
植物根系吸水模型研究进展
2024-02-29 07:15杨乐龙兰艾施荣施俊林王龙飞李紫怡
科学技术与工程 2024年4期
杨乐, 龙兰, 艾施荣, 施俊林, 王龙飞, 李紫怡
(江西农业大学计算机与信息工程学院, 南昌 330045)
水资源紧缺严重制约着中国国民经济的可持续发展,农业一直是用水需求最大的产业,主要消耗于灌溉[1]。为解决当前的缺水问题,大力发展节水农业,提高节水农业的效率,已成为当今世界各国发展节水农业的一项重要战略。现代农业节水技术是节水和高效用水的措施之一,如喷滴灌,但是其效果并不理想,因此探索农业高效用水理论体系和农业节水的发展势在必行。目前,节水灌溉理论与技术的研究和创新是很多研究者关注的热点,具有广泛的应用前景。
农业高效用水的实现必须对农业土壤-植物-大气连续体(soil-plant-atmosphere continuum,SPAC)水分动力学进行研究分析[2]。SPAC中的物质转移和能量传输,以及农业中水资源的高效利用与管理都依赖于土壤水分转移规律的研究,而对根系吸收规律的定量描述是 SPAC体系中土壤水分迁移的重要依据[3]。
水稻是中国的主要粮食作物之一。中外很多学者对不同水分条件下的旱作物根系的吸水模型进行了研究,但是有关水稻根系的吸水模型研究是被忽略的重要群体。不同作物、同一作物的不同品种在不同生长环境下,其根系的生长状况也会有所不同[4]。水稻根系的生长受土壤水分等因素的影响,根系对土壤水分的吸收状况又与根系在土壤中的分布有关[5-7]。因此,水稻根系在土壤中的生长分布的研究和根系吸水模型的逐步完善对提高水稻产量和实施节水灌溉具有很重要的意义。
鉴于此,对吸水模型进行了分类和对比分析,分析水盐胁迫的条件下根系的吸水模型,综述当下根系吸水模型的研究现状,展望了水稻根系吸水模型的未来发展趋势。
1 根系的吸水机理
植物的根系吸水机理十分复杂,根系不同部位对水分的吸收速率不一致,根系的发育时间及程度、根系在土壤中的分布以及土壤的性质等差异会影响根系对水分的吸收[8]。植物通过根系从土壤中吸水水分,为植物提供生长、代谢等多种生理活动。植物根系的吸水机理主要有两种:一种是主动吸水,即在弱蒸腾条件下,由离子主动吸收,以及在根内外水势差的影响下根系主动吸水;另一种是被动吸水,即通过蒸发引起的水分差异,使根系吸收水分[9]。
2 根系吸水的影响因素
作物根系吸水受到多种因子的影响,其中土壤性质、作物生理特性和微气象条件是决定作物根系吸收水分的关键因子。
2.1 土壤因子
土壤因素对根系吸收水分的影响,主要表现为土壤水力特征对土壤、土壤-根系间水分迁移的影响,以及不同土壤物理特征对根系生长的影响[10-11]。已有的植物根系吸收模型表明,土壤水分不饱和导水率、土壤含水量等对植物根系吸水速率有重要影响。由于土壤的质地、结构的不同,土壤的持水率也会有很大的不同,这就会对土壤的水分转移和根系吸收土壤水分的程度产生影响。另外,在不同的土壤条件下,其根系发育及空间分布也存在着明显的差别。根系在土壤中的分布直接影响着根系的吸水能力和吸水速率。
2.2 作物因子
在 SPAC系统中,作物是连接土壤和大气的关键。根系是植物吸收水分最重要的器官,一般以其有效根长密度来表示其在土壤中的分布状态。通常认为,根系的有效根长密度越大,根系的吸水能力越强,在土壤含水量充足的情况下,根系的吸水速度越慢,而当土壤干燥后,根系的吸水能力就越强,这与发达根系可以降低土壤的水流阻力有关[12]。但是这种效应具有一定的时效性,在长时间的干旱胁迫下,植株会表现为根系萎缩,失去正常的生理机能,最终导致植物的死亡。植物的吸水速率除了与植物根系有关外,还与植物本身生长和对水的需求有关。土壤水分含量较高时,叶表面积与根系吸水速率之间存在着显著的相关性,而土壤水分含量较低时,叶片指数减小,叶表面积会显著下降,降低了作物对水分的敏感度[13]。
2.3 气象因子
气候条件是影响作物需水量的主要因素,因此, SPAC内部的水势梯度是可以调节的。在不同的蒸腾条件下,作物-土壤水分的变化规律是不一样的,即使在高空气蒸发力、高湿度情况下,作物也会出现干旱;在大气蒸发量低的情况下,即使在土壤水分含量较低的情况下,作物也不一定会出现缺水现象[14]。
综上所述,很多因素会影响作物根系的吸水率,但在研究根系吸水并建立相关根系吸水模型的时,必须将以上3个因素考虑进去,才能更加客观的描述作物根系的吸水过程。
3 根系吸水模型的分类
由于植物根系吸水机理十分复杂,并且影响因素众多,学者们对其进行大量的理论分析并提出各种根系吸水模型。根系吸收模式的研究始于20世纪50年代末60年代初。根系吸水模型是根据植物根系的吸水机理和实验结果建立的,利用数学等方法对植物根系的吸水性进行定量描述。将根系吸水模型分为:微观模型和宏观模型[15],根系吸水模型的分类及其优缺点如表1[16-34]所示。

表1 根系吸水模型的分类及其优缺点[16-34]Table 1 Classification of root water uptake models and their advantages and disadvantages[16-34]
3.1 微观模型
微观模型以单根作为研究对象,主要利用单根径向流模型描述根系的吸水性,进而构建根系吸水模型,因此又称为单根径向流模型。该类模型以单根吸水特性作为分析切入点,将植物根系分解成一系列长度无限、半径均匀、吸水均匀的柱状体,在不考虑地心引力的情况下,假设水从土壤向根部流动为径向流,从而建立土壤中水分流向根系的吸水模型。该类模型的主要代表有Gardner模型[16]、Molz模型[17]、Campbell模型[18]等。
3.1.1 Gardner模型
Gardner[16]首次提出微观模型,该模型将根系视为一个半径rr无限长的圆柱体,根系在不同方向上的吸水性、土壤的初始条件以及渗透系数等参数都随着根系的生长保持不变。此外,忽略重力作用的影响,土壤中水的流动可以视为往径向流动,其模型表达式为
(1)
3.1.2 Molz模型
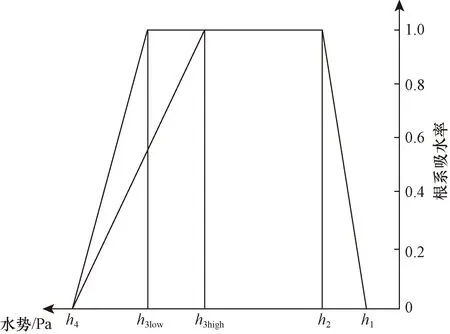
Molz模型[17]的几何结构由半径为rr的圆柱形根部和半径为rs的均质土壤圆柱体组成,不考虑沿根部长度变化的问题。该模型将根系吸水条件下土壤中水分向根表面流动和水分在根组织内的流动联系起来,考虑了根的水力特性,提出了一个单根吸水时土壤-根系系统水流运动模型。由于假设根组织在re和rr之间是均匀的,因此边界条件将应用于位于半径为re的根内皮层。穿过内皮层后,水在进入木质部之前穿过几个细胞。因此,组织域由re 土壤中,水分在根部的流动近似呈径向流动,其基本方程为 (2) 忽略溶质效应,根系内部组织细胞保持平衡,水分在根组织内流动的基本方程为 (3) 3.1.3 Campbell模型 Campbell模型[18]假设水分在一定时期内在SPAC中的运动处于稳定状态,且SPAC中的水势是连续的,由内聚力控制。因此该模型可以用模拟电流传输的欧姆定律来描述水分在这个系统中的运动。 (4) 上述模型主要对根系的吸水机理进行了分析,理论上能够描述土壤-根系的水分运动。但由于根阻力、根水势、根系吸水率等参数的测定很困难,所以难以用单根吸水模型对整体土壤-根系的水流运动进行定量研究。此外,由于该类模型假设的根系几何形态与现实存在较大差异,而且不能完全考虑到根系吸水能力随根系生长在空间中分布的变化,根系与土壤之间边界条件的确定也十分困难。因此,该类模型难以应用于田间实际的土壤水分运动研究。 宏观模型是将根系作为一个整体进行研究,其原理是将作物根系看作一个以一定方式分布在整个根层的吸水器,认为同一土层深度的根系是均匀分布,或者同一深度在主根径向距离r处均匀分布。该类模型不需要弄清楚根系的整个吸水机理,它不需要去研究那些难以界定的土壤-根的参数。根系吸水模型的构建是在描述土壤水分运动的Darcy-Richards方程[35]上加一个汇源项来实现,即通过根对水分的吸收速率来定量分析作物生长过程中土壤水分的运动,其数学方程为 (5) 式(5)中:▽为矢量微分算子。 宏观模型的分类主要围绕S展开,根据S的不同构造原理和方法,将宏观模型通常分为两大类:①由微观模型引申而来的Ⅰ类模型;②基于根对土壤水分的吸收受根区土壤水势的影响提出的Ⅱ类模型[19]。 3.2.1 Ⅰ类模型 Cardon等[19]基于Gardner[16]描述的土壤水分向根部流动的微观物理学,提出了通用的Ⅰ类模型,其基本表达形式为 S=BK′G (6) 式(6)中:B为水流的几何参数;K′为传导率项(通常指土壤的水力传导率);G为从土壤到根的水势梯度(通常表示根和土壤的水势差)。 事实上,微观模型也属于Ⅰ类模型。很多研究者采用Ⅰ类模型去研究根的吸水性,最具代表性的Ⅰ类模型就是借鉴欧姆定律的思想,根据根系吸水速率与土壤-根系的水势差来建立模型,也称为欧姆定律模型,其基本表达式为 (7) 该模型的应用需要测定一系列的参数,比如土壤根系水势、土壤和根系对水流的阻力等。比较常见的模型有Hillel模型[20]、Herklranth模型[21,36]等。 (1)Hillel模型。Hillel模型[20]是在Cardon等[19]研究的基础上加入土壤对水流的阻力参数,可表示为 (8) (2)Herklranth模型。Herklranth等开发了一种强调根-土接触效应可能性的替代模型,在Vanden-Honert[9]假定的基础上增添了一个饱和度的概念,通过假设有效根系渗透性与土壤的相对饱和度成比例,进行更好的数据拟合。 (9) 该模型解决了土壤-植物系统中的总流动阻力会随着土壤干燥而增加的问题,但是观测数据没有排除根系随着土壤的干燥而收缩产生的额外接触阻力,从而降低水分的吸收率。 上述模型的重点在于土壤-根系系统,模型中涉及的参数只有土壤参数和根系参数,没有考虑SPAC系统中大气因素的影响,即只考虑了作物因子和土壤因子,忽略了气象因子。此外,模型中的一些参数测量具有一定难度,比如根系阻力、土壤阻力等。因此,该类模型在实际情况中很难被应用。 3.2.2 Ⅱ类模型 Ⅱ类模型由宏观经验函数组成,基于根对土壤水分的吸收受根区土壤水势影响提出,其一般形式为 S=α′(h,π)Smax (10) 式(10)中:π为渗透压头;α′(h,π)为无量纲变量,相当于实际S和最大吸收Smax之间的比率;h为当前土壤水势。 根据研究的主要方式方法不同,Ⅱ类模型可以分为权重因子类根系吸水模型、补偿性吸水模型、基质通量模型。 (1)权重因子类根系吸水模型。该类模型将蒸腾作用作为主要的参考因子,蒸腾量的分布与根系层土壤剖面相对应,作物蒸腾量在深度上按比例分配。权重因子通常是指土壤水分含量、土壤水势、水分传导系数、扩散系数、根长密度等相关函数。该类吸水函数的基本优点是吸水函数在整个根层中的积分等于作物的蒸腾速率,即 (11) 式(11)中:Tp(z)为根系在土壤深度为z时的蒸腾速率;Sp为根系在土壤深度为z时的吸水函数。 根据方程可以得到以蒸腾作用为参考因子根系吸水模型的一般表达式为 Sp(z)=β′(z)Tp(z) (12) 该类模型[式(12)]按照一定的权重因子β′(z)将潜在蒸腾量Tp(z)分配到不同土壤深度,得到不同土壤深度的潜在吸水量Sp(z)。 Molz等[22-24]提出了具有代表性的权重因子类根系吸水模型,该模型以作物根系密度和土壤水扩散度之积作为权重因子对蒸腾量在整个根系层中进行分配。该模型的根系吸水函数包含了根系吸水的3个主要影响因子,即作物因子、土壤因子、气象因子,体现了一定的物理含义。 (13) Molz等[37]用K(θ)替代D(θ),它强调从土壤剖面高水率处的吸水量,但是在很多情况下,K(θ)随土壤含水率的变化幅度比较大,用K(θ)会放大高含水率处的吸水量。 (14) 罗毅等[38]对Selim-Iskandar模型进行了改进,在权重因子中加入土壤水势对根系吸水的影响函数,考虑了土壤水分因子的影响,可表示为 (15) 式(15)中:α(θ)为根效率函数。 (16) (17) 式(17)中:Rsr为土壤与根系接触处的阻力;λ(θ)为与土壤含水率有关的参数,其值在0~1。 当根长密度L(z)和作物蒸腾量T一定时,含水率越大,吸水速率就越大,土壤剖面饱和处的吸水速率就越大。但在实际情况下,当土壤处于饱和状态或者临近饱和状态时,大多数作物根系的吸水量等于0或接近0。因此,有学者对Molz-Remson吸水公式[式(13)]进行改进,既保留了原有吸水函数公式的优点,同时又引入根效率函数α(θ)调节较高含水率处的吸水含量,更加符合实际情况。 (18) 式(18)中:θ0为枯萎点含水率;θan为微生物厌气点含水率;θi为当S=Smax时的土壤临界点最低含水率;θs为土壤饱和含水率。 表2[22-24,37-40]为对Molz-Remson模型的改进对比。 表2 Molz-Remson模型的改进对比[22-24,37-40]Table 2 Improved comparison of the Molz-Remson model[22-24,37-40] 1978年,Feddes等[26]提出了另一种根系吸水模型,该模型采用根区土壤水势作为参变量,构建出一种根系吸水模型,可表示为 (19) 式(19)中:α[h(z)]为根区土壤水势对根系吸水的影响函数,即水分胁迫因子,定义为 (20) 式(20)中:h3为水分胁迫的临界水势,指的是图1中的h3low或h3high、h3low、h3high分别为低水分胁迫的临界水势和高水分胁迫的临界水势。 h3low、h3high分别为低水分胁迫的临界水势和高水分胁迫的临界水势图1 土壤水势响应函数[25]Fig.1 Soil water potential response function[25] h1为饱和土壤中厌氧状态下的水势(通常为0);h2为氧气胁迫的临界水势;h4为植物枯萎状态的临界水势。 图1[25]中是对式(20)中分段公式进行展示。根系的吸水率在 [h2,h3]之间达到潜在最大值。在 (h1,h2)、 (h3,h4)两个区域中根对水分的吸收速率呈直线下降趋势,其原因是受到氧气胁迫和水分胁迫。当h≤h1或h≥h4时根系的吸水率为0。 Feddes模型虽然考虑了根区土壤水势的变化,但是忽略了根系在土壤中的分布得根系的吸水也会造成一定的影响。根据资料表明[10],根系的吸水能力与根系密度分布、土壤-根系水势差成正比。 随后罗毅等[38]对Feddes模型进行改进,在权重因子中加入了根系密度分布函数,考虑了根系分布对根系吸水能力的影响,可表示为 (21) 权重因子类根系吸水函数较为全面的考虑了根系的作物因子、土壤因子、气象因子3个根系吸水影响因素,比较符合实际情况。该类吸水函数最大的优点是吸水函数在整个根层中的积分等于作物的蒸腾速率,也反映了植物通过根系从根系中吸收水分然后进行蒸腾作用的整个运动过程。 量化根系吸水率有助于了解植物的用水及其对环境变化的响应,土壤水势、土壤导水率和土壤水分扩散率是影响根系吸水率的关键因素,Deng等[41]以青藏高原高寒草甸作为研究对象,选取改进的Feddes模型[25-26]、Molz-Remson模型[22-24]和Selim-Iskandar模型[37]来模拟高寒草甸的根系吸水率,充分考虑上述影响因素并将其作为模型的主要参数。在Feddes模型的改进中,将实测的根系密度指数添加到模型当中,得到的模型能够更加准确的反应土壤剖面中的根系吸水率,改进的模型可定义为 (22) 在模型的比较中,考虑土壤导水率的模型更好。Selim-Iskandar模型[37]模拟的结果比改进的Feddes模型[25-26]和Molz-Remson模型[22-24]更准确。Selim-Iskandar模型的仿真性能比改进的Feddes模型和Molz-Remson模型更高。因此,Selim-Iskandar模型更适合高山草甸。改进的Feddes模型在根系吸水率的研究中能够得到更加广泛的应用,主要应用于农作物和树木,并显示良好的模型性能,但是在一些水分盐分胁迫的环境下可能会影响模型的参数。Molz-Remson模型和Selim-Iskandar模型的研究相比较而言较少,模型中的参数在实际环境中获取复杂且困难,但是其对根系吸水率的作用方式很难受到自然环境的影响,更加的稳定。针对其他类型的土壤环境需进一步细化研究,以扩大模型在不同环境下的适用性。 (2)补偿性吸水模型。大多数模型的假设条件过于简化,尤其在一些初始和边界条件都理想化的状态下,不能精确地表述根系吸水过程。如Feddes模型[25-26]认为,局部根区根系的吸水不会受到其他根区土壤水势的影响,但是植物的根系系统是一个整体,根区之间的吸水会相互影响。然而,通过引入一个附加参数,可以更加灵活的描述根系吸水的过程。Skaggs等[11]提出引入一个补偿机制能够平衡灌溉条件下出现水分分布不均匀的现象。Jarvis[42]首次提出了根系吸水补偿的概念,通过计算胁迫指数函数ω,即未补偿实际蒸腾量与潜在蒸腾量的比值,将宏观补偿吸水模型概念化。植物胁迫指数函数为 (23) 式(23)中:Ta为未补偿实际蒸腾量;Tp为潜在蒸腾量;Lr为最大根系深度;β(z)为根系密度分布函数;α[h(z)]为水分胁迫因子;当ω=1,表示整个土壤根区没有应力。 补偿吸水的情况下,实际蒸腾量为 (24) 式(24)中:ωe为植物胁迫指数的临界值,0<ωe<1。 补偿吸收的物理基础可以用类似于土壤-大气连续体中水流的欧姆定律来解释。补偿机制根系吸水模型中比较典型的模型有:Lai and Katul模型[27]和Li K Y模型[28-30]。 Lai and Katul模型[27]中把吸水项表达成潜在蒸腾量的线性函数,对S的表达采取Perrochet[43]的规范形式,其模型表达形式为 S(θ,z,t)=α(θ)g(z)Ep(t) (25) 式(25)中:Ep(t)为潜在蒸腾作用;α(θ)为根效率函数,大多数情况下取决于土壤的水势,Lai等[27]将其定义为 (26) 式(26)中:α1为补偿机制函数;α2为水分胁迫函数;θ0为枯萎点的土壤含水率。 g(z)为根密度函数,借鉴Hoogland等[12]和Jackson等[44]的研究将其表示为 (27) g(z)=-azlna (28) 式中:c为常数;a为经验参数。 该模型通过局部含水量和整体根系含水量的最大值来描述,确保表层含水率接近枯萎含水率时也能进行吸水。通过枯萎含水率和饱和含水率来表示水分胁迫函数,其水分胁迫函数对土壤水势参数比较敏感。 Li K Y模型[28-30]引入一个加权胁迫指数,模型的表达式为 (29) 式(29)中:i为土壤层;Pt为潜在蒸腾速率;Fi为根长密度分数;Si为第i层的吸水量;αi为加权胁迫指数;ΔZi为所考虑的层厚度。 由于根区某一部位的水分胁迫可以通过其他较湿润部位的水分吸收来补偿,通过加权胁迫指数在根区分配潜在蒸腾量(PT),该指数是根系分布和土壤水分有效性的函数。 如果根区内的土壤分为n层,则给定层i的加权应力指数βi可表示为 (30) 式(30)中:λ为指数项常数。 该模型引入一个与根系分布和土壤水分胁迫有关的加权胁迫函数,但其水分胁迫函数对土壤水势参数不是很敏感。 (3)基质通量模型。由于上述补偿机制的根系吸水模型对根系吸水机理方面的表述较少,涉及的参数难以获取。因此,可以通过在模型中引入基质通量势法来明确无量纲胁迫参数与可测量系统参数之间的相关关系,从而建立基于物理机制的根系补偿吸水模型。这些可测量的系统参数包括根长密度和根系导水率等。基质通量势是某一土壤非饱和导水率的积分,是描述水分从土壤向根系径向流动的土壤导水特征的线性参数,与土壤水分运动、根系吸水和限制土壤水力条件密切相关。de Jone等[31]提出的吸水模型引入基质通量。 (31) 式(31)中:h、hw分别为当前土壤水势和枯萎时的水势;K为土壤水分的传导系数,即导水率。 基于基质通量势的根系吸水模型可表示为 Si=ρ(Mi-M0) (32) 式(32)中:Si为第i层的吸水量;Mi为第i层的基质通量势;M0为根表面基质通量势;ρ为与根长密度、根长和根半径有关的参数。 土壤在湿润的状态下,根系的实际蒸腾量和最大的潜在蒸腾量分别表示为 (33) (34) 式中:Ea为实际蒸腾量;ΔZi为第i层的厚度;Emax为最大的潜在蒸腾量;ρi为第i层与根长密度、根长和根半径有关的参数。 de Jone等[31]没有采用胁迫指数函数的形式,而是考虑实际根系吸水能力和潜在蒸腾量之间的关系。当根系的实际吸水能力大于潜在蒸腾量时,实际蒸腾量等于潜在蒸腾量;当根系的实际吸水能力小于潜在蒸腾量时,实际蒸腾量等于水分胁迫下的蒸腾量。 在一些干旱地区或半干旱地区,植物经常会遭遇水分和盐分胁迫,水盐胁迫会对根系的吸水功能造成影响,因此对水盐共同胁迫作用下根系吸水的定量研究,对于干旱地区或半干旱地区的农作物产量提升以及水资源管理十分重要。 水盐胁迫对根系吸水的影响主要体现在根系细胞的渗透作用上。在大多数研究中都是单独研究水分胁迫对根系吸水的影响,或者单独研究盐分胁迫对根系吸水的影响。相比于单独水分胁迫、盐分胁迫,植物在两者共同交互作用下需要消耗更多的能量来吸收水分[4]。 当植物在生长过程中,实际的蒸腾速率小于潜在蒸腾速率时,根系的吸水速率就会比潜在的吸水速率小。Feddes等[26]和Belmans等[45]提出了一个根系吸水模型,认为根系吸水速率与潜在蒸腾速率成比例,可表示为 S(z)=β(z)α(h)Tp (35) 式(35)中:β(z)为根系密度分布;Tp为潜在蒸腾速率;α(h)为水分胁迫系数。 对于盐分胁迫的情况下,就是将盐分胁迫系数α(π)替换水分胁迫系数α(h),可表示为 S(z)=β(z)α(π)Tp (36) 基于水分胁迫的条件,α(h)具有多种表示形式,其中较为经典的两个形式分别是分段线性函数和连续平滑S型函数。Feddes等[26]提出了分段线性函数,通过影响根系吸水的4个水势临界值h4 (37) 式(37)中:p1为可调节的经验参数;h50为α(h)=0.5时的水势。 Musters等[47]提出了一种非线性函数,可表示为 (38) 式(38)中:t为植物生长时间;α(h)随着h的减小而减小。 与水分胁迫响应函数相比,盐分胁迫响应函数也有两种形式,即分段线性函数和连续光滑函数。线性函数的常见形式为 (39) 式(39)中:a和b为经验参数;π为渗透压头。 van Genuchten等[48]提出了盐分胁迫效应的平滑S型函数为 (40) 式(40)中:p2为可调节的经验参数;π50为α(π)=0.5时的渗透压头。 水盐共同胁迫对根系吸水的影响主要体现在细胞渗透作用上。如何结合水分和盐分胁迫的影响是具有一定难度的挑战[4]。学者们通过实验模拟作物根系吸水,并提出了两种模型来描述水分和盐分胁迫下根系吸水的减少。研究表明,假设水分胁迫和盐分胁迫交互作用下对根系吸收水分的影响存在两种情况:一是两者影响效应相加;二是两者影响效应之积。van Genuchten等[48]提出了加权相加模型,可表示为 (41) 式(41)中:a1和a2分别为土壤水势和渗透压头的经验系数,但在实际情况下,a1和a2难以通过实验获取。 在加权相加模型中,根系吸水是响应于土壤水势和渗透压头的加权和而发生。与加法模型相比,乘法模型以下形式应用得更广泛。 α(h,π)=α(h)α(π) (42) 式(42)中:α(h)和α(π)分别为前面提到的任何水胁迫响应函数或盐度胁迫响应函数。 此外,Homaee等[33-34]通过引入两个阈值,分别为土壤水势h*和土壤渗透压头π*,提出改进的模型。但是这两个阈值受土壤环境、植物蒸腾作用、植物品种等因素的影响,在应用时,这两个阈值的数据会存在较大误差。 (43) 学者们用不同的加法或乘法模型模拟水分和盐度胁迫下的根系吸水。Wang等[49]以中国西北地区种植的春玉米为研究对象,在田间灌溉试验的基础上,采用水量平衡法,设定不同的水量和含盐浓度组合,计算了春玉米根系的吸水率。通过构建加法模型和乘法模型来模拟水分和盐分胁迫对根系吸水的影响。任何类型的水分胁迫乘以任何类型的盐分胁迫都可以导致在水分和盐分共同胁迫下的根系吸水变化。因此,共构建4个根系吸水乘法模型[模型(a)~模型(d)]和一个根系吸水加法模型(e),可分别表示为 模型(a): (44) 模型(b): (45) 模型(c): (46) 模型(d): (47) 模型(e): (48) 加法模型的理论基础是假设水分胁迫和盐度胁迫对植物蒸腾作用的影响相似。但中国西北地区广泛种植的玉米属于抗旱品种,对水分胁迫的抗性高于对盐度胁迫的抗性,加法模型没有考虑这种差异。对于乘法模型,分别考虑了水分胁迫因子和盐分胁迫因子,两种胁迫因子之间的差异能够合理地反映在乘法模型中。5个模型的比较结果表明,乘法模型(d)最好,加法模型(e)最差。 环境中的各种因素都会影响植物根系的吸水速率,水分或者盐分的胁迫在不同生长阶段的影响不同,且不同时期所获取的数据推到的模型参数可能并不准确。因此,为了使模拟值更接近测量值,水盐共同胁迫下模型的改进是之后研究的重点。 植物根系吸水模型是研究植物根系吸水过程的数学模型,它有助于理解植物根系的水分吸收机制以及水分运输过程。针对现有吸水模型存在的问题进行总结,并对根系吸水模型在未来发展进行展望。 (1)根系参数的确定。根系吸水模型的参数通常需要基于大量的实验数据进行估计,但收集和测量这些数据是困难且耗时的过程[50-52]。此外,参数估计的准确性也受到实验条件的限制。根系吸水模型在未来的研究过程中,反映根系吸收机理的同时,可以根据具体情况引入合理的简化假设,减少模型的复杂性,增加模型的实用性。 (2)影响因素的确定。随着根系吸水模型研究的不断发展,根系吸水模型通常是基于一些简化假设和近似方法构建的,无法完全准确地反映根系吸水过程中的各种复杂因素[53]。例如,有些模型忽略了土壤非均质性和根系的精细结构,可能对吸水速率和水分分布的预测产生一定的误差。同时在实际生长中根系的吸水情况还会受到多环境因子的影响,比如光照、温度等,因此,通过模型改进的方法来模拟根系的水分吸收以及土壤水分分布的动态变化,以及根系的吸水情况与多环境因子之间的定量关系也是之后研究中需考虑的一个重要方向。 (3)动态根系吸水模型的构建。现有的根系吸水模型都是建立在根系静态的基础之上。但在实际情况下,根系的生长是动态的,这意味着根系在生长过程中会有老根死去新根产生[54],因此根系吸水模型在考虑根系分布时,应该考虑根系的动态生长。 (4)时空尺度问题的改进。根系吸水模型通常是基于特定的时空尺度开发的[55-56],对于较大尺度和长时间尺度的根系吸水行为的模拟还有待进一步研究。此外,不同类型植物的根系吸水模型也需要进一步研究和改进。 (5)宏微观模型的结合。微观模型和宏观模型在植物根系吸水模拟和预测中都发挥着重要作用[57-58]。微观模型提供了对吸水过程细节的深入理解,但需要更多的参数和输入数据。宏观模型简化了复杂性,适用于大规模土壤和植物系统,但其预测结果可能缺乏精确性。未来的研究可以集成微观和宏观模型,通过多尺度建模和耦合的方法提高模型预测的准确性和适用性。 (6)水盐共同胁迫下的模型构建。水盐胁迫交互作用下的根系吸水模型更符合实际情况,对农作物的研究具有重要意义。目前的研究主要侧重于测量根系吸水速率、根系水分和盐分的分布等指标[59-61],建立一组动态方程模拟根系吸水率、水分和盐分的分布以及植物对胁迫的响应。但是,现有的植物根系吸水模型在水分和盐分共同胁迫下的应对机制上还比较简化,没有充分考虑到根系水分和盐分的动态变化。模型的复杂性需要进一步提高,以更好地反映实际植物的生理和生态过程。 根系吸水模型的发展对深入了解植物根系吸水机制、优化农作物灌溉和改善土壤水分管理等方面具有重要作用。然而,目前的根系吸水模型在描述植物根系的复杂结构和吸水过程时存在一些局限性。在今后的研究中需要通过突破观测手段和实验技术,获取更精确、全面的根系吸水数据,深入研究根系吸水的机制,探索根系与土壤水分的相互作用,从而更准确地建立根系吸水模型,并提高其预测能力和适用性。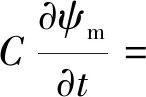
3.2 宏观模型
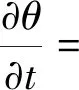
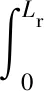



4 水盐胁迫下的根系吸水模型
4.1 水分胁迫
4.2 盐分胁迫
4.3 水盐共同胁迫
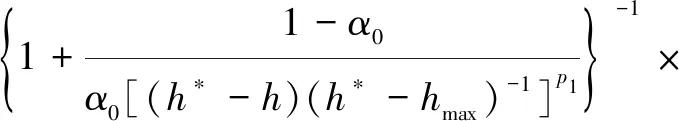
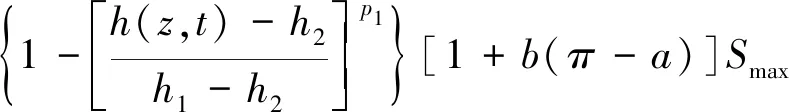
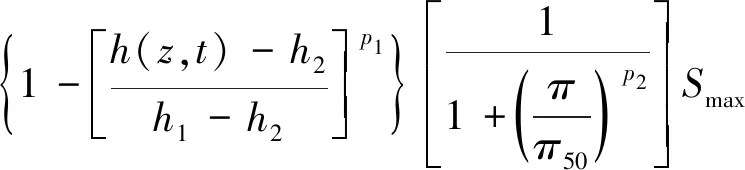
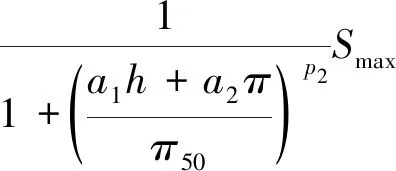
5 展望
6 结论
猜你喜欢
四川劳动保障(2021年3期)2021-06-09现代园艺(2017年13期)2018-01-19高原山地气象研究(2016年2期)2016-11-10西南农业学报(2016年5期)2016-05-17中国农业文摘-农业工程(2016年5期)2016-04-12水利科技与经济(2016年3期)2016-04-09水利科技与经济(2016年4期)2016-04-09水利科技与经济(2016年2期)2016-04-09塔里木大学学报(2014年3期)2014-03-11自然资源遥感(2014年2期)2014-02-27
