
脂肪酸平衡在反刍动物上的研究进展
2024-03-07 06:17:12张永根白海鑫
饲料工业 2024年3期
■ 张永根 白海鑫
(东北农业大学动物科学技术学院,黑龙江哈尔滨 150030)
与单胃动物不同,日粮脂质在反刍动物体内首先要经过瘤胃的分解氢化,大部分的不饱和脂肪酸会被氢化为饱和脂肪酸,大大降低了脂质的利用价值。随着现代油脂工艺的发展,氢化脂肪、脂肪酸钙和包被脂肪等过瘤胃脂肪已被广泛应用。通过瘤胃后,脂肪酸更能以其原有的补充形式发挥其营养和生物学功能,因此脂肪酸平衡在过瘤胃后更加强了对反刍动物营养消化代谢、吸收转化和肉奶品质的影响。脂肪酸营养是六大营养素研究的一块短板,脂肪、脂肪酸平衡也和蛋白质、氨基酸平衡同等重要。
1 脂肪酸平衡的概念及重要性
脂肪酸平衡又被称为理想脂肪酸模式,是指根据动物在不同时期的代谢吸收特征、机体需求特征给予其最精准的脂肪酸搭配,最大限度地促进脂肪酸向肉蛋奶的转化和富集[1]。
1.1 从外源摄取角度看脂肪酸平衡的重要性
动物从日粮中摄取的脂质主要包括三酰甘油、磷脂和半乳糖脂等几种类型,其中摄入最丰富的是三酰甘油。一分子的三酰甘油由三分子的脂肪酸和一分子的丙三醇通过酯键酯化生成,所以饲料脂质中脂肪酸的不同决定了脂质发挥不同的生物学效应。常规饲料原料和油脂中的脂肪酸构成比较稳定且具有一定规律。由图1 可见,玉米、小麦和亚麻籽等常规饲料原料中的脂肪酸主要以C16∶0、C18∶0、C18∶1、C18∶2 和C18∶3 为主[2]。而反观动物产品肉和奶中脂肪酸的构成(见图2),脂肪组织和肉中丰富度排名C18∶1>C16∶0>C18∶0,而在牛奶中C16∶0>C18∶1>C18∶0[3]。可见饲料原料和动物产品中脂肪酸的构成和丰富度基本是保持一致的,从同源转化效率更高的角度来看,日粮中脂肪酸的搭配越接近动物产品脂肪酸的构成,即补充的脂肪酸与动物机体越保持平衡,转化效率应该更高。

图2 动物产品中脂肪酸构成
1.2 从内源合成角度看脂肪酸平衡的重要性
动物体内的脂肪以三酰甘油的形式存在,组成三酰甘油的脂肪酸来源于两条途径:从日粮脂肪中直接摄取和利用葡萄糖、乳酸和乙酸为底物从头合成[4-5]。从图3 可知,一分子的底物乙酸在乙酰辅酶A 合成酶(ACS)的作用下生成乙酰CoA,然后在乙酰辅酶A 羧化酶的作用下生成Malonyl CoA,接着在脂肪酸合成酶(FASN)的作用下生成短链脂肪酸,此后在2分子还原型辅酶2(NADPH)的作用下每次以一个2碳单位来延长脂肪酸链[6]。C16∶0 是内源合成的起始脂肪酸,经过延伸可变为C18∶0,然后在硬脂酰辅酶A去饱和酶的作用下在9 号碳位插入一个双键,生成C18∶1。然而内源合成步骤到C18∶1 就结束了。动物体内缺乏在12 号碳位上插入双键的能力,因此C18∶1 不能去饱和生成C18∶2,同样的,C18∶2也不能通过直接去饱和生成C18∶3[7]。C18∶2 和C18∶3 都需要从日粮中摄取,接着再代谢为ω6和ω3系列的多不饱和脂肪酸,所以这两种脂肪酸也被称为必需脂肪酸。常规饲料原料中含有丰富的C18∶2,一般都过剩,所以从内源合成的角度来说,更加强调补充脂肪酸的平衡,既不能缺乏,也不能过多补充造成浪费。
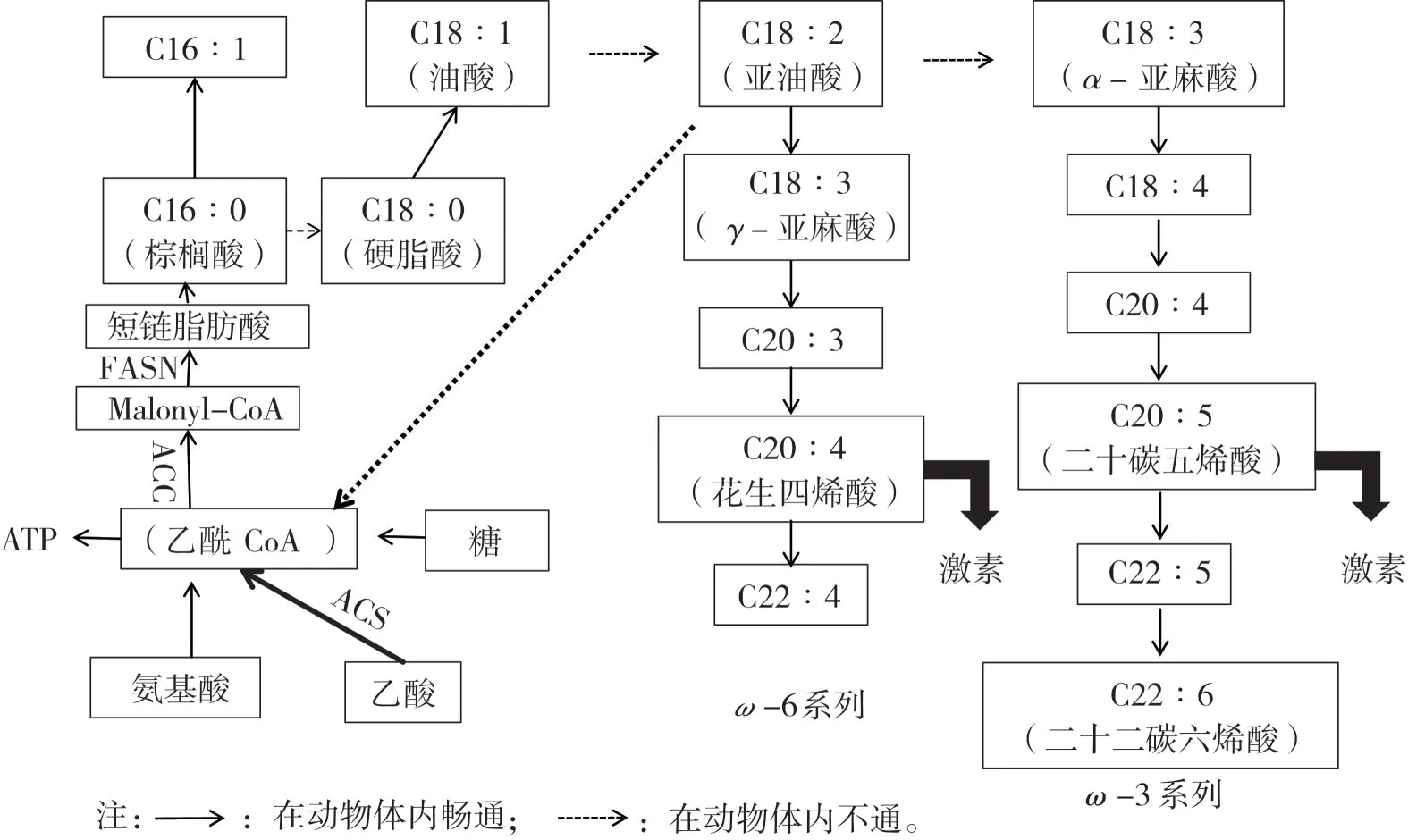
图3 动物脂肪酸合成途径
2 脂肪酸平衡在奶牛上的研究进展
2.1 对奶牛采食、消化和吸收的影响
2.1.1 对采食量的影响
新版的NASEM(2021)[8]建议奶牛日粮中额外添加的脂肪不超过日粮干物质的4%,总日粮脂肪不超过7%。2%~4%的脂肪添加量不会影响奶牛的干物质采食量(DMI),但超过5%则会使DMI 下降[9]。在补充形式上,脂肪酸钙盐形式的过瘤胃脂肪与氢化脂肪、包被脂肪相比,可能由于其钙皂的特殊气味更容易造成奶牛的DMI下降[10]。大量研究表明,补充脂肪的脂肪酸构成也会影响奶牛的DMI,具体来说,在正常补充剂量内(4%以下)饱和脂肪酸不会影响DMI,但随着不饱和脂肪酸比例的增加,DMI逐渐下降。这可能是由于不饱和脂肪酸比饱和脂肪酸更容易引起饱腹感激素胆囊收缩素(CCK)的释放[11-12],由此降低DMI。
2.1.2 对营养物质消化率的影响
Boerman 等[13]总结了61 项研究发现,随总脂肪酸摄入增加和十二指肠流量增加,总脂肪酸的消化率都呈线性下降。从个体脂肪酸来看,C18∶1 在奶牛小肠中的消化率为79%~85%,C16∶0 为75%~79%,C18∶0 为72%~77%,整体趋势为不饱和脂肪酸消化率高于饱和脂肪酸;饱和脂肪酸中C18∶0 消化率低于C16∶0[14]。通过总结了61 项奶牛研究,Glasser等[15]发现,C16∶0消化率随十二指肠流量增加而线性下降,而C18∶0 在高肠流量下,其肠道消化率呈二次下降或线性下降。同样作为饱和脂肪酸,C16∶0 对消化率的不利影响要低于C18∶0。这可能是由于C18∶0在肠道胶束中的溶解度较低,使其从胶束中脱落,导致肠道消化率下降更明显[16]。Prom 等[17]通过真胃灌注C18∶1,发现随C18∶1灌注量(20、40、60 g/d)的增加,总脂肪消化率和吸收量呈线性上升,C18∶1对反刍动物总脂肪消化率的积极影响可能是由于其胶束溶解度更高,肠细胞摄取和再酯化更快,另外,C18∶1 与脂肪酸转运蛋白(FABP)的亲和力更高,这两者的叠加效应可能使C18∶1 具有提高脂肪酸消化率的潜力。根据Western 等[18]的研究,当C16∶0、C18∶0、C18∶1 一起组合饲喂的时候,增加C18∶1补充量不仅能提高18 碳脂肪酸消化率,还能协同提高其他饱和脂肪酸消化率,而增加C18∶0 补充量,则拉低了总脂肪的消化率。这充分说明C18∶1 对于整体脂肪消化率的积极促进作用,因此在补充脂肪时要综合考虑加入一定量C18∶1来提高消化率。
Weld 等[19]对38 项泌乳奶牛补充脂肪的荟萃分析(见图4)表明,中链饱和脂肪酸C12∶0、C14∶0 和不饱和脂肪酸会抑制纤维的消化,而长链脂肪酸钙和饱和脂肪不影响纤维消化,甚至对纤维消化产生有利影响。不饱和脂肪酸对纤维的消化不利,主要有两方面的原因:一是瘤胃内纤维的消化需要微生物与饲料颗粒紧密接触,而覆盖饲料颗粒的脂肪层阻断了饲料颗粒与微生物和水解酶的接触;二是长链不饱和脂肪酸直接附着在微生物细胞膜的脂质双层上,改变细胞膜流动性并阻断转运蛋白,从而直接影响瘤胃微生物的正常功能[20]。

图4 不同脂肪补充类型对NDF消化率的影响
2.2 对奶牛产奶量、乳脂率的影响
Drackley 等[21]总结了伊利诺伊大学10 项研究发现脂肪的添加量和产奶量呈二次下降关系(图5)。最大产奶量为3%~4% DM 脂肪添加量,这与Nasen(2021)的建议添加量(外源补充不超过4%)基本保持一致。从单一脂肪酸的影响来看,C16∶0与C18∶0、C18∶1 相比,可以更好地提高产奶量和乳脂率。但大部分研究表明,C16∶0 对乳脂率和乳脂含量的积极影响大于对产奶量的影响[22]。C16∶0 对奶牛乳脂含量和产奶量的积极影响可以归结为以下两方面的原因:首先,C16∶0 可以刺激乳腺组织丁酸的合成,并将所有从头合成的脂肪酸并入三酰甘油。从而增加乳脂的内源合成,另外16∶0 可以增加能量负平衡时期血液中的神经酰胺,而神经酰胺和产奶量呈正相关关系[23]。
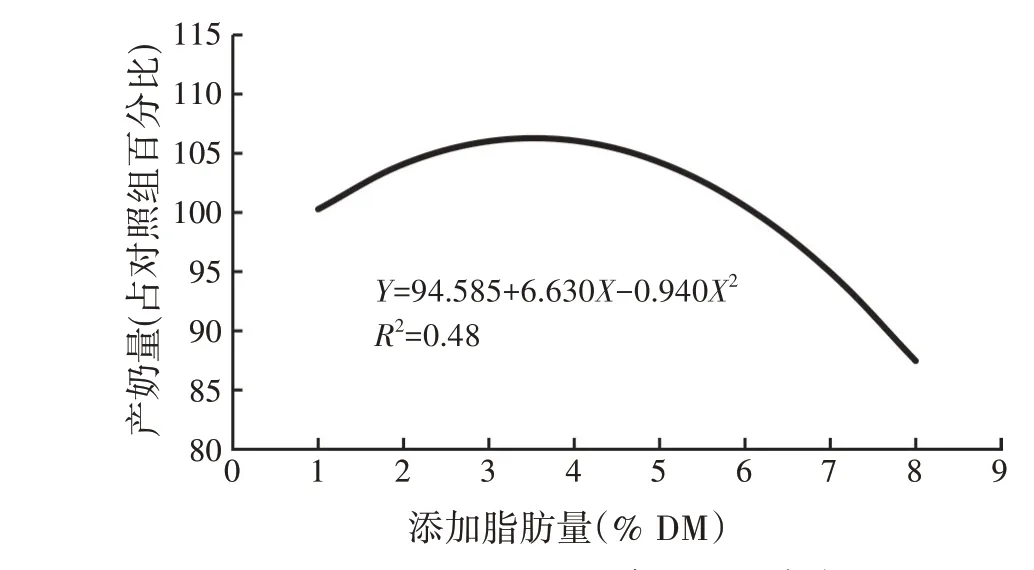
图5 脂肪添加量与产奶量的关系
对于乳脂中脂肪酸的来源,研究表明,有40%左右的脂肪酸可直接从日粮脂肪中摄取、沉积,50%的脂肪酸可利用乙酸为底物从头合成,10%左右可由体脂转移得到。从单一脂肪酸来看,碳链长度小于12 个碳的所有脂肪酸都是通过从头合成得到。大部分的C14∶0 从头合成,只有少部分直接摄取。50%左右的C16∶0 从头合成,另外50%直接从日粮中摄取得到,几乎所有的18碳脂肪酸(包括C18∶0、C18∶1、C18∶2、C18∶3)都是直接从日粮中摄取得到[24]。由此看来,日粮中的脂肪酸构成非常容易直接改变乳脂的脂肪酸构成,特别是16 碳、18 碳的脂肪酸,能直接沉积同化到畜产品中。这更加提示我们在补充脂肪时要考虑脂肪酸结构,提高沉积效率。
2.3 对奶牛能量分配的影响
胰岛素、非酯化脂肪酸(NEFA)通常与奶牛的能量代谢和脂质动员联系在一起,在奶牛围产期,低胰岛素血症会刺激脂质动员,释放NEFA。研究发现,补充高比例的C18∶0使奶牛胰岛素浓度升高,NEFA 降低,减少了奶牛内源脂质的动员,更有利于奶牛恢复体况,C18∶0 在产前和产后早期能更好地被肝脏氧化作为能量来源[25]。另外,以大鼠为模型的研究发现,C18∶1 可快速刺激胰腺β-细胞分泌胰岛素来抑制脂质动员,提示C18∶1 在奶牛上也很可能通过快速的氧化供能来抑制内源脂质动员,从而维持机体能量平衡[26]。研究表明,C16∶0摄入量与用于产奶的能量分配呈正相关关系,而C18∶1 摄入量与用于增重的能量分配呈正相关关系。De Souza 等[27]也发现C16∶0和C18∶1能改变乳腺和脂肪组织之间的能量分配,补充C16∶0 更有利于增加产奶量,而补充C18∶1 更有利于增加体重。这提示要根据奶牛在不同生产阶段的特征给予合理的脂肪搭配,如在泌乳早期能量负平衡时如果大量补充C16∶0,不仅不会使奶牛体况恢复,反而还会加速体脂动员向产奶流失,体况评分下降越发严重。这时要适当考虑补充一定量的C18∶1,在不降低乳脂率的同时再增加一部分能量向体重增加进行分配,逐渐恢复奶牛体况和健康状况。
3 脂肪酸平衡对肉牛生产的影响
C18∶1、C16∶0 和C18∶0 因在肉中丰富的含量(70%~80%)而被称为“造肉脂肪酸”。肉牛育肥期的主要特征是脂肪增加(包括皮下、内脏、肌内和肌间脂肪)。以一头450~500 kg的育肥牛为例,日增重中,超过50%增加的是脂肪(见表1)[28],所以长肉就是长脂肪,也就是增加C18∶1、C16∶0 和C18∶0 的含量。因此日粮中脂肪酸的结构和组成直接决定肉牛长脂肪的量和增重效率。

表1 中等体型阉牛不同生长阶段日增重中脂肪所占比例(%)
3.1 对肉牛育肥性能的影响
脂肪酸通过β-氧化释放的能量是碳水化合物和蛋白质的2.25 倍[29],补充脂肪的能量效率更高。肉牛的体脂处在不断沉积和动员的动态循环中,摄入的脂肪经瘤胃微生物产生的脂肪酶分解后形成游离脂肪酸,游离脂肪酸被小肠吸收,经过淋巴系统通过血液循环运输到外周组织进行β-氧化供能,不必再动员体脂来供能,因此补充脂肪能使能量向正平衡转化,提高肉牛的体脂沉积。肉牛脂肪沉积来源于从头合成和直接摄取;但据报道,脂肪酸从头合成是耗能且低效的,日粮脂质直接以脂滴的形式直接沉积到脂肪组织中的效率更高[30]。Bai 等[31]研究发现,与不补充脂肪的对照组相比,以2.5%日粮干物质补充脂肪酸钙提高了安格斯育肥牛的平均日增重,平均日增重提高的原因可能是干物质采食量(DMI)的提高,根据Fiorentini等[32]的研究,动物生长性能的提高60%~90%可以归因于DMI 的提高,因此,DMI 被认为是决定动物生长性能最重要的因素。
另外,个体脂肪酸可能由于氧化代谢路径先后顺序差异,对肉牛生长性能也表现出不同的效应。特定的脂肪酸可能通过刺激能量代谢基因的表达,从而导致肉牛平均日增重的差异。研究发现,C16:0 与C18∶1相比能更效地促进肉牛体重的增加[31],这与奶牛上体现的是不同的,泌乳奶牛补充C18∶1 更能促进能量向身体储备增加[27]。这说明,C16∶0和C18∶1面对泌乳能量需要和增重能量需要很可能遵循不同的代谢机制通路。当然,也有研究报道,在能量正平衡时期,补充C16∶0 可以增加机体能量储备[33],C16∶0 促进了更多脂肪组织的生成,因其是三酰甘油合成的首选底物[34],也是机体从头合成的起始脂肪酸,可以进一步代谢成C18∶0和C18∶1。
3.2 对肉牛肌内脂肪沉积的影响
嫩度是影响牛肉品质最重要的因素,牛肉的嫩度与肌内脂肪含量、肌纤维特征等息息相关,肌内脂肪同时会影响牛肉的多汁性和口感,是评价牛肉品质的重要因素。影响牛肌内脂肪沉积的主要因素包括品种、基因、性别和营养等[35]。Bai 等[31]研究表明,日粮中补充87% C16∶0+10% C18∶0 与60% C16∶0+30% C18∶1 相比,可以更有效地促进安格斯育肥牛背膘厚的增加和肌肉中脂肪的沉积,增加的肌内脂肪提高了牛肉的蒸煮损失。饱和脂肪酸C16∶0 和C18∶0 促进肌内脂肪沉积的原因可能是这两种脂肪酸与不饱和脂肪酸相比,可以强烈地促进脂肪合成基因的表达,从而产生更多的脂肪[36]。此外Bai 等[31]还发现,虽然补充高浓度的饱和脂肪酸C16∶0和C18∶0 致使背最长肌中饱和脂肪酸的含量有一定提高,但同时不饱和脂肪酸C18∶1 的含量也显著提高,从而平衡了饱和与不饱和脂肪酸的比例,降低了饱和脂肪酸引发健康问题的可能性。
ω6和ω3脂肪酸属于多不饱和脂肪酸的范畴,ω6系列脂肪酸主要包括C18∶2 和C20∶4,ω3脂肪酸主要包括α-C18∶3、C20∶5 和C22∶6。大量研究表明,其具有抗炎、免疫调节、改善繁殖性能的生物活性效应[37]。有研究表明,生长阶段的反刍动物每天需要必需脂肪酸(主要是ω3系列的α-C18∶3 和ω6系列的C18∶2 和C20∶4)88 mg/kg 代谢体重[38]。近年来,一些富含ω6和ω3脂肪酸的藻粉及高山被孢霉等饲料原料被添加至奶牛和肉牛的日粮中,以期实现C20∶4和C22∶6 在奶和肉中的富集,最终生产功能性的产品[39]。研究表明,草饲肉牛比谷饲或集约化生产的肉牛含有更多的ω3多不饱和脂肪酸[40]。向反刍动物日粮中补充油菜籽、棉籽、葵花油和玉米油可以提高牛肉、羊肉中ω6多不饱和脂肪酸的含量,而补充亚麻籽能显著提高肉中ω3脂肪酸的浓度[41]。C18∶2 存在于大多数植物和谷物的种子中,含量非常丰富,因此饲料中ω6脂肪酸的浓度将大于ω3脂肪酸的浓度,并直接导致反刍动物肉中ω6/ω3值更高[42]。因此,在补充ω6和ω3脂肪酸时,应首先了解基础日粮的结构,玉米-豆粕型日粮中一般C18∶2较高,导致ω6脂肪酸的比例偏高,再补充过多的外源ω6脂肪酸很可能导致脂肪酸的失衡浪费。
4 总结与展望
脂肪酸平衡既需要考虑饲料原料中脂肪酸的构成,还需考虑脂肪酸结构对反刍动物消化代谢、吸收转化、生产性能的综合影响。从目前的结果看来,不饱和脂肪酸会通过刺激饱腹感激素(CCK)的释放来降低DMI,增加饱和脂肪酸C16∶0 和C18∶0 的补充会对脂肪消化率产生不利影响,单不饱和脂肪酸C18∶1 能通过提高胶束溶解性来提高自身和协同提高总脂肪消化率。C16∶0 对乳脂率和乳脂产量的正向影响大于对产奶量的正向影响,C16∶0 还能增加能量分配给产奶,而C18∶1 增加能量分配给体重。在肉牛上,C16∶0的增重效果比C18∶1好,但是否由于机体内代谢路径的差异而造成这种不同,还有待深入研究。目前的研究只是初步表明,补充更多的C18∶1 能提高脂肪消化率,更多的C16∶0 能促进生长,既能促进消化也能促进生长的最佳饱和与不饱和脂肪酸平衡比例还未确定,需持续进行深入研究。
猜你喜欢
现代畜牧科技(2021年11期)2021-12-21 06:11:16
中国乳业(2019年12期)2020-01-17 08:47:40
湖南饲料(2019年5期)2019-10-15 08:59:10
分析化学(2018年4期)2018-11-02 10:16:04
天然产物研究与开发(2018年9期)2018-10-08 03:26:00
特产研究(2016年3期)2016-04-12 07:16:12
中国粮油学报(2016年5期)2016-01-23 02:44:57
合成化学(2015年10期)2016-01-17 08:56:06
分析测试学报(2015年8期)2016-01-13 06:19:29
动物营养学报(2015年10期)2015-12-01 03:52:55
