISSASIP2201),the Major Science and Technology of Inner Mongolia Autonomous Region,China(No.NMKJXM202009),and the Program for Excellent Postdoctoral Talents of Jiangsu,China(No.2022ZB533).

- Pedosphere的其它文章
- Developing the new soil science-Advice for early-career soil scientists
- Biophotoelectrochemistry:An emerging frontier for channeling photoelectric effect into darkness zone of soils and sediments
- Balancing machine learning and artificial intelligence in soil science with human perspective and experience
- Soils in extraterrestrial space:Need for studies under microgravity
- Role of biochar in raising blue carbon stock capacity of salt marshes
- Long-term fertilizer nitrogen management-Soil health conundrum