
β-细辛醚通过抑制TRPV4的表达缓解谷氨酸诱导的Ca2+超载
2024-03-04 11:34蒋兰兰陈向涛蔡静雯刘浩宇尹兰香
合肥工业大学学报(自然科学版) 2024年2期
蒋兰兰, 陈向涛, 储 涛, 蔡静雯,2, 刘浩宇, 尹兰香
(1.安徽医科大学 药学院,安徽 合肥 230022; 2.中国科学技术大学附属第一医院(安徽省立医院) 药剂科,安徽 合肥 230001)
随着人口老龄化的加剧,全球神经退行性疾病的负担正在以惊人的速度增加。虽然导致神经退行性疾病的分子机制还不完全清楚,但兴奋性毒性已被证明在其中起着关键的作用[1]。
谷氨酸是中枢神经系统中最重要的兴奋性神经递质,在记忆、突触可塑性和神经元发育中发挥着重要的作用[2],但是谷氨酸被认为是“双刃剑”,既是神经递质又可以作为神经毒素。谷氨酸过度释放会导致Ca2+稳态失调,触发Ca2+下游信号通路的改变,最终导致神经元死亡的过程称为兴奋性毒性,其中Ca2+超载是兴奋性毒性的主要发病机制[3]。由于谷氨酸损伤神经系统具有多靶点效应,谷氨酸导致的神经毒性机制是一个重要的研究课题。
瞬时受体电位香草素受体4(transient receptor potential vanilloid 4,TRPV4)是一种广泛表达、多模态门控[4]、非选择性Ca2+通道参与多种病理和生理反应[5]。当它被内源性和外源性刺激激活时,Ca2+内流增加,导致细胞内游离Ca2+超载[6]。在许多类型的细胞中,TRPV4的活性增加会导致活性氧(reactive oxygen species,ROS)和NO的产生[7],在许多病理过程中会导致细胞凋亡[8],是脑疾病[9]发生的危险因素,包括脑出血[10]、脑水肿[11]、癫痫[12]和阿尔茨海默病[13]。TRPV4的激活上调参与脑缺血时的神经元损伤[14],导致神经元Ca2+超载[15]。TRPV4在细胞毒性方面的作用越来越受到研究者的关注,然而有关TRPV4是否参与谷氨酸诱导的Ca2+超载报道并不多。
高分化的PC12 细胞为大鼠肾上腺髓质嗜铬细胞瘤克隆化的细胞株,具有丰富的神经元性质和特征,近年来,广泛用于细胞神经毒性和神经保护机制的研究。β-细辛醚是石菖蒲的主要挥发油成分,具有抗炎、抗肿瘤等作用,对谷氨酸所致 PC12细胞损伤具有神经保护作用[16]。本实验深入探究β-细辛醚对40 mmol/L谷氨酸诱导的神经毒性的抑制作用及机制。
1 材料与方法
1.1 材料与试剂
谷氨酸(Sigma-Aldrich);β-细辛醚(成都曼思特生物科技有限公司);胎牛血清(四季青),DMEM不完全高糖培养基、EDTA细胞消化液(江苏凯基生物公司);Fluo-4 AM(上海同仁化学研究所);Cell Counting Kit-8 (碧云天);Axygen小量制备总RNA试剂盒(Axygen Silicon Valley);SYBR@Green Pro Taq HS预混型qPCR试剂盒(AG);反转录试剂盒、化学发光显影液(北京全式金生物技术有限公司);脱脂牛奶(伊利);PVDF膜(Millipore);Triton X-100、抗荧光猝灭剂(索莱宝);DAPI(Invitrogen);Anti-TRPV4 (Alomone公司);GAPDH、HRP-羊抗兔IgG、羊抗兔FITC(博士德);NC-siRNA、TRPV4-siRNA、pEX-3-Ctrl、pEX-3-TRPV4试剂(GenePharma);Lipofectamine 2000 (Invitrogen);其余试剂均为进口或国产分析纯。
1.2 仪器与设备
化学发光显影仪、CFX96实时荧光定量聚合酶链式反应(polymerase chain reaction,PCR)仪(Bio-Rad Company,USA);恒温CO2培养箱(The Thermo Company);倒置荧光显微镜(奥林巴斯);恒温摇床(上海天能科技有限公司);离心机(Bio-Rad Company)。
1.3 方法
1.3.1 细胞培养
PC12细胞在37 ℃、5% CO2的湿度环境中培养,添加不完全高糖DMEM培养基(10%胎牛血清、1%青霉素、1%链霉素)。细胞培养48 h后,PC12细胞分别用不同浓度的β-细辛醚(终浓度为15、30、60 μmol/L)预处理4 h后去除溶液,40 mmol/L谷氨酸处理PC12细胞,实时记录Ca2+浓度的变化。在实验中,谷氨酸溶解于HBSS溶液中 (136.9 mmol/L NaCl、5.4 mmol/L KCl、1.3 mmol/L CaCl2、0.4 mmol/L MgSO4·7H2O、0.5 mmol/L MgCl2·6H2O、0.3 mmol/L Na2HPO4·2H2O、0.4 mmol/L KH2PO4、5.6 mmol/L葡萄糖、4.2 mmol/L NaHCO3,调节pH值为7.2~7.4),对照组给予等量的HBSS溶液。β-细辛醚溶于二甲基亚砜(DMSO)中,形成100 mmol/L储备液,保存于-20 ℃。DMSO的最终体积分数小于0.1%。实验分为对照组(A组)、 谷氨酸模型组(B组)、谷氨酸+15 μmol/Lβ-细辛醚组(C组)、谷氨酸+30 μmol/Lβ-细辛醚组(D组)和谷氨酸+60 μmol/Lβ-细辛醚组(E组)。
1.3.2 CCK-8实验检测细胞活力
PC12细胞置于DMEM高糖培养基中,细胞以104个/孔的密度接种在96孔板,培养12 h后,用β-细辛醚(15、30、60、80、100 μmol/L)孵育24 h。每孔加入10 μL CCK-8试剂和90 μL 无血清培养基,在37 ℃下孵育90 min,用酶标仪在450 nm处测定A值。
1.3.3 钙成像技术检测胞内Ca2+浓度
细胞培养48 h,β-细辛醚孵育4 h,用Ca2+荧光探针Fluo-4 AM测定细胞内Ca2+浓度的变化。加入HBSS溶液稀释的2.5 μmol/L Fluo-4 AM在培养箱中孵育40 min,用HBSS溶液洗涤细胞3次。加入HBSS溶液覆盖细胞,37 ℃培养箱孵育25 min,弃除HBSS溶液。使用倒置荧光显微镜,在488 nm波长下激发PC12细胞,10倍物镜将激发光聚焦于细胞。每组至少圈出85个细胞,40 mmol/L谷氨酸处理PC12细胞实时记录Ca2+浓度的变化。利用MetaFluor软件对图像进行采集和分析。计算Ca2+浓度时,每个荧光中减去非特异性Ca2+荧光。
1.3.4 实时荧光定量PCR检测TRPV4的表达
PC12细胞培养在6孔板48 h,β-细辛醚孵育4 h,谷氨酸处理5 min;将培养板置于冰上,预冷的1×PBS洗3次后弃去;加入300 μL Buffer R-1,反复吹打后转移至1.5 mL EP管并加入150 μL Buffer R-Ⅱ,涡旋机混匀30 s,12 000g、4 ℃离心5 min;取上清至2 mL离心柱中,加入200 μL异丙醇,6 000g、4 ℃离心1 min;弃滤液,加入500 μL Buffer W1A,12 000g、4 ℃离心1 min;弃滤液,加入700 μL Buffer W2,12 000g、4 ℃离心1 min;弃滤液,重复上述加入Buffer W2步骤1次;弃滤液,12 000g、4 ℃离心空转1 min;将离心柱放入新的1.5 mL EP管中并加入30 μL TE-Buffer,室温静置1 min,12 000g、4 ℃离心1 min,所得滤液为各组细胞RNA。将制备好的RNA用NanoDrop2000分光光度计进行定量。使用反转录试剂盒按照说明书加样反转录为cDNA。以此为模板,在目标基因的上下游引物作用下,加入热稳定的DNA聚合酶针对目标基因进行扩增。扩增完成后导出实验结果并进行分析。
GAPDH作为内参基因,引物上游序列为:
5′-AGCCCTCCCTTCTCTCGAAT-3′;
下游序列为:
5′-CCCCACAACACTGCATTCAC-3′。
TRPV4作为目的基因,上游序列为:
5′-GGAACCATCCACAGGGAAGA-3′;
下游序列为:
5′-CACTGGGATGGTGTCGTTTC-3′。
1.3.5 Western Blot技术检测TRPV4的表达
PC12细胞培养在6孔板48 h,β-细辛醚孵育4 h,谷氨酸处理5 min,预冷的1×PBS洗涤2次。每组加入150 μL Laemmli裂解液,得到全细胞蛋白提取液,沸水中变性10 min得到蛋白样品。用BCA法测蛋白浓度,蛋白样品经SDS-PAGE电泳压缩并分离目的蛋白。将分离的蛋白质湿转移到PDVF膜上进行膜转移。在室温脱脂牛奶中阻断印迹1 h,一抗(1∶1 000)覆盖,4 ℃冰箱孵育过夜,PBST洗涤3次后用二抗(1∶1 000)覆盖,室温孵育1 h广泛洗涤。采用化学发光法显影,用Image J对样品条带进行量化分析。
1.3.6 免疫化学技术检测TRPV4的表达
PC12细胞接种在24孔板多聚赖氨酸包被的盖玻片上,培养48 h后,β-细辛醚孵育4 h,谷氨酸处理5 min,预冷的1×PBS清洗2遍。4%多聚甲醛固定15 min,1×PBS慢速清洗3次。 0.5% Triton X-100 通透10 min,清洗3次,5% BSA封闭1 h并清洗,200 μL anti-TRPV4 (1∶100)覆盖,4 ℃冰箱孵育过夜,次日取出,室温复温1 h,荧光二抗(1∶1 000)覆盖,室温孵育1 h,加入300 μL DAPI原液,室温避光静置20 min,洗涤细胞2次。在载玻片上滴一小滴抗荧光猝灭剂,将盖玻片倒扣在载玻片上。透明指甲油封片,40倍物镜进行荧光成像。
1.3.7 Lipofectamine 2000脂质体转染实验
当细胞密度达60%时转染NC-siRNA、TRPV4-siRNA、pEX-3-Ctrl和pEX-3-TRPV4试剂,将 Lipofectamine 2000和Opti-MEM混匀配制转染混合液,将NC-siRNA、TRPV4-siRNA、pEX-3-Ctrl、pEX-3-TRPV4和Opti-MEM 混匀配制稀释液,将转染混合液与稀释液混合静置20 min,转移至对应组别中继续培养6 h后,将培养液更换成高糖DMEM,过夜培养。当细胞再次进入对数生长期且密度合适时,对细胞加药处理。沉默TRPV4实验分为NC-siRNA组(A1组)、Ctrl+TRPV4-siRNA组(B1组)、 谷氨酸+NC-siRNA组(C1组)、谷氨酸+TRPV4-siRNA组(D1组)和谷氨酸+NC-siRNA+60 μmol/Lβ-细辛醚组(E1组);过表达TRPV4实验分为pEX-3-Ctrl组(A2组)、 Ctrl+pEX-3-TRPV4组(B2组)、谷氨酸+pEX-3-Ctrl组(C2组)、谷氨酸+pEX-3-TRPV4+60 μmol/Lβ-细辛醚组(D2组)和谷氨酸+pEX-3-Ctrl+60 μmol/Lβ-细辛醚组(E2组)。NC-siRNA上游序列为:
5′-UUCUCCGAACGUGUCACGUTT-3′;
下游序列为:
5′-ACGUGACACGUUCGGAGAATT-3′。
TRPV4-siRNA上游序列为:
5′-GGAGUCCUGUUCUUCUUUATT-3′;
下游序列为:
5′-UAAAGAAGAACAGGACUCCTT-3′。
1.3.8 数据分析
实验结果运用GraphPad Prism 6软件进行处理和分析,实验结果用(平均值±标准差)表示。以P<0.05(*),P<0.01(**),P<0.001(***),P<0.000 1(****)作为显著性水平进行分析。
2 结果与分析
2.1 β-细辛醚对Ca2+浓度的影响
本文采用高分化的PC12细胞培养48 h,用β-细辛醚预先培养PC12细胞4 h,再用谷氨酸实时处理PC12细胞,测定Ca2+浓度和TRPV4的表达;沉默和过表达TRPV4检测PC12细胞内Ca2+浓度的变化。
β-细辛醚对谷氨酸处理PC12细胞Ca2+浓度的影响如图1所示。
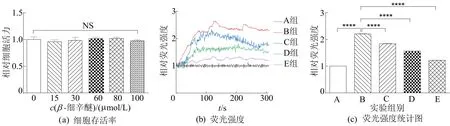
图1 β-细辛醚对谷氨酸处理PC12细胞Ca2+浓度的影响
图1中,NS表示无显著差异。从图1a可以看出,PC12细胞用不同浓度β-细辛醚处理24 h后细胞活力没有降低;从图1b、图1c可以看出,与正常对照组(A组)相比,40 mmol/L谷氨酸(B组)处理PC12细胞导致Ca2+浓度显著增加,说明细胞内钙稳态发生变化;与B组相比,谷氨酸+15 μmol/Lβ-细辛醚组(C组)、谷氨酸+30 μmol/Lβ-细辛醚组(D组)和谷氨酸+60 μmol/Lβ-细辛醚组(E组)分别用不同浓度的β-细辛醚预先处理PC12细胞4 h明显抑制Ca2+浓度的增加,抑制效果具有剂量依赖性。结果表明,β-细辛醚能够抑制谷氨酸诱导的PC12细胞Ca2+超载。
2.2 β-细辛醚对TRPV4表达的影响
采用实时荧光定量PCR、Western Blot和免疫荧光技术评估谷氨酸处理及不同浓度β-细辛醚对TRPV4表达的影响,如图2、图3所示。

图2 TRPV4 mRNA和蛋白的表达
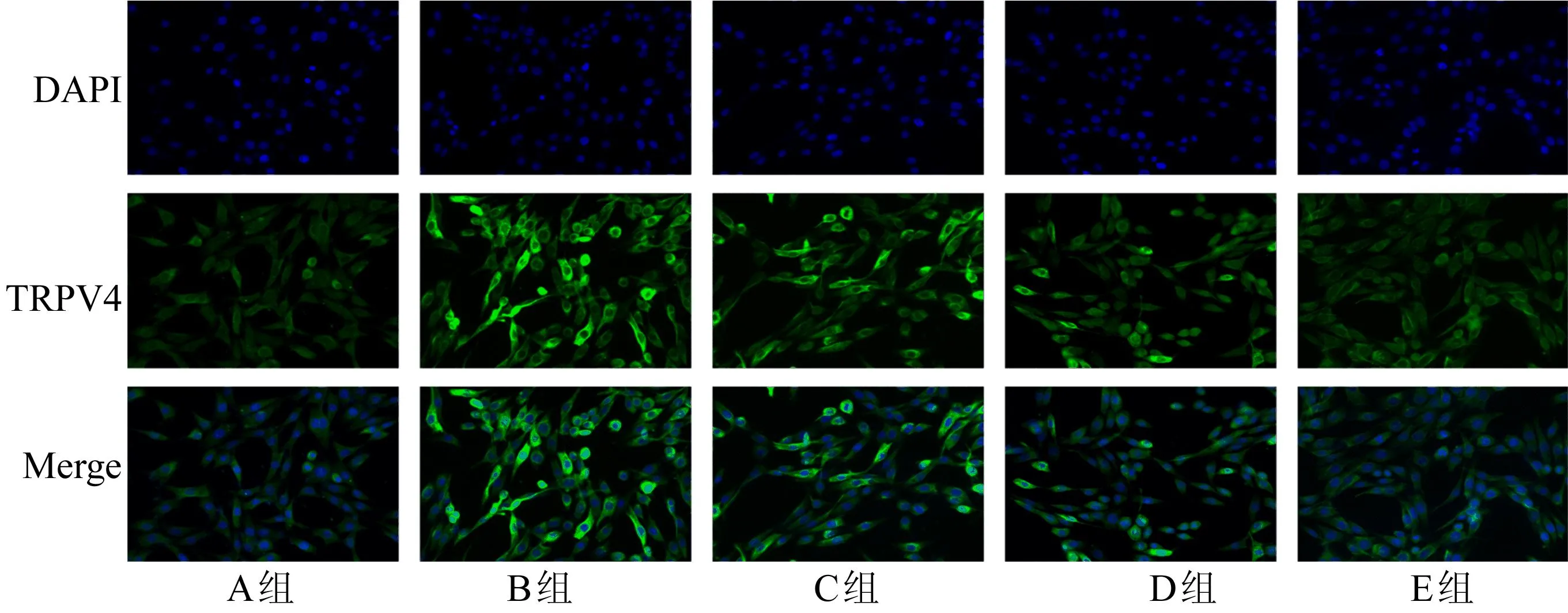
图3 TRPV4荧光图
由图2、图3可知,与A组相比,B组TRPV4在mRNA和蛋白水平表达明显升高;与B组相比,C组、D组和E组有效地抑制TRPV4 mRNA和蛋白的表达,抑制效果呈剂量依赖性。结果表明,40 mmol/L谷氨酸处理PC12细胞提高了TRPV4的表达,β-细辛醚预先处理PC12细胞4 h剂量依赖性地降低了TRPV4的表达。TRPV4通道是否参与谷氨酸诱导的Ca2+超载有待深入研究。
2.3 沉默TRPV4对Ca2+浓度的影响
为了研究TRPV4通道在谷氨酸诱导PC12细胞Ca2+超载中的作用,实验构建TRPV4-siRNA载体来沉默TRPV4的表达,采用实时荧光定量PCR和Western Blot确定转染效率,结果如图4所示。
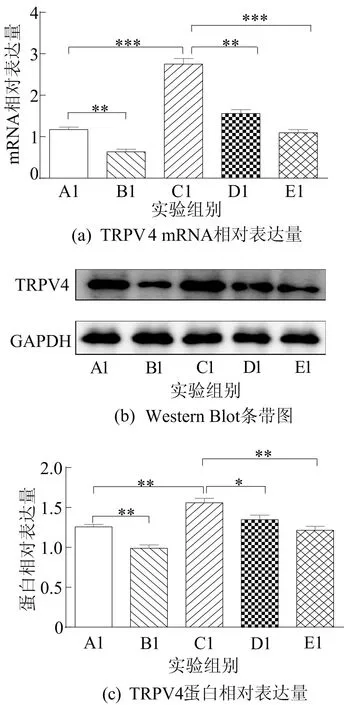
图4 TRPV4-siRNA载体沉默TRPV4的表达
从图4可以看出,与NC-siRNA组(A1组)相比,Ctrl+TRPV4-siRNA组(B1组)中TRPV4 mRNA和蛋白表达显著降低;与谷氨酸+NC-siRNA组(C1组)相比,谷氨酸+TRPV4-siRNA组(D1组)中TRPV4 mRNA和蛋白的表达显著降低。因此,TRPV4-siRNA载体构建成功。与C1组相比,谷氨酸+NC-siRNA+60 μmol/Lβ-细辛醚组(E1组)中β-细辛醚预先处理4 h显著降低TRPV4的表达。
随后对转染后的细胞进行钙成像实验,结果如图5所示。从图5可以看出,与A1组相比,C1组Ca2+浓度明显上升;与C1组相比,D1组和E1组都显著降低Ca2+浓度水平。实验结果表明沉默TRPV4和预先给与60 μmol/Lβ-细辛醚处理两者均能明显降低谷氨酸诱导的Ca2+超载。
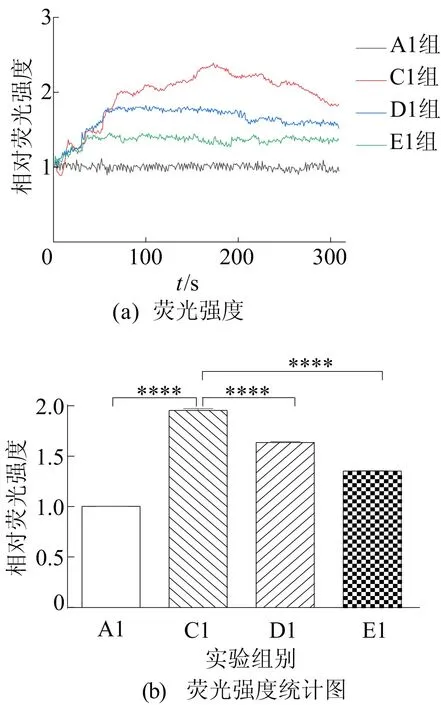
图5 沉默TRPV4对Ca2+浓度的影响
2.4 过表达TRPV4对Ca2+浓度的影响
为进一步探究β-细辛醚降低谷氨酸诱导Ca2+超载的靶点,实验构建pEX-3-TRPV4载体过表达TRPV4,采用荧光定量PCR和Western Blot技术确定转染效率,如图6所示。
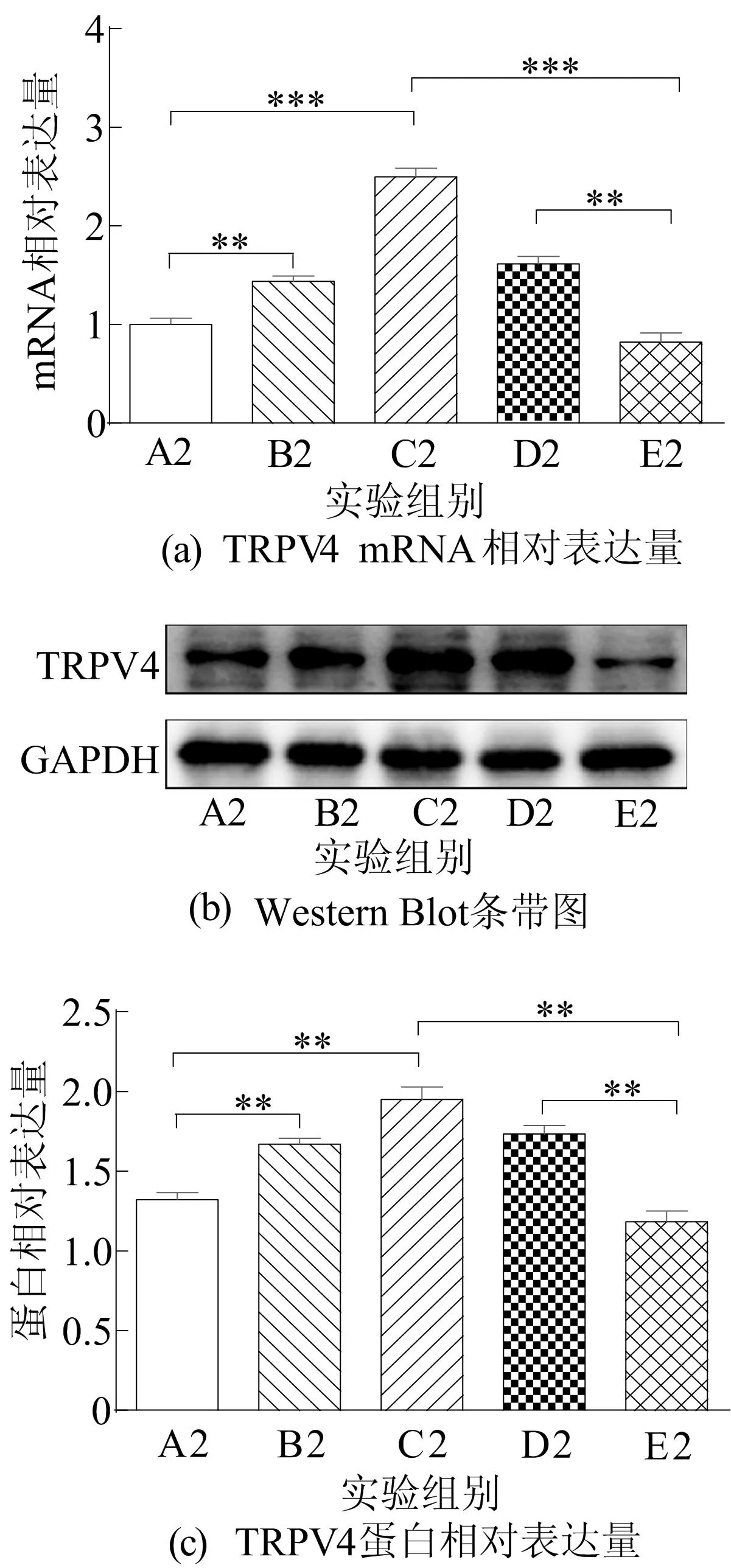
图6 pEX-3-TRPV4载体过表达TRPV4
从图6可以看出:与pEX-3-Ctrl组(A2组)相比,Ctrl+pEX-3-TRPV4组(B2组)的TRPV4 mRNA和蛋白表达明显增加;与A2组相比,谷氨酸+pEX-3-Ctrl组(C2组)的TRPV4表达明显增加;与谷氨酸+pEX-3-Ctrl+60 μmol/Lβ-细辛醚组(E2组)相比,谷氨酸+pEX-3-TRPV4+60 μmol/Lβ-细辛醚组(D2组)部分逆转了β-细辛醚抑制TRPV4的表达。因此实时荧光定量PCR和Western Blot结果表明,pEX-3-TRPV4载体构建成功。
本文对Ca2+浓度进行分析实验,结果如图7所示。
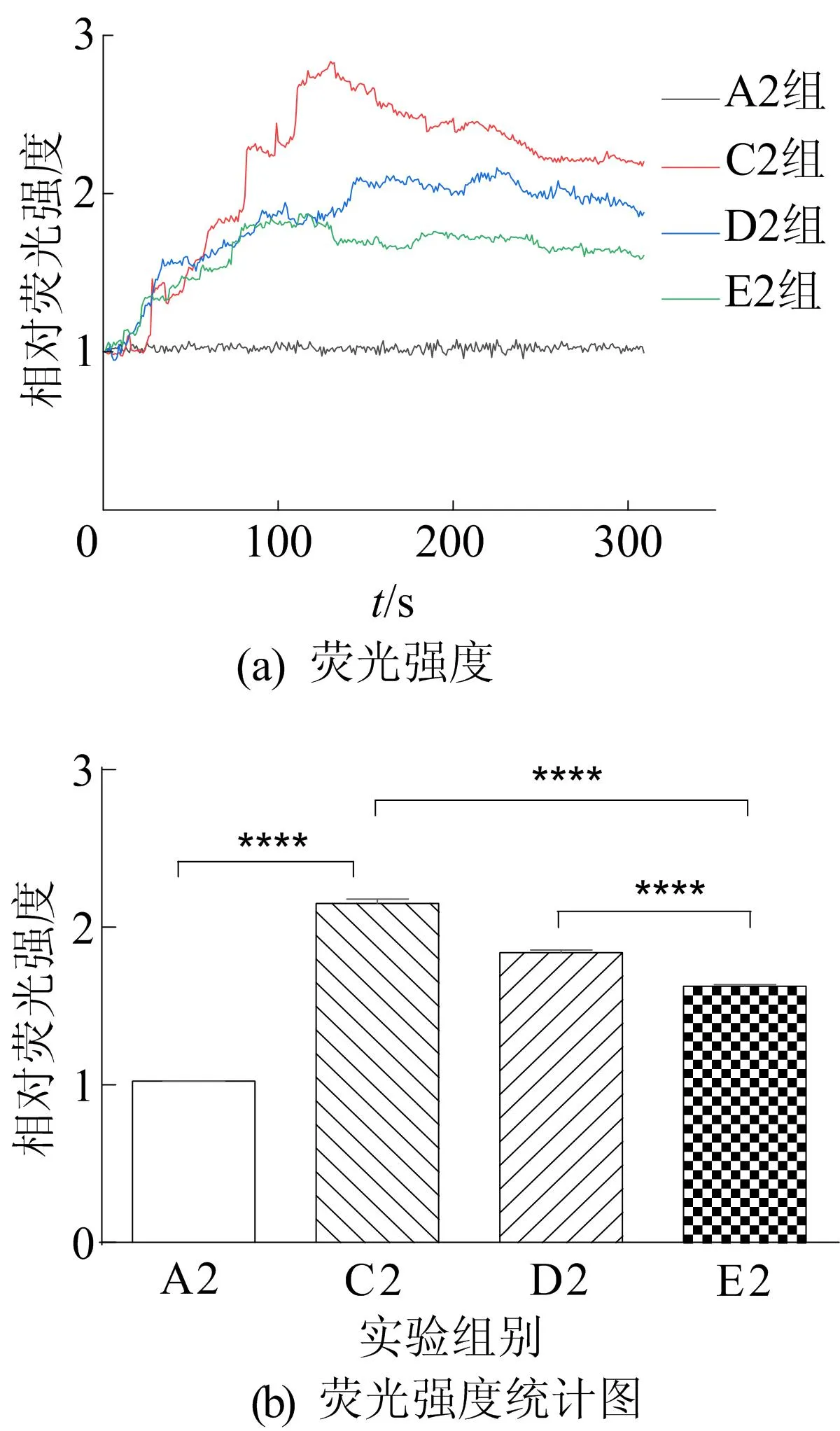
图7 过表达TRPV4对Ca2+浓度的影响
从图7可以看出:与A2组相比,C2组Ca2+浓度显著升高;与C2组相比,E2组Ca2+浓度显著减少;与E2组相比,D2组过表达TRPV4部分逆转了60 μmol/Lβ-细辛醚抑制谷氨酸诱导的Ca2+浓度的增加。钙成像结果表明,60 μmol/Lβ-细辛醚能够抑制谷氨酸诱导的Ca2+超载;过表达TRPV4能够促进谷氨酸诱导的Ca2+超载。
3 结 论
鉴于Ca2+超载是导致兴奋性毒性细胞死亡的主要途径,本研究表明谷氨酸可以引起PC12细胞Ca2+稳态紊乱,β-细辛醚剂量依赖性地抑制谷氨酸诱导的Ca2+超载;谷氨酸促进TRPV4的表达,β-辛醚剂量依赖性地抑制TRPV4的表达。为了探究β-辛醚降低谷氨酸诱导Ca2+超载的作用靶点,构建TRPV4-siRNA和pEX-3-TRPV4载体来沉默和过表达TRPV4。结果表明:沉默TRPV4显著缓解了谷氨酸诱导的Ca2+超载;过表达TRPV4促进了谷氨酸诱导的Ca2+超载。
上调TRPV4的表达可增强Ca2+超载,促进谷氨酸毒性。下调TRPV4的表达可能间接作用于谷氨酸受体有效抑制Ca2+超载。谷氨酸毒性是一种复杂的损伤,研究表明,β-细辛醚作为TRPV4的拮抗剂,是抑制兴奋性毒性的潜在药物。实验结果提高了人们对谷氨酸毒性发病机制的认识,并为谷氨酸毒性的治疗提供了新的靶点。
猜你喜欢
小哥白尼(野生动物)(2019年5期)2019-08-27
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
三门峡职业技术学院学报(2017年1期)2017-06-05
中国洗涤用品工业(2017年2期)2017-04-16
中国比较医学杂志(2017年5期)2017-01-17
吉林大学学报(医学版)(2015年4期)2015-12-17
华南农业大学学报(2015年5期)2015-12-04
医学研究杂志(2015年12期)2015-06-10
发明与创新(2015年33期)2015-02-27
